菊花在形态、组织结构和基因表达方面对弱光的响应
2019-03-11朱晓琴裴冬丽
韩 霜,朱晓琴,裴冬丽
(商丘师范学院生物与食品学院,植物与微生物互作河南省重点实验室,河南商丘476000)
植物对弱光一般有两种反应:服从或逃避。一方面是通过提高叶面积降低叶绿素a/b(Chl a/b)比率来适应弱光,另一方面是通过改变叶片与水平面的角度以便捕获更多光子来适应弱光[1]。弱光条件下光合速率受抑制,从而影响植物生长和产量[2],同时还会引起叶面积增大,叶绿素含量增加[3]。
光对植物的生长、发育、形态、生理,乃至于演化都具有极其重要的作用,而作为同化器官的叶片在弱光环境中其形态及解剖结构发生适应性变化,这些在辣椒[4]、茉莉[5]和菊花[6]等多种植物上已有研究报道。淀粉体是白色体的一种,是贮藏光合产物淀粉的一个重要场所,所以它的多少能够反映光合效率的大小[7]。叶绿体是行使光合作用的细胞器,最易受到伤害,短期弱光促使基粒片层加厚以适应弱光[8]。随着弱光处理时间的延长,叶绿体膨胀变大内部出现裂缝和孔洞,基质片层肿胀[5],通过分析马铃薯[9]、花生[10]等物种耐弱光与超微结构的关系发现,耐弱光性强的种或品种超微结构受伤害程度较小。一般认为,1-氨基环丙烷-1-羧酸氧化酶是植物体内乙烯生物合成的关键酶,其活性大小决定着乙烯的生物量[11]。一些研究认为乙烯伴随着庇阴症状产生,而且低浓度的乙烯引起弱光条件下西红柿茎伸长[12]。光系统Ⅱ捕光色素蛋白包括6种蛋白质,Lhcb1、Lhcb2和Lhcb3基因编码三聚体多肽为LHCII蛋白[13],前期研究发现Lhcb1基因在转录水平上受光的调控[14]。
菊花品种较多,对光照的适应性差异明显,本研究以清露和公子2个对弱光响应差异的2个品种为试验材料,设置自然光照和重度遮阴2个处理,分析这2个菊花在形态、组织结构、超微结构和基因等方面对弱光响应差异,为探索光能高效利用机制提供依据,为设施菊花生产提供理论依据。
1 材料和方法
1.1 生长条件和遮阴处理
供试材料为弱光敏感品种清露和非敏感品种公子,由南京农业大学中国菊花种质资料保存中心提供。选择生长一致的插穗扦插,生根后移植到口径14 cm大小的花盆中,所用基质配方为营养土∶珍珠岩∶蛭石=1∶1∶1,不施营养液。缓苗后,随机分组,光照度分别是:自然光照(对照,中午12时平均光合有效辐射830~1 260μmol/m2s)和20%自然光照(弱光处理160~260μmol/m2s),每个处理60盆。试验期间,温度、光周期和相对湿度分别是:28/15℃,13/11 h和70%。所有植株接受自然降水,需要水分时灌溉,自然光照条件2 d一次,遮阴条件下每3~4天一次,分别在0、1、2、3和4 d上午11:00取样,处理20 d后出现明显的节间变长、叶柄变长、叶片变薄变大等庇阴症状时取样。
