草鱼高脂日粮磷酸二氢钙适宜添加量的研究
2019-03-11李战福罗金强杨慧施陈拥军
李战福 罗金强 杨慧施 罗 浩, 李 玉 陈拥军 罗 莉
(1. 西南大学动物科技学院, 淡水鱼类资源与生殖发育教育部重点实验室, 重庆 400715;2. 北京师范大学统计学院, 北京 100875; 3. 新希望六和股份有限公司, 成都 610041)
磷是鱼类生长必需的常量矿物元素之一, 是骨骼、齿和鳞的组成成分, 能直接参与细胞中产生能量的各种反应, 与体内脂肪代谢有着强相关性[1, 2]。在脂类代谢过程中, 通常以磷脂的形式参与脂类物质和脂溶性维生素的代谢过程, 具有提高饲料中脂肪的氧化代谢速率, 加快营养物质的吸收, 促进鱼类生长的同时减少体内脂肪沉积的作用[3]。由于水体磷含量和鱼对饲料原料磷的利用率较低, 一般需在饲料中添加无机矿物磷来满足鱼类对磷的需求,以达到改善鱼体的生长及健康状况的目的。对于无胃的鲤科鱼类, MCP是最为高效和经济的磷源,且在草鱼饲料中添加磷酸二氢钙的效果优于其他无机磷源[3, 4]。
草鱼(Ctenopharyngodon idella)属鲤形目(Cypriniformes)、鲤科(Cyprinidae)、草鱼属(Ctenopharyngodon), 是我国重要的淡水养殖经济鱼类, 其养殖产量、饲料需求量占居第一位。随集约化高密度养殖的推广, 常通过增加饲料脂肪水平来达到提高养殖品种的生长速率、节约蛋白和提高饲料转化率的目的[5, 6]。而实际养殖生产中适量提高饲料脂肪水平是否意味着磷的添加量应适当的增加是一个值得探讨的问题。迄今, 国内外关于鱼类对磷需要量的研究已有大量报道, 主要淡水养殖品种的磷需要量为0.5%—1.57%, MCP的添加量为0.8%—3.6%[3, 7—10]。但因磷的添加量评定多采用纯化或半纯化日粮, 脂肪多设置在较低水平, 而在实际生产中饲喂的是较高脂肪水平的实用饲料, 其能量原料、蛋白原料和磷源矿物质原料可利用磷参数并不全面, 对磷的添加仍存在不确定性。目前, 唯有肖顺应[3]研究指出, 草鱼幼鱼饲料中MCP适宜添加量为1.2%。然而, 由于该研究中脂肪设置在较低水平(CL2.3%), 故草鱼实用高脂饲料中磷添加量需重新评定。为了使养殖鱼类达到较好的生长效果并尽量减少对环境的污染, 对草鱼高脂实用饲料中MCP的添加研究有很重要的意义。
鉴于此, 本研究在草鱼高脂(7.0%)日粮中分别添加0、1.0%、2.0%、3.0%和4.0%的MCP, 探讨其对草鱼生长性能、抗氧化及脂质代谢的影响, 确定草鱼高脂日粮中MCP的适宜添加量, 同时提供理论依据。
1 材料与方法
1.1 试验鱼及饲养管理
