能源植物柳枝稷基因工程研究进展
2019-03-15曹慧颖张立军阮燕晔董小妹张馨悦
曹慧颖,张立军,阮燕晔,张 敖,董小妹,张馨悦
(沈阳农业大学生物科学技术学院 / 辽宁省植物基因工程技术研究中心, 辽宁 沈阳110866)
改革开放以来,中国经济持续高速发展,目前已经成为仅次于美国的第二大经济体[1]。经济的发展伴随着能源的巨大消耗,但我国的能源分布情况是富煤、贫油、少气。从1997年开始,中国成为能源的纯进口国,1998年石油进口量为0.55亿t,而 2015年石油进口量升至3.36亿t[2],石油进口量正以惊人的速度攀升。能源不足,是困扰中国经济发展的重要问题。除了进口,科学家们正在努力从多个角度开发新能源,比如风能、太阳能、核能、水能、地热能。在生物领域,植物通过光合作用将大量能量储存在其生物质中,因此研究利用植物生物质能源是一个重要课题。柳枝稷(Panicum virgatum)也因此受到更多研究者的关注。
柳枝稷属于禾本科黍属,起源于北美。其根系发达,生产力高,水利用率高,同时环境适应性好,在干旱、贫瘠甚至是盐碱地都能生长[3-4]。研究发现,将柳枝稷的生物质与煤炭混燃发电,是生物质利用成本最为低廉的方式。将10%柳枝稷和90%煤炭混合具有较好的燃烧性能,SOx和NOx的排放量也相应降低[5]。储存在柳枝稷纤维素生物质中的多糖可以通过微生物发酵、高温分解或者气化转化为生物燃料,美国能源部已经把柳枝稷作为专用的能源作物[6-8],我国已经引种并且正在推广[9-10]。
由于柳枝稷木质素含量高,同时进一步提高抗逆性有利于其更好地服务于能源生产,因此有必要对柳枝稷进行适当的遗传育种改良[11]。但是柳枝稷具有自交不亲和及异源多倍体等遗传特性,应用传统育种方法难以改良[12]。基因工程技术是一种快速、有效的育种手段,可以克服传统育种方法的不足。本文从基因型、遗传转化方法及功能基因改良等方面综述能源作物柳枝稷的基因工程研究进展,以期为培育更加优良的柳枝稷品种、提供新能源材料奠定基础。
1 柳枝稷遗传转化影响因素
1.1 转化技术的影响
