粒用高粱F2群体农艺性状数量遗传分析
2019-03-08白晓倩于澎湃李延玲高建明裴忠有孙守钧
白晓倩,于澎湃,李延玲,高建明,裴忠有,罗 峰,孙守钧
(天津农学院 农学与资源环境学院,天津 300384)
高粱(Sorghumbicolor(L.) Moench)为禾本科高粱属作物,是世界上种植的第五大禾谷类作物,也是我国主要旱粮作物之一[1-2],具有抗旱、耐涝、耐贫瘠、耐盐碱等优良特性。在我国,高粱有着悠久的种植历史,自古以来便在我国粮食作物栽培中占据重要地位,且为解决我国粮食生产和饥荒问题做出了重大贡献[3-4]。高粱从用途上可以分为粒用高粱、饲用高粱、甜高粱及帚用高粱四大类;其中,粒用高粱主要用于酿酒、食用、饲料等方面,是重要的酿造原料和粮饲作物[5-8]。一直以来,培育高产高粱品种被视为首要攻坚任务,而大多数产量性状为数量遗传性状,受多个表型性状的综合影响,且各性状之间又相互关联、彼此制约[9-10]。因此,要想培育高产的高粱品种,必须对与高粱产量相关的农艺性状进行分析。
农艺性状是影响高粱育种的重要因素之一,目前,国内大多数粒用高粱品种植株偏高,为机械化收获增加难度;穗柄较短、旗叶鞘较长,会出现“鞘包穗”现象,这种现象致使未抽出的穗发生败育,严重影响产量,有时也会因“包穗”现象,导致旗叶鞘内长期积水,致使穗柄木质化程度降低,容易发生病虫害,穗柄极易折断,然而旗叶鞘过短,又容易出现穗弯曲,严重则会引起倒伏,原因是旗叶鞘包被可增强穗柄的支撑力度[11-12],此外,穗柄长还与单株产量存在负相关关系[13];穗较长,导致开花期延长,相同时间内籽粒成熟度不一致;主茎茎节数越少、平均茎节长越长,其植株的节间数越少,植株在收获后可以作为反刍动物的饲用原料,使其秸秆在收获后可以得到充分利用。农艺性状的选择比较直观且方便测量,同时又与其他性状相关联,对其遗传规律进行深入研究,可以为育种工作者提供可靠的遗传信息[14-16]。
本试验选取农艺性状差异较大的国内粒用高粱品种忻粱52和美国引进的粒用高粱品种美引-20为亲本进行杂交,得到F1,经过F1自交获得F2分离群体,对F2群体单株的株高、穗长、穗柄长、旗叶鞘长、主茎茎节数及平均茎节长6个农艺性状进行数量遗传分析,旨在通过这6个农艺性状的主-多基因分析深入研究其遗传规律并确定其广义遗传率,为田间选择优良性状提供参考。
1 材料和方法
1.1 试验材料
试验通过对多个高粱品种农艺性状的测定,选取性状差异大的国内粒用高粱品种忻粱52为父本、美国引进粒用高粱品种美引-20为母本,进行杂交得到F1,经F1自交得到375株F2分离群体。
1.2 试验设计
试验于2016年8月在天津市静海县良种场进行亲本杂交,配制正反交组合,同年在海南试验田进行加代繁殖,2017年初获得F2群体种子。2017年5月在静海良种场按顺序播种亲本、F1和F2,其中,亲本各种植2行,F1正反交各种植3行,F2分离群体种植30行。本试验材料田间种植均行长5 m,行间距0.5 m,株距0.2 m,田间管理同大田生产。