试验草鱼购于重庆市长寿鱼种场, 经3.0%浓度食盐水消毒后在室内养殖缸(392 L)中暂养, 用商品饲料(CP32%, CL5%, 重庆大发饲料有限公司)进行2周驯化。正式试验前, 选择体质健壮、体质量为(105.23±0.55) g的草鱼, 随机分为5个处理组, 每组3个重复, 每组15尾鱼, 日投喂量为体质量的2%—4%, 每天投喂3次(8:30、12:30、17:30), 生长试验为8周。据试验鱼的增重情况, 期间每隔10天调整1次投喂量。饲养期间水温为(22.0±2.5)℃, 溶氧量(6.5±0.4) mg/L, 氨氮<0.1 mg/L, 亚硝酸盐<0.1 mg/L,硫化氢<0.05 mg/L, pH为 (6.7±0.3)。
1.2 试验配方
本试验采用全植物蛋白饲料(豆粕、菜粕和棉粕)为蛋白源、大豆油作为脂肪源制成试验日粮,其组成及营养成分分析见表 1。日粮采用单因子浓度梯度法设计, 在基础饲粮中分别添加0 (对照)、1.0%、2.0%、3.0%和4.0%的MCP (四川川恒化工股份有限公司小太子牌, 磷含量19.09%, 实测), 分别表示为MCP0% (TP0.80%)组、MCP1.0%(TP1.02%)组、MCP2.0% (TP1.22%)组、MCP3.0%(TP1.44%)组和MCP4.0% (TP1.66%)组。为保持试验日粮中钙含量一致, 配方先经氯化钙(CaCl2)调平后用膨润土相应配平至100.00%, 配制成5种试验日粮, 饲料原料全部过40目筛, 采用逐级稀释法混合,挤压制成直径2.0 mm的颗粒, 风干分装, -20℃保存备用。
1.3 样品采集和分析方法
样品采集养殖试验结束后试验鱼停食24h,每组随机选取8尾鱼, 用MS-222进行麻醉后, 分别测其体高、体长、称重。取5尾鱼在冰盘上解剖,取出内脏团并分离肝胰脏和肠道, 内脏团和肝胰脏称重, 肠道用滤纸吸干表面水分, 剪碎混匀后用液氮速冻, 置于-80℃冰箱中用于消化酶测定。另3尾鱼静脉取血, 4℃静置3h后, 4℃冷冻离心(4000 r/min, 10min), 取血清-80℃保存备用, 用于血液生化指标的测定。
常规成分分析饲料原料及成品、全鱼、肌肉、肝脏常规成分的测定方法: 水分、粗蛋白、粗脂肪、粗灰分含量, 分别采用105℃烘箱干燥法(GB/T 6435—2006)、凯氏定氮法(GB/T 6432—1994)、索氏抽提法(GB/T 6433—1994)、550℃灼烧法(GB/T 6438—1992)进行测定; 磷的测定方法:饲料总磷(GB/T 6437—2002)。
生长及形体指标测定准确称取每缸鱼体质量和剩余饲料, 各生长及形体指标计算公式如下:(Wt-W0);