植物基因工程育种是一种快速、便捷的育种技术,可以克服传统育种周期长、不能实现超远缘杂交等缺点。但是要应用该技术,首先要建立高效的柳枝稷遗传转化体系。目前柳枝稷的转化主要采用农杆菌介导法。农杆菌能将自身染色体外Tumor-inducing (Ti)质粒的一段 DNA (T-DNA)转移到宿主植物细胞中,如果有外源基因插入到T-DNA中,也可以随着T-DNA的转移一起整合进入植物基因组[13]。农杆菌介导的遗传转化系统最早被应用在双子叶植物中,由于单子叶植物尤其是禾本科植物不是农杆菌的天然宿主,所以该系统在单子叶植物中的应用进展缓慢。之后随着人们对农杆菌侵染机制了解的深入,转化技术也有所改进完善,农杆菌介导的遗传转化首先在水稻(Oryza sativa)[14]和玉米(Zea mays)[15]中取得了突破性进展。与其他转基因技术相比, 农杆菌介导的遗传转化系统具有遗传稳定、拷贝数低、成本低和基因沉默现象少等优点[16-17]。
为了提高农杆菌介导的植物遗传转化效率,在实际工作中,研究人员开发出一些辅助措施,以增加外源DNA进入宿主细胞的几率。如使用超声波[18-19]、真空渗透[19]、热激、离心[20]、表面活性剂[21]、碳化硅纤维[22]处理外植体,以及共培养时使用半胱氨酸等巯基化合物[23-24]等。这些辅助技术也被应用在柳枝稷转化中。
Li等[25]在柳枝稷的转化中,首先由种子诱导愈伤组织,农杆菌侵染愈伤组织的过程中,进行真空减压。随后的共培养阶段,并不使用培养基,而是直接将愈伤组织放在一张滤纸上进行干燥处理。共培养之后,没有直接进入选择培养阶段,而是将愈伤培养在不含筛选抗生素的培养基上稍作休息,让外源基因复苏,再进行筛选。同时在愈伤诱导和选择阶段添加脯氨酸。通过这样的转化流程,48.5%的Alamo愈伤组织和90.3%的Performer愈伤组织可以检测出潮霉素抗性。笔者也通过真空渗透加干燥的处理方法,得到了40%以上的Alamo愈伤组织出苗率(数据未发表)。
有报道[26]称,农杆菌侵染前将愈伤组织置于3% 麦芽糖、300 μmol·L-1谷氨酰胺 (Gln)溶液中冰浴20 min,随后的转化中,也采取真空、干燥处理,结果表明,AlamoⅡ型愈伤组织转化效率可达72%,Performer转化率高达96%;即使是高地型品种Blackwell也获得成功转化,转化效率为5.6%。一些柳枝稷转化试验的结果如表1所列,可以看出,即使是同一个品种、不同实验室、不同转化方法获得的转化率也不尽相同。但值得注意的是,不同文献采用的转化率计算方法也不完全一致。