1.3 测定方法
田间性状测定:亲本、F1、F2于完熟期在田间进行农艺性状的测量(为避免边际效应,除去小区两边的两行和每行两端的植株),F2群体需对单株进行各性状的测定[17]。
株高(cm):在完熟期,用钢卷尺测定单株植株从地面至穗顶端的距离;穗长(cm):在完熟期,用钢卷尺测定单株植株从穗顶端到穗基部的距离;穗柄长(cm):在完熟期,剥去旗叶叶鞘,用钢卷尺测定穗基部到第一节间的距离;旗叶鞘长(cm):在完熟期,用钢卷尺测定旗叶叶鞘到第一节间的距离;主茎茎节数(节):在完熟期,剥去整个植株的叶片及叶鞘,数出主茎茎节数;平均茎节长(cm):在完熟期,测定的株高和穗长的差与茎节数的比值。
1.4 数据分析
基本数据采用Microsoft Excel 2010和SPSS Statistics 17.0进行处理,F2世代分析根据王建康、盖钧镒[18-21]提出的单个分离世代群体的遗传模型方法-主-多基因遗传分析模型进一步分析,得到本试验F2群体的6个农艺性状的遗传模型以及与之相应的AIC值,并计算出遗传率。
2 结果与分析
2.1 高粱F2群体各农艺性状相关性分析
忻粱52×美引-20杂交F2代群体各农艺性状的相关性分析见表1。由表1可知,株高与穗长、穗柄长、主茎茎节数、平均茎节长呈极显著正相关,其中相关程度最高的性状为平均茎节长,相关系数为0.789,其次是主茎茎节数和穗长,相关系数分别为0.511和0.278;穗长与穗柄长、旗叶鞘长呈极显著正相关,相关系数分别为0.203和0.356,与平均茎节长呈显著正相关,相关系数为0.110;穗柄长与旗叶叶鞘长、平均茎节长呈极显著正相关,相关系数分别为0.711和0.368,与主茎茎节数呈极显著负相关,相关系数为-0.331;旗叶鞘长与平均茎节长呈极显著正相关,相关系数为0.208,与主茎茎节数呈极显著负相关,相关系数为-0.283;而主茎茎节数与平均茎节长无显著相关。

表1 F2群体表型性状相关性分析Tab.1 Correlation analysis of phenotypic traits in F2 population
注:**.在0.01水平(双侧)上显著相关;*.在0.05水平(双侧)上显著相关。
Note:**.Significant correlation at 0.01 level(both sides);*.Significant correlation at 0.05 level(both sides).
2.2 高粱F2群体农艺性状表型分析
为了了解粒用高粱农艺性状的遗传规律,在忻粱52×美引-20杂交F2群体各农艺性状统计分析(表2)和各农艺性状频数分布图中可以看出(图1),株高、穗长、穗柄长、旗叶鞘长、主茎茎节数、平均茎节长这6个农艺性状的频数分布连续且呈正态分布,符合数量性状遗传特征,说明这6个性状属于数量遗传性状。