表 1 实验日粮配方及其营养组成(风干样基础)Tab. 1 Formulation and proximate analysis of experimental diets(air-dry basis) (%)
内脏指数(Viscerosomatic index,VSI, %)=Wv/W×100%;
肥满度(Condition factor,CF, g/cm3)=W/L3×100%;
式中:W0为初始鱼尾重(g);Wt为终末鱼尾重(g);F为尾均摄食量(g);L为鱼体长(cm);W为鱼体重(g);Wv为内脏重(g)。
消化酶及血液生化指标的测定肠道淀粉酶(Amylase)、脂肪酶(Lipase)、胰蛋白酶(Trypsin),血清总胆固醇(TC)、甘油三酯(TG)、总抗氧化能力(T-AOC) 、碱性磷酸酶(AKP)、谷丙转氨酶(ALT), 肝脏丙二醛(MDA)等指标均采用试剂盒(南京建成生物工程研究所) 并按照规定方法测定。肝脏肉碱脂酰转移酶(CPT-Ⅰ)和乙酰辅酶A羧化酶(ACC)采用试剂盒(上海江莱生物科技有限公司) 并按照规定方法测定。
1.4 数据处理
试验数据均采用统计软件SPSS 22.0分析。经过单因素方差分析(One-way ANOVA)之后, 采用Duncan’s多重比较,P<0.05表示差异显著。实验数据均用平以均值±标准差表示, 折线回归模型[Y=L-U(R-XLR),XLR表示X<R, 当X>R时, (R-XLR)=0][12]来分析饲料中MCP的适宜添加量。
2 结果
2.1 高脂日粮中MCP添加水平对草鱼生长性能和形体指标的影响
随日粮中MCP添加水平的增加, 草鱼FBW与WGR显著上升, 其中3.0%组WGR比对照组升高了135.40% (P<0.05), 当添加量大于3.0%后差异不显著(P>0.05), 而FCR和VSI逐渐下降, 分别在3.0%和4.0%组达到了最小值,而 4.0%组VSI较0组降低了23.06%。2.0%和3.0%组CF显著高于其他组(P<0.05, 表 2)。
采用折线模型拟合WGR(Y1)、FCR(Y2)和MCP添加水平(X)之间的关系, 分别得到高脂日粮中满足草鱼最佳生长需求和最低FCR的MCP添加量为3.26% [Y1=92.4-17.579 (3.26-X),R2=0.948]和2.96%[Y2=1.54+0.307 (2.96-X),R2=0.950](图 1、图 2)。
2.2 在高脂日粮中MCP添加水平对草鱼营养组成的影响
随着日粮中MCP添加水平增加, 草鱼全鱼粗蛋白和肌肉粗蛋白显著上升, 在3.0%组水平达到最大值(P<0.05), 而全鱼、内脏、肝脏和肌肉粗脂肪随日粮中磷含量增加而显著下降, 在3.0%组达到最小值(P<0.05), 较0组降低了13.71%。全鱼粗灰分在0—3%水平组有增加趋势但差异不显著(P>0.05);而全鱼总磷在0—2.0%水平组逐渐升高, 在2.0%—4.0%水平组差异不显著(P>0.05, 表 3)。