1.2 基因型对遗传转化的影响
柳枝稷因其原产地纬度不同,可以分为低地和高地生态型[33-34]。低地生态型起源于低纬度地区,喜好温暖湿润的气候环境,其株型高大,生物量高,但耐寒性差,主要品种有Alamo、Performer、Bomaster、Kanlow和Colony等。髙地生态型起源于较高纬度地区,其茎秆略微细矮,生长较慢,生物量较低,但抗旱、耐寒能力强,其品种有Blackwell、Cave-in-Rock、Trailblazer、Carthage和Shawnee等[35]。
柳枝稷染色体基数 x = 9,它的倍性从 2 n =18 到 2 n = 108 都有分布,最常见的倍性是 2 n = 4 x =36 和 2 n = 8 x = 72 两种[36-38]。由于基因型不同,不同柳枝稷品种的组培再生能力及遗传转化效率明显不同[39]。刘燕蓉和张万军[40]对11个不同生态型柳枝稷品种进行组织培养,并统计分析其愈伤组织形态及比例,结果表明,低地型品种Alamo和Performer的种子发芽率和愈伤诱导率均较低,但容易分化、色泽鲜亮、结构松脆的Ⅱ型愈伤组织比例高;虽然高地型品种Dacotach、Blackwell和Nebraska 28的愈伤诱导率和种子发芽率较高,但Ⅱ型愈伤组织较少,不易获得组培再生苗。
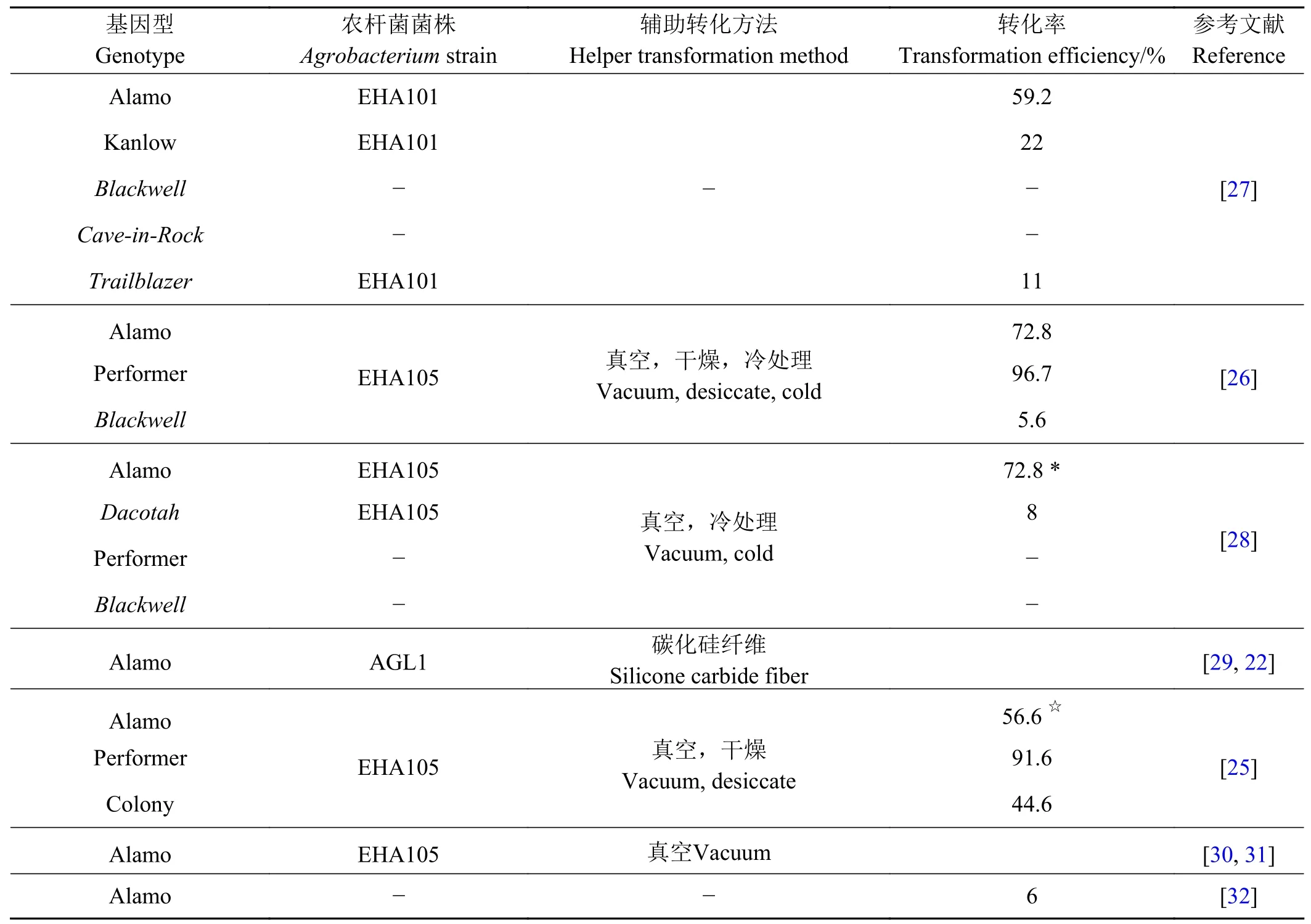
表1 柳枝稷遗传转化方法Table 1 The method of genetic transformation of switchgrass
Ogawa等[27]诱导柳枝稷种子形成结构致密、生长较慢的Ⅰ型愈伤组织,并利用农杆菌介导转化低地品种Alamo、Kanlow以及高地品种Trailblazer,但在该试验所采用条件下,高地品种Blackwell、Cave-in-Rock不能产生Ⅰ型愈伤组织。目前柳枝稷转基因研究常选用低地型品种,如Alamo。高地型品种由于基因型复杂等原因,愈伤组织再生及遗传转化均较低地型困难。
2 柳枝稷功能基因改良