1.2 节间长、叶柄长、叶面积和比叶重
每个处理5株,从上往下数第4片完全展开叶,测量4~5叶片的节间,然后把叶片平展放到信封中,LI-3100C叶面积仪(LI-COR,Lincoln,NE,USA)测定叶面积后放到80℃烘箱中,烘干称重。比叶重(SLW,g/m2)=干重/叶面积。
1.3 茎剖面结构和叶片超微结构
石蜡切片制作参照李正理[15]的方法。取叶脉两侧部位5 mm×1 mm大小的样品,用2.5%戊二醛抽气4~5 min进行固定,然后送电镜室完成缓冲液冲洗、锇酸固定、乙醇梯度脱水、丙酮置换渗透、包埋、染色等步骤[16],最后选择典型视野观察、拍照。
1.4 CmLhcb2和CmACO2基因的表达
根据菊花脑转录组信息,叶绿素a/b结合蛋白基因Lhcb2、1-氨基环丙烷-1-羧酸氧化酶基因ACO2特异引物和定量内参引物GAPDH,引物序列见表1。

表1 用于荧光定量PCR分析的基因引物序列Table 1 Primer sequence used for real-time quantitative PCR analysis
1.5 统计分析
结果用平均值±标准误表示。利用SPSS v17.0(SPSS Inc.,Chicago,IL,USA)进行统计分析和双因素分析,P<0.05。
2 结果与分析
2.1 植物生长发育和形态
光照强度影响植物生长和叶片形态,20 d时,弱光条件下,出现节间变长、叶柄变长、叶片变薄变大等庇阴症状(见表2)。
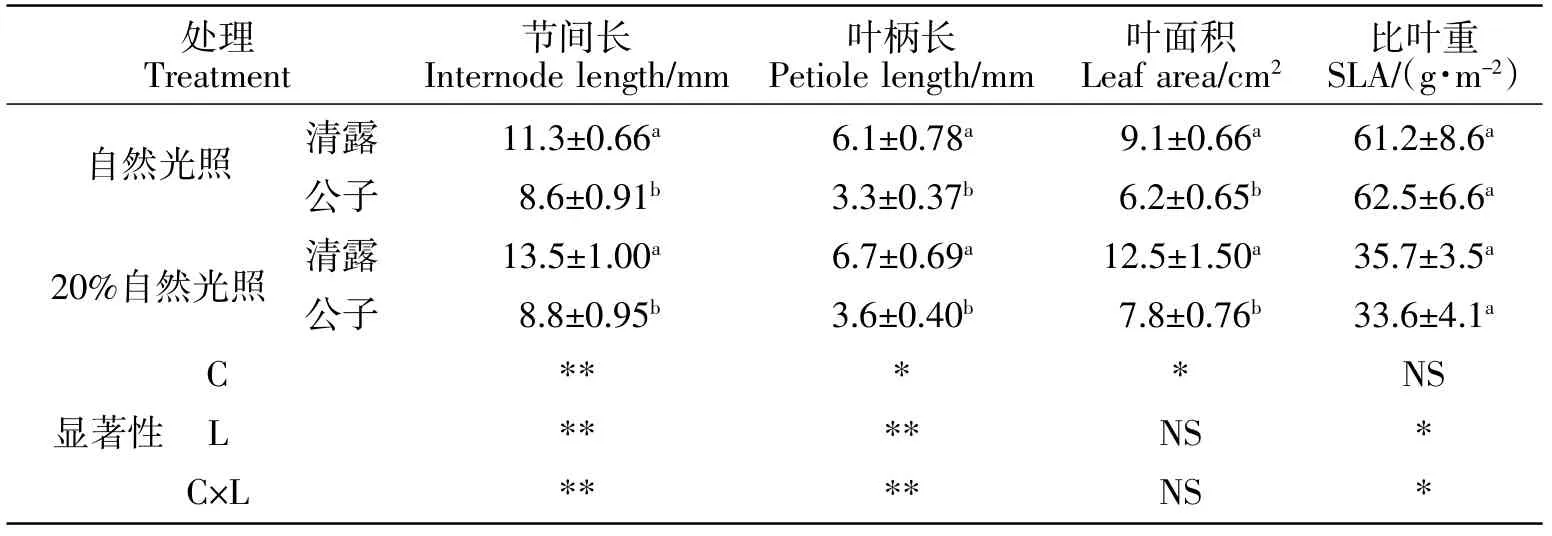
表2 不同菊花品种在自然光照、20%自然光照条件下处理20 d节间长、叶柄长和叶面积Table 2 The length of the stem,internode and petiole,and the leaf area of different Chrysanthemum cultivars exposed to full sunlight and 20% irradiance for 20 days
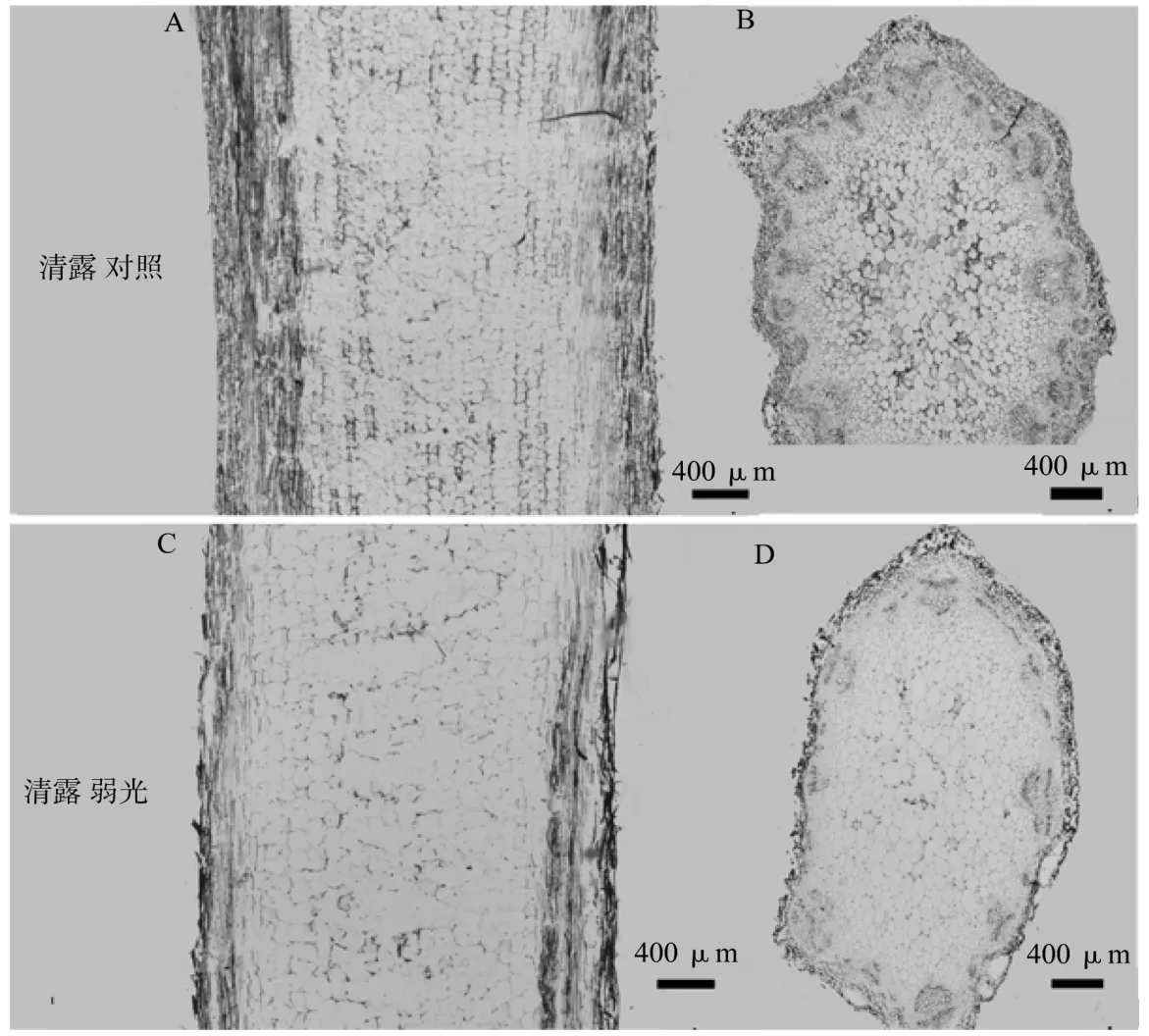