采用折线模型拟合全鱼磷含量(Y3) 和MCP添加水平(X)之间的关系, 得到满足草鱼对磷最大沉积需要的饲料MCP最低添加量为2.63%[Y4=1.83-0.228 (2.63-X),R2=0.925 (图 3)]。
2.3 在高脂日粮中MCP添加水平对草鱼血清生化指标的影响
日粮中MCP添加水平对草鱼血清TG、TC、AKP和ALT均有显著影响(P<0.05)。其中MCP添加组中血清TG显著低于对照组(P<0.05), 但MCP添加组之间差异不显著(P>0.05)。而血清TC随日粮中添加磷的增加显著下降, 添加量大于3.0%后急剧上升, 其血清TG和TC均在3.0%组达到最小值。当MCP添加水平从0上升至4.0%时, 血清AKP活性先增后降, 3.0%组最高, 较0组上升了100.66%。而血清ALT活性与血清AKP活性相反, 其中3.0%组显著低于其他组(P<0.05), 较0组下降了47.83% (表 4)。
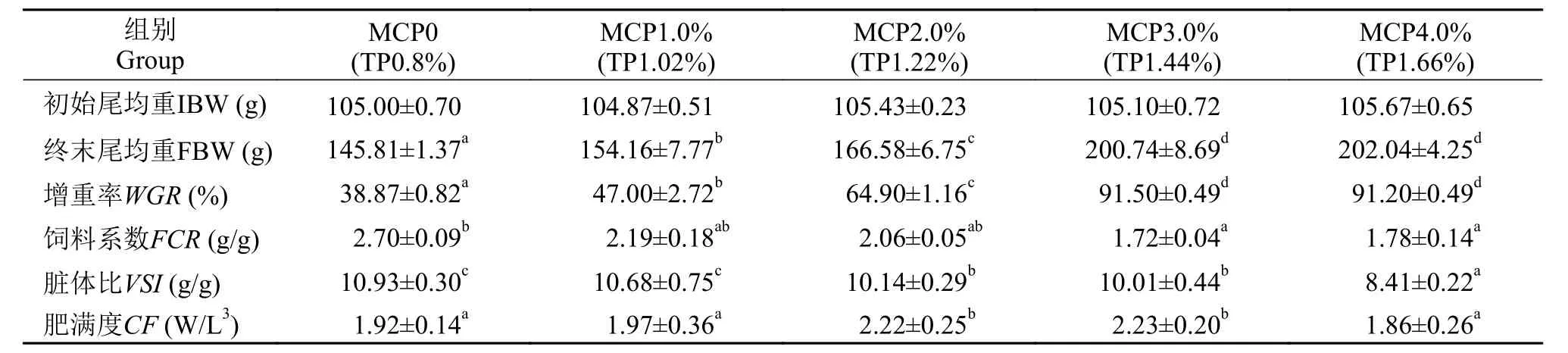
表 2 在高脂日粮水平下MCP添加水平对草鱼生长性能和形体指标的影响Tab. 2 Effects of different MCP levels on the growth performance and body index of grass carp with high lipid diet
2.4 在高脂日粮中MCP添加水平对草鱼消化及抗氧化能力的影响
日粮MCP添加水平对草鱼肠道消化酶活性及抗氧化能力均有显著影响(P<0.05)。肠道淀粉酶和脂肪酶活性随日粮MCP添加水平从0上升至4.0%时显著上升, 均在4.0%水平达到最高(P<0.05), 与0组相比分别升高了270.21%和70.77%, 而肠道胰蛋白酶活性呈先上升后下降趋势, 最大值出现在3.0%,较0组升高了8.42% (P<0.05)。这说明日粮中添加适宜的MCP对淀粉和脂肪的消化能力的提升远高于蛋白质。当MCP添加水平从0上升到4.0%时, 血清T-AOC含量呈上升趋势, 在4.0%水平达到最高, 较0组升高了175.90%, 而肝脏MDA含量则呈先下降后上升的趋势, 在3.0%水平达到最小值(P<0.05, 表 5)。
2.5 在高脂日粮中MCP添加水平对草鱼肝脏脂质代谢的影响