2.1 降低木质素含量
植物生物质的主要成分为木质纤维素,包括半纤维素、纤维素和木质素等。木质素不但难以转化利用,而且还影响纤维素的生物降解。有研究指出,木质素含量为22%的原料可产生的糖只有木质素含量为17%的原料的一半,木质素含量为26%的原料几乎不产糖[41]。因此植物生物质必须经过预处理破坏木质素对纤维素的包裹[42],纤维素和半纤维素才可通过水解和发酵来生产乙醇。柳枝稷的木质素含量为17.56%,与常见的农业副产物玉米秸(18.59%)、麦秆(16.85%)木质素含量持平(表2)。如果采用生物技术适当降低木质素含量,就可以更好地利用柳枝稷的生物质[43]。
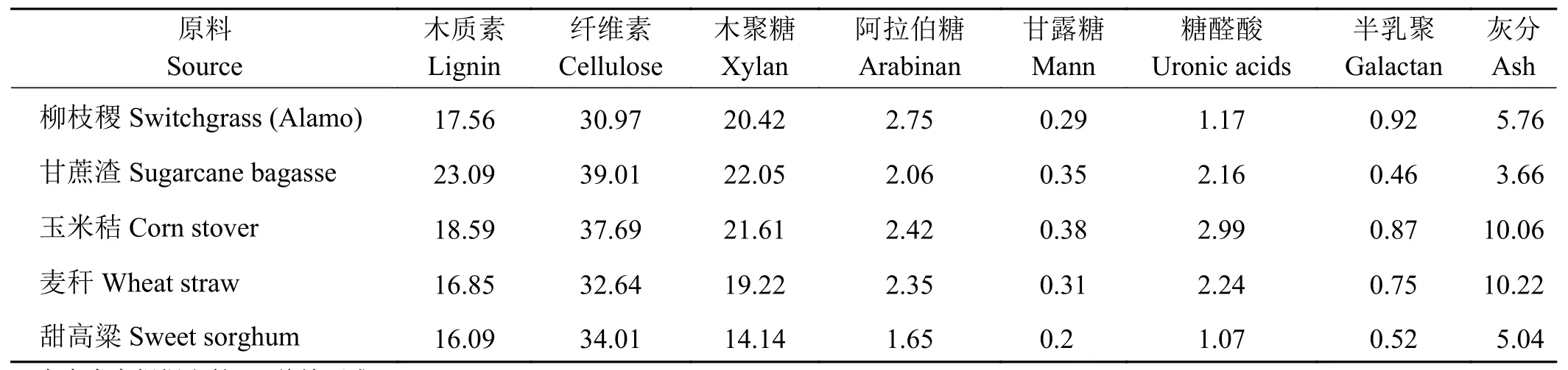
表2 一些常见能源植物的物质组成Table 2 Composition of some prospective energy crops
肉桂醇脱氢酶(CAD)是木质素合成途径的一个关键调控酶,它可以将木质素前体转化成木质素单体。已有报道通过下调柳枝稷CAD基因的表达,降低柳枝稷体内木质素的含量,从而使柳枝稷中糖化效率达到23%,并且不需要酸预处理[29, 44]。类似的可以降低木质素含量的酶还有咖啡酸-O-甲基转移酶(COMT)。木质素是由多种芳香醇单体形成的一种复杂酚类聚合物,COMT是参与苯丙酸途径中的一个甲基化酶。Fu等[45]下调了COMT基因的表达,在没有酸预处理的情况下,糖化效率增加了 29.2%~38.3%。然而有研究指出, 下调COMT的柳枝稷中低分子量酚类化合物会抑制糖化作用以及酿酒酵母的发酵[46]。另有研究表明,MYB家族转录因子调控植物木质素生物合成过程,对植物木质素含量及结构都起到了重要作用。Shen等[47-48]在柳枝稷中过表达了R2-R3-MYB转录因子家族的PvMYB4,在未经酸预处理的条件下就可以使糖化效率增加300%,最终使纤维素乙醇产量比普通植株提高了2.6倍。
2.2 提高生物量