图1 菊花品种清露解剖结构Figure 1 The anatomical structure of Chrysanthemum cultivar Puma Sunny
2.2 茎解剖结构和叶绿体超微结构
2个品种的茎均由表皮、皮层、维管束和初生木质部4部分组成(图1、图2),自然光照条件下细胞形状规则、排列紧密,弱光条件下清露细胞呈不规则变形,皮层以内的维管束细胞更为明显(图1C,1D),公子受到的影响较小(图2)。20%光照使清露叶肉细胞内淀粉粒减少,叶绿体变长(图3B),基质片层排列松弛(图3D);公子叶肉细胞内淀粉粒受到的影响较小,叶绿体长度变化不明显(图4B),基粒片层较规则(图4D)。

图2 菊花品种公子解剖结构Figure 2 The anatomical structure of Chrysanthemum cultivar Gongzi

图3 清露在不同光照条件下叶绿体超微结构Figure 3 The chloroplast ultrastructure of Chrysanthemum cultivar Puma sunny under different light conditions

图4 公子在不同光照条件下叶绿体超微结构Figure 4 The chloroplast ultrastructure of Chrysanthemum cultivar Gongzi under different light conditions
2.3 CmLhcb2和CmACO2基因的表达
在弱光条件下,清露和公子CmLhcb2均有不同程度的上调。20%光照条件下,清露在0~4 d快速上调,第2天时达显著水平,3 d时比自然光照条件下高1.57倍,随后下降(图5A),公子CmLhcb2表达量在0~4 d时逐步上升,弱光处理显著高于对照,第3天时达显著水平(图5B);弱光条件下,清露CmACO2在第1天显著增加,2~4 d缓慢下降(图5C),公子的CmACO2表达量在0~4 d时逐步升高(图5D)。因此,清露叶片CmLhcb2和CmACO2基因表达对弱光的响应比公子更快,清露更敏感。
3 讨论与结论
遮阴使植物形态、组织结构和基因表达发生变化[5]。但菊花对庇阴的响应到目前没有报道。以前的研究认为,遮阴适应型花叶万年青通过扩大叶面积来调整光合适应性[17],低光照能促使干物质运输分配用于扩大叶面积[18]。本研究发现弱光条件下,清露出现节间长、叶柄和叶面积增加等庇阴症状,公子不明显,此结论与邹长明等[19]关于不同品种小豆对弱光的响应差异相同,弱光敏感型受到的影响更大。本课题组前期研究发现弱光改变了叶脉筛管-伴胞复合体的结构,使同化物的运输受阻,清露受到的影响比公子显著[20]。本研究发现弱光引起表皮细胞和皮层细胞呈现不规则变形,维管束细胞更为明显,这个结论印证了弱光引起物质运输不畅,从而导致根冠比下降的结论。