在日粮中添加不同水平MCP对草鱼肝脏脂质代谢酶活性有显著影响(P<0.05)。当MCP添加水平从0上升至4.0%时, 肝脏CPT-1活性呈上升的趋势,在4.0%组达到了最大值(P<0.05), 较0组升高了30.80%。肝脏ACC活性呈先下降后上升的趋势, 在3.0%达到最小值(P<0.05), 较0组下降了36.66%。随日粮磷添加量的增加, 对肝脏CPT-I/ACC活力比值有显著影响(P<0.05), 在0—4.0%时CPT-I/ACC比值先增后降, 在MCP3.0%组达到最大 (表 6)。
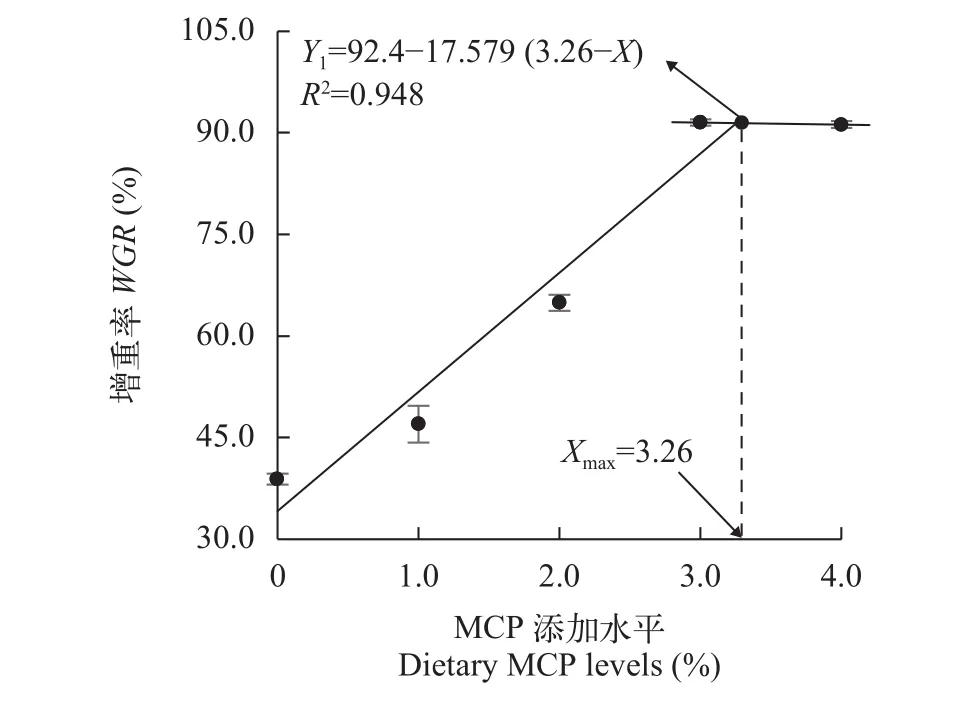
图 1 MCP添加水平对增重率的影响Fig. 1 Effects of dietary MCP levels on the WGR
2.6 日粮MCP添加水平相关指标间的相关性分析
将随着日粮MCP水平的变化的指标2(增重率,全鱼、内脏、肝脏和肌肉粗脂肪, 肠道脂肪酶)与指标1(MCP水平) , 指标4 (TC、TG、CPT-I/ACC、CPT-I和ACC)与指标3 (全鱼和肝脏粗脂肪)之间进行相关性分析后, 发现全鱼、内脏和肌肉脂肪,FCR与MCP水平, TC、TG和与全鱼粗脂肪均呈显著二次曲线相关(P<0.05);WGR、肠道脂肪酶活性与MCP水平和ACC活性与肝脏粗脂肪呈显著正相关, CPT-I/ACC比值与全鱼粗脂肪呈显著负相关(P<0.05); 而肝脏粗脂肪与MCP、CPT-I与肝脏粗脂肪存在一定的线性相关关系(P>0.05, 表 7)。
3 讨论
3.1 高脂日粮添加适宜MCP对草鱼生长性能的改善
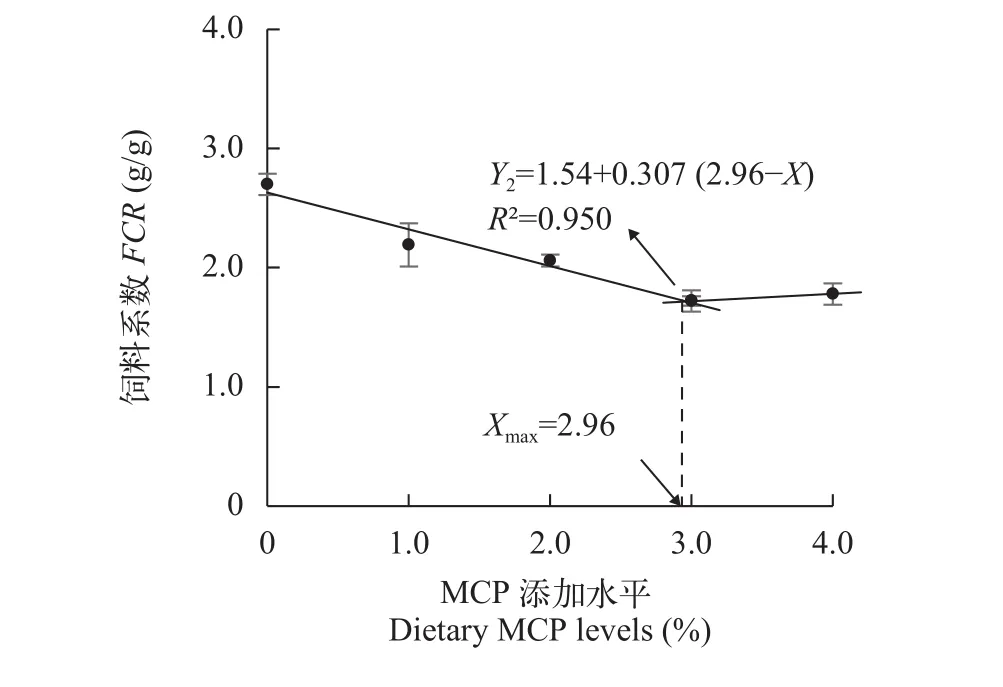
图 2 MCP添加水平对饲料系数的影响Fig. 2 Effects of dietary MCP levels on the FCR
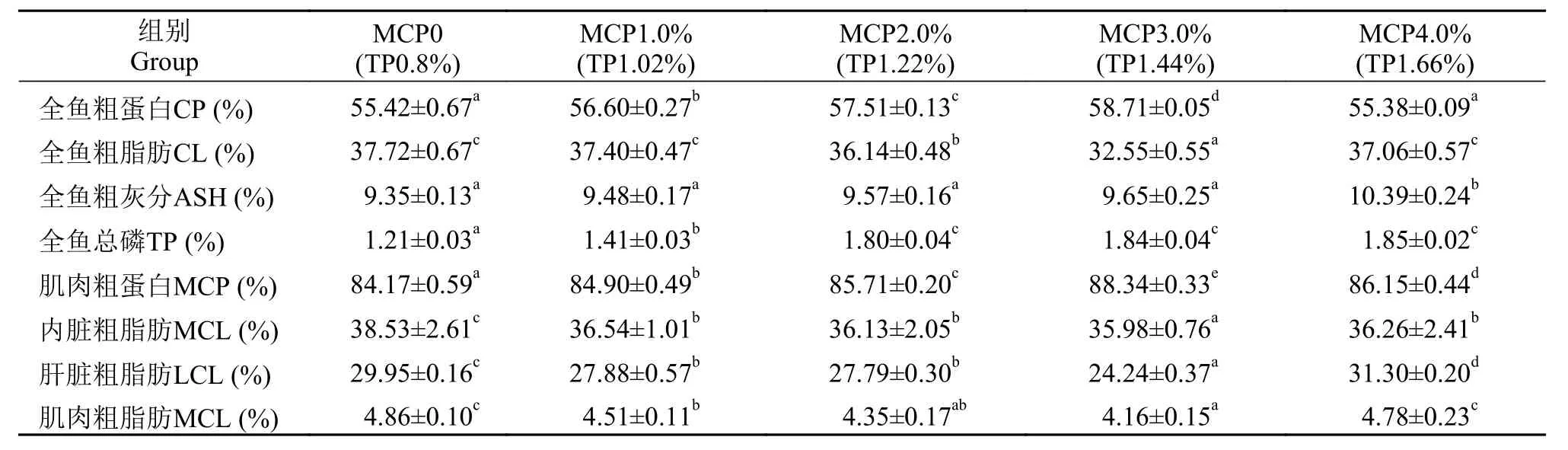
表 3 在高脂日粮水平下MCP添加水平对草鱼营养组成的影响(干物质基础)Tab. 3 Effects of different MCP levels on the nutrient composition of grass carp with high lipid diet (dry matter basis)
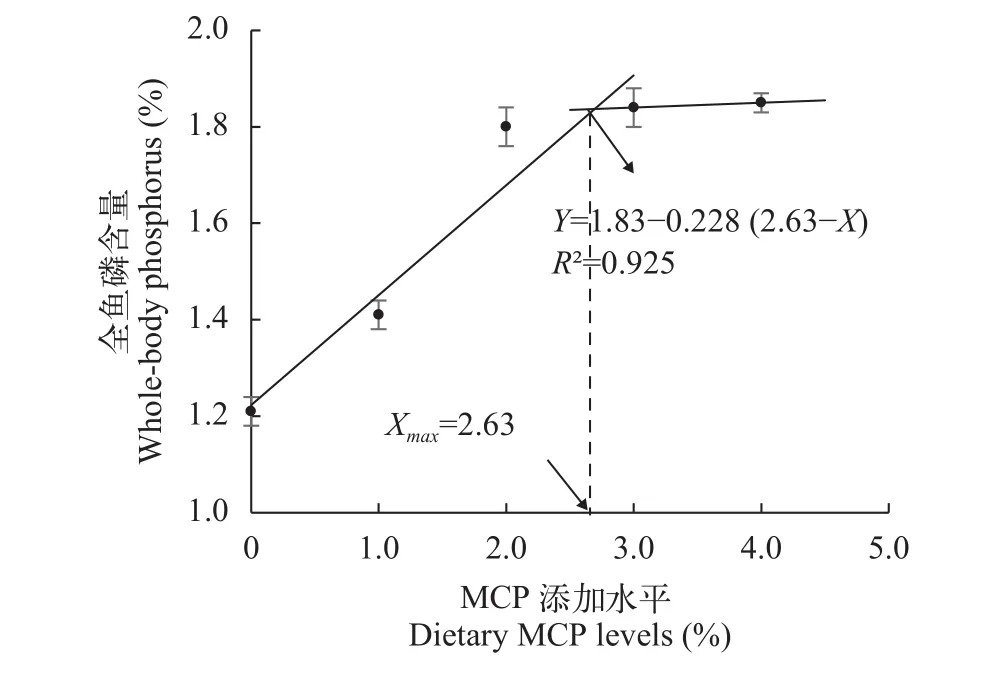
图 3 MCP添加水平对全鱼磷含量的影响Fig. 3 Effects of dietary MCP levels on whole-body phosphorus