MicroRNA(miRNA)是真核生物及病毒中存在的由19~24 nt组成的一类内源性非编码的单链小RNA,能够识别并指导靶基因的降解或者抑制靶基因的翻译。已有研究表明,miRNA广泛参与调控植物的生长发育、细胞分化、形态建成及激素代谢等各种过程[49]。miR156是植物中鉴定的第1个miRNA,在柳枝稷中miR156通过调控植物特有的一类多功能的转录因子家族SPL (SQUAMOSA promoter binding like)控制植物发育阶段。Fu 等[50]利用基因工程的方法过表达了miR156b前体基因,在较低的miR156表达水平下,柳枝稷可以正常开花,并且生物量增加了58%~101%。
蔗糖合成酶(SUS)催化蔗糖和UDP转变为UDP-葡萄糖和果糖,UDP-葡萄糖又可在纤维素合成蔗糖合成酶(SUS)催化蔗糖和UDP转变为UDP-葡萄糖和果糖,UDP-葡萄糖又可在纤维素合成酶作用下转变成组建细胞壁的纤维素。Poovaiah等[31]鉴定了4个柳枝稷SUS,过表达pvSUS1的转基因柳枝稷,植株高度增加37%,生物量增加13.6%。
2.3 抵抗逆境胁迫
自然环境中的植物,经常受到生物胁迫,比如病原菌的侵染。同时因其不可移动性,还要忍受干旱、洪涝、低温以及盐碱等非生物胁迫。研究发现,一些miRNA在抵抗逆境胁迫中发挥了重要的作用[51]。Matts等[52]利用生物信息学结合实验生物学的方法,鉴定了20个柳枝稷保守miRNA家族,预测了37个miRNA的靶基因。通过对miR395和miR399表达模式的分析,并且比较了它们在拟南芥(Arabidopsis thaliana)和蒺藜苜蓿(Medicago truncatula)中的表达,发现这两个miRNA与柳枝稷在贫瘠土壤中的生长适应性有关。
Xie等[53]对柳枝稷进行了干旱和盐处理,随后用测序的方法在该材料中鉴定出17个与干旱有关的 miRNAs,其中 4个是保守的 miRNAs,另外13个是柳枝稷特有的miRNAs。另有研究从柳枝稷中克隆了Pvi-MIR319a,并且用反向遗传学的方法研究了它的功能[54]。过表达Pvi-MIR319a的水稻表现出叶片宽大、植株矮小、开花延迟等表型,说明该基因对植株发育具有重要的调控作用,因此Pvi-MIR319a也被认为是柳枝稷遗传改良的良好候选基因。
多年生柳枝稷因其具有较高木质纤维素含量而成为生物能源生产的重要原料。如果通过代谢工程和基因工程进一步降低木质素含量,增加产量,提高抗胁迫能力,将使柳枝稷成为更为理想的能源作物。目前柳枝稷基因工程概况如表3所列。