光合反应发生在叶绿体层膜系统类囊体上,所以类囊体结构决定着光合活性的高低[7]。试验结果显示,自然光照下叶绿体结构发育良好,基粒片层和基质片层排列整齐(图3A、C)。同时20%弱光条件下,清露叶绿体细长,基质片层和基粒片层都受到严重破坏(图3A、C),说明20 d时20%自然光照影响到叶绿体的发育。20%弱光条件下公子基粒片层加厚,这与前人在马铃薯的研究结果相似[9],因为类囊体垛叠高是植物最大限度利用光能的一种适应方式;类囊体垛叠使LHCII间连接更紧密,可增加捕光能力和能量传递效率[21]。淀粉体是贮藏光合产物淀粉的一个重要场所,20%弱光条件下,清露淀粉粒数量显著下降,下降幅度比公子大。基于以上论述,说明在20%光照条件下,公子比清露的适应能力更强。
在高等植物的发育过程中,光不但直接参与光合作用,而且还作为一种重要的信号物质调控相关基因的转录[22]。叶绿素a/b结合蛋白(LHC)是核基因编码的类囊体蛋白,是由定位于核内的cab基因编码,Lhcb2是编码光系统Ⅱ蛋白的其中一个基因,低等生物绿藻在50μmol/m2s光强下表达量比在200μmol/m2s光强下都高[23],与以上结论一致,弱光处理使菊花Lhcb2转录水平提高。低光照会引起茎徒长,乙烯参与到茎伸长的调控中[24],对向日葵的研究发现,处理7 d时,10μmol/m2s光照条件下的乙烯水平极显著高于1 000μmol/m2s光照条件下的乙烯水平[25]。与以上结论相同,本研究发现,弱光条件下乙烯水平与对照相比显著增加(图5C、D),清露CmACO2在第1天出现个峰值,公子CmACO2在1~4 d逐渐增加,说明清露在基因调控方面比公子更敏感。

图5 CmLhcb2和CmACO2在不同光照条件下的表达分析Figure 5 Expression analysis of CmLhcb2 and CmACO2 under different conditions
由此可见,在基因水平上,2种菊花叶片的CmLhcb2和CmACO2在弱光条件下都显著增加,清露受到的影响更显著,而且比公子反应更早;生长状况和组织结构方面,清露随着弱光处理时间的延长,栅栏组织和类囊体结构组织细胞出现畸形,公子在弱光条件下作了适应性调整来抵御弱光胁迫。