磷是鱼类营养的重要元素, 摄食含磷量不同的饲料会导致鱼体生长性能及饲料效率发生变化[3]。磷添加不足会造成水产动物生长速率缓慢、饲料利用率下降和骨骼发育不良, 而过量又会导致鱼类代谢紊乱、增加饲料成本和污染水体等[7, 8, 11]。本实验结果表明, 磷的添加可显著影响草鱼的生长性能, 除此之外, 磷添加过低还表现在体脂肪和组织脂肪蓄积, 消化能力下降以及机体抗氧化能力降低等, 但未出现骨骼发育异常等缺乏症状, 这可能是由于基础原料中所含磷能够满足草鱼基本生长需要和骨骼的正常发育, 而磷添加过量会导致鱼类代谢紊乱增加脂肪沉积和饲料成本。评价鱼类饲料中磷适宜添加量的指标很多, 并且不同的指标评定出的结果有所差异。为使养殖鱼类达到较好生长效果并尽量减少对环境的污染, 本研究以WGR和FCR为观测指标, 折线模型分析得日粮MCP适宜添加水平为2.96%—3.26% (总磷TP为1.43%—1.50%)。这一结果与Yuan等[13]胭脂鱼(Myxocyprinus asiaticus)结果相近, 而高于肖顺应[3]研究的草鱼(1.2%)和湘云鲫(Triploid crucian carp)[14]、建鲤[10](Cyprinus carpiovar. Jian)、鲤(Cyprinus carpioL.)[15]、吉罗罗非鱼(Oreochromis niloticus)[16]等。本研究相较于前人对草鱼磷需求量研究所得结果要高, 其原因可能是与本试验磷添加量评定采用的是高脂实用日料, 而前人研究多采用纯化或半纯化日粮, 脂肪多设置在较低水平有关, 这也进一步说明了适量提高日粮脂肪水平, 磷的添加量应适当的增加, 可见日粮磷同样影响草鱼的脂肪代谢。不同鱼类最适磷添加量的差异可能是由于种类、规格、生长阶段、日粮构成、磷源以及养殖环境等不同所致[17]。
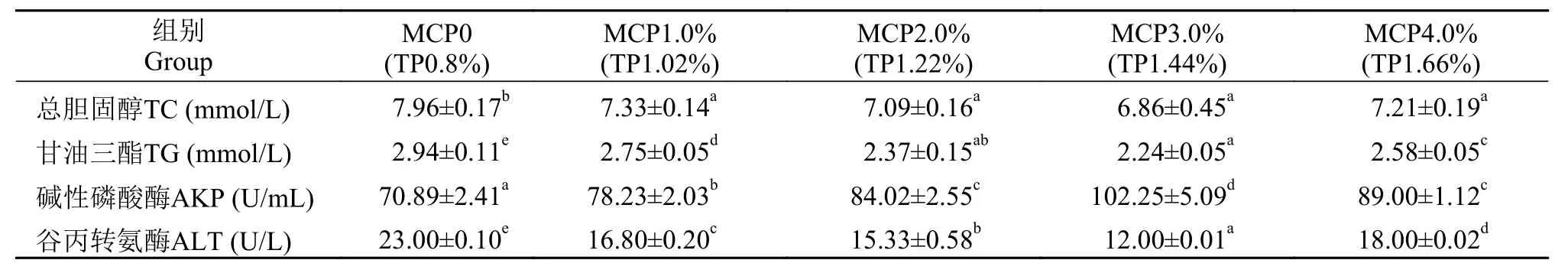
表 4 在高脂日粮水平下添加不同MCP对草鱼血清生化指标的影响Tab. 4 Effects of different MCP levels on grass carp serum biochemical index with high lipid diet
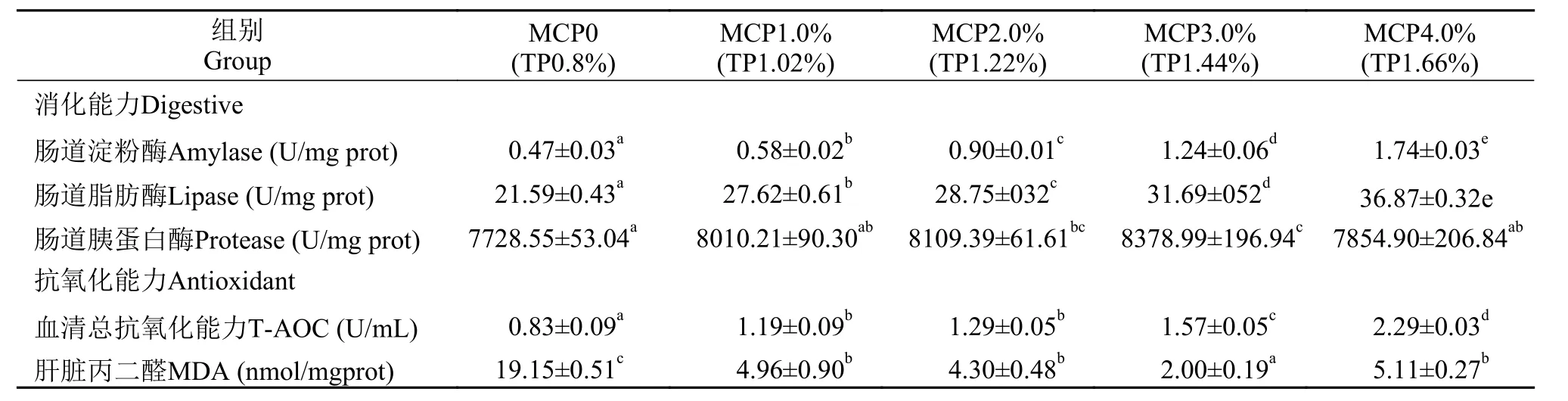
表 5 在高脂日粮水平下添加MCP对草鱼消化及抗氧化能力的影响Tab. 5 Effects of different MCP levels on intestinal digestive and antioxidant ability of grass carp with high lipid diet
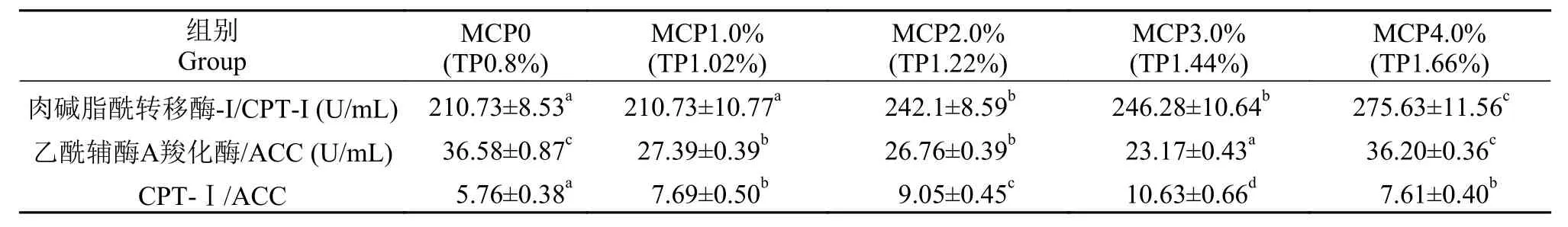
表 6 高脂日粮水平下添加MCP对草鱼肝脏脂质代谢酶活性的影响Tab. 6 Effect of different MCP levels on lipid metabolism enzymatic activity of liver of grass carp with high lipid diet