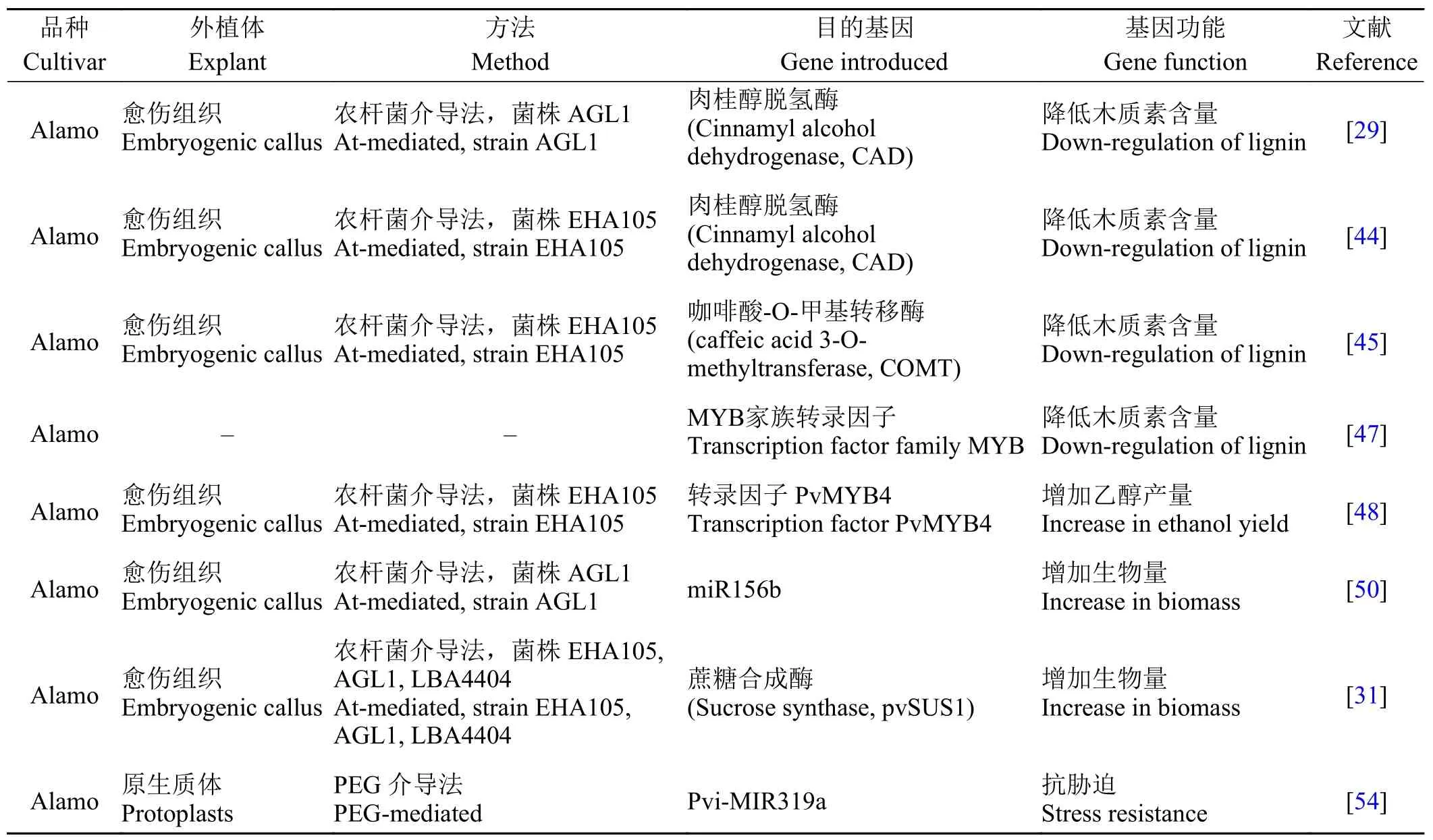
表3 柳枝稷基因工程概况Table 3 Summary of genetic modifications of switchgrass
3 研究展望
植物基因工程是把特定基因引入到宿主细胞,并整合到宿主基因组,从而改变宿主的遗传组成,产生具有特定性状的转基因植物。该技术是对传统育种方法的有益补充,也是改善作物品质、提高作物产量的重要手段。目前,转基因技术已经在大豆(Glycine max)、玉米、棉花(Gossypium hirsutum)、油菜(Brassica campestris)等植物中广泛应用,生产出了抗虫、抗病、抗除草剂等多个品种,产生了巨大的经济效益和社会效益。将基因工程技术应用到柳枝稷中,从而增加燃料乙醇的生产率,也必将成为柳枝稷育种的重要方面[55-56]。
柳枝稷的重要应用价值是生产纤维素乙醇,但实际生产中遇到了二代乙醇生产成本过高的问题。成本消耗在预处理、糖化、发酵等各个步骤。通过基因工程,改变柳枝稷结构组成,简化生产步骤,降低成本,可为开发经济可行的生产工艺奠定基础。可行的柳枝稷遗传工程改造包括降低木质素含量,改变细胞壁中糖的组成,从而使糖可以最大限度转化成为生物燃料。也有学者提出可以降低细胞壁交联来增加有效生物质[43]。所有这些工作都需要对柳枝稷进行系统的功能基因组学研究,详细了解各基因功能及其代谢调控,才能筛选出目的基因。Okada等[57]以柳枝稷Kanlow为母本、Alamo为父本进行杂交,杂交后代利用SSR标记绘制了遗传图谱。USDOE-JGI基于Roche 454和Illumina平台对柳枝稷开展了全基因组测序分析,致力于提供重要功能区域准确、全面的参考基因组信息,目前已经获得的序列信息公布于网站http://switchgrassgenomics.org/[58]。柳枝稷基因组研究的开展,有助于发现与细胞壁组成、作物产量和其他抗逆性相关的基因。科学家通过比较柳枝稷和水稻、高粱及其他植物的基因组数据,可以更好地理解柳枝稷基因组,为柳枝稷提供更多的生物燃料奠定坚实的研究基础。