3.2 高脂日粮添加适宜MCP对草鱼脂质消化、代谢及沉积的影响
关于磷的添加对鱼类脂肪消化吸收、肝脏脂肪酸合成与分解代谢及沉积的影响研究已有部分报道。肝胰脏是鱼类进行脂肪代谢和调节脂肪蓄积的主要器官, 也是生成脂肪酶的主要器官, 而肠道是无胃鱼主要的消化、吸收营养物质的场所。脂肪的运输主要借助血清, 血清中的总胆固醇(TC)和甘油三酯(TG)的含量在一定程度上代表着全身的脂类代谢程度[18—20]。本研究表明, 在高脂日粮中添加适宜的磷时显著提高了草鱼肠道脂肪酶的活性, 促进了脂质的消化, 降低了草鱼血清TC和TG, 与EI-Zibdeh等[19]在大黄鱼中的研究结果一致,大黄鱼血清中TC和TG含量随饲料中磷水平的升高而降低, 表明在饲料中适量磷的添加有利于改善草鱼的肝脏脂肪代谢。肉碱脂酰转移酶-Ⅰ(CPT-Ⅰ)是线粒体肉碱转运系统重要组成酶, 是脂肪酸分解酶中的关键限速酶, 在能量代谢中起着重要作用, 长链脂肪酸及其辅酶A只有通过该转运系统才能进入线粒体基质进行β氧化, 而乙酰辅酶A羧化酶(ACC)是脂肪酸从头合成途径的关键限速酶。Sugiura等[21]在虹鳟的研究中发现, 饲喂高低2种不同磷水平的饲料, 肝脏CPT-I表达量没有显著差异, 但是肝脏ACC表达量在高磷饲料组显著下降。石亚庆[14]在湘云鲫的研究发现, 肝脏ACC活性随饲料MCP添加水平的升高呈先降低后上升的趋势。本试验肝脏ACC活性呈先下降(MCP0—3.0%)后上升(MCP3.0%—4.0%)趋势, 而肝脏CPT-1活性随磷添加水平的增加逐渐升高, 其中MCP3.0%组CPT-I与ACC比值最大, 说明添加量脂肪酸的分解代谢速率高于脂肪酸合成速率, 使得草鱼肝脂下降。Takeuchit等[22]认为发生变化的原因可能是因为鱼体摄取饲料磷的数量与体内脂肪的氧化速度存在联系有关,Roy等[23]认为可能是因为摄食高磷水平饲料的鱼体内, 脂肪酸的氧化增强, 糖原生成增加, 导致蛋白沉积的增加和脂肪沉积的降低。
鱼类摄食含磷量不同的日粮会直接影响鱼体常规成分。在本试验日粮中磷的添加与脂肪沉积关系表现在添加量不足或过量组中全鱼、内脏、肝脏和肌肉中脂肪含量均显著高于适宜添加组(P<0.05)。Eya等[24]研究发现, 斑点叉尾鮰肌肉中脂肪含量与饲料中磷含量之间呈负相关关系, 而蛋白含量与饲料磷含量之间呈正相关关系。这也进一步验证在日粮中添加适宜磷可显著减少脂肪沉积。综上, 在日粮中添加适宜的MCP可以促进草鱼肠道脂肪消化吸收, 加快肝脏脂肪酸合成与分解代谢速率, 从而减少鱼体脂肪沉积, 其深入机理有待进一步探讨。
3.3 高脂日粮添加适宜MCP对草鱼抗氧化能力的影响
需氧生物体在糖脂代谢过程中会产生大量自由基(Free radical, FR), 线粒体呼吸链电子漏生成的超氧阴离子是有机体活性氧的主要和恒定来源, 其中作用最广的氧自由基有超氧自由基羟自由基(OH·)和过氧化氢(H2O2)[19, 25]。自由基的过度产生是机体过氧化损伤、代谢紊乱和抗氧化失衡的主要原因。磷脂是构成生物膜的重要部分, 因富含多不饱和脂肪酸故极易受自由基破坏, 膜中磷脂发生过氧化作用, 会引起膜中蛋白质及酶的交联或失活, 导致膜通透性的变化, 严重影响膜的各种生理功能。若细胞内线粒体膜被氧化受损, 则会使能量生成系统受到影响, 从而引起细胞功能极大紊乱[20, 26]。总抗氧化能力(T-AOC)是一个用于衡量生物机体抗氧化系统状况的综合性指标, 它可以在一定程度上反映出机体自由基代谢的情况。丙二醛(MDA)是脂质过氧化反应的最终代谢产物, 其含量的高低可以间接反映机体脂质过氧化水平。李安林等[27]研究表明高脂饲料磷添加不足可导致大鼠T-AOC显著降低, MDA含量显著升高。而本研究在高脂日粮中添加适宜磷后显著升高草鱼T-AOC,降低肝脏中MDA含量, 这与Feng等[28]和Tang等[29]分别在磷对建鲤和鲶鱼的抗氧化能力影响的研究得出的结果相似。可能与磷是构成细胞膜磷脂的主要成分, 在饲料中添加适量的磷能保证细胞膜的完整性, 降低一些超氧化物的侵袭有关。碱性磷酸酶(AKP)是磷代谢过程中的调控酶, 而谷丙转氨酶(ALT) 是反映肝细胞受损程度最灵敏的指标, 在肝细胞受到损伤时, ALT释放入血液中, 会引起血清ALT偏高。本试验表明在MCP3%组时, AKP活性最高, 而ALT活性最低, 这与陈任孝[30]在吉富罗非鱼上的研究结果相似, 说明在日粮中添加适量MCP能促进AKP活性, 进而增强磷代谢利用以及对肝脏有一定的保护作用。AKP作为磷代谢的重要酶, 它受日粮等因素影响的变化范围有待进一步的确定, 为后续的研究提供更多的依据。
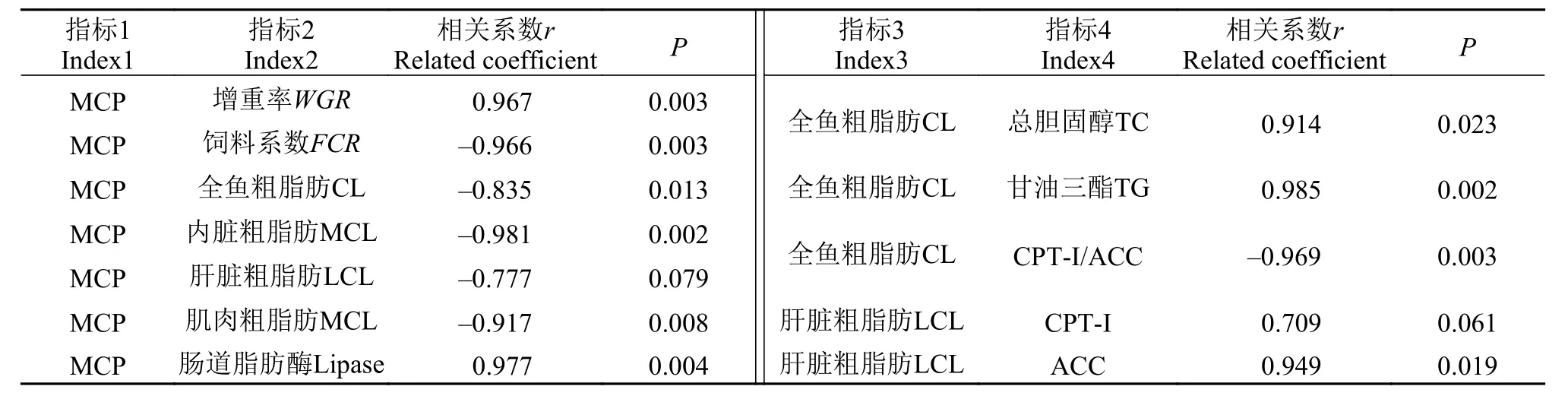
表 7 高脂日粮MCP添加水平与体组织脂肪及脂质消化和代谢指标间的相关性分析Tab. 7 The correlations between dietary MCP supplementation and body, tissue fat, lipid digestion and metabolism indices
4 结论
在本试验条件下, 得出草鱼日粮在脂肪水平为7.0%时磷酸二氢钙(MCP)适宜添加量为2.96%—3.26%, 即饲料总磷量为1.43%—1.50%。该添加水平能提高鱼体对饲料中淀粉和脂肪的消化吸收, 加快营养物质的吸收, 增加糖原生成, 减少体内脂肪蓄积, 并提高抗氧化能力, 从而促进鱼体生长。