表2 F2群体农艺性状统计分析Tab.2 Statistical analysis of agronomic characters in F2 population
2.3 遗传模型的选择
根据盖钧镒-主-多基因混合遗传模型F2单世代分析方法,得到株高、穗长、穗柄长、旗叶鞘长、主茎茎节数和平均茎节长这6个农艺性状的极大似然值和AIC值(表3-4)。根据AIC值最小原则,每个性状挑选出4个AIC值最小的模型作为备选模型(表中用“*”标出)。
2.4 遗传模型检测
利用U12、U22、U32(均匀性检验)、nW2(Smirnov检验)、Dn(Kolmogorov检验)对这6个农艺性状的4个备选模型的F2世代进行适合性检验(表5),选出6个性状最适的遗传模型。
在株高的4种遗传备选模型中, Kolmogorov检验均达到显著水平(P<0.05),其中只有Model B_1 的Smirnov检验达到显著水平(P<0.05),而其他统计量均未达到显著水平,且Model B_1的AIC值最小;因此,选择Model B_1作为该群体株高性状的最适遗传模型,说明在该群体中株高受2对主基因控制,为加性-显性-上位性的混合遗传模型。
在穗长的4种遗传备选模型中, Kolmogorov检验均达到显著水平(P<0.05),而Smirnov检验均未达到显著水平,但相较于另外3种备选模型来说,Model A_0的AIC值最小;因此,选择Model A_0模型作为该群体穗长性状的最适遗传模型,说明穗长没有主基因控制,而是微效多基因遗传的性状。
在穗柄长的4种遗传备选模型中,Kolmogorov检验均达到显著水平(P<0.05),其中只有Model A_1在Smirnov检验中达到显著水平(P<0.05),而且Model A_1在4种备选模型中的AIC值最小;因此选择Model A_1作为该群体穗柄长的最适遗传模型,说明穗柄长是受1对主基因控制,为加性-显性混合遗传模型。

图1 F2群体农艺性状频数分布Fig.1 Frequency distribution of agronomic traits in F2 population

表3 高粱株高、穗长、穗柄长遗传模型的极大似然值和AIC值Tab.3 Maximum likelihood value and AIC values of genetic model of plant height, panicle length and ear length of sorghum
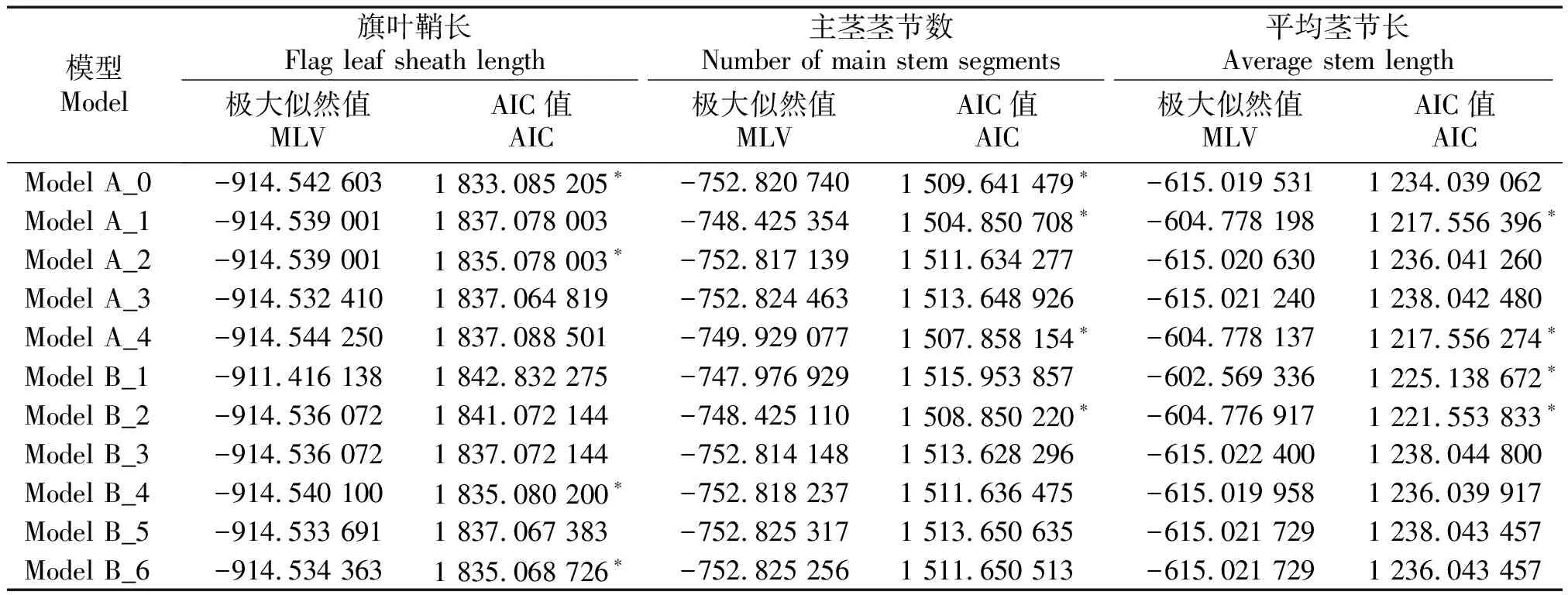
表4 高粱旗叶鞘长、主茎茎节数、平均茎节长遗传模型的极大似然值和AIC值Tab.4 Maximum value likelihood value and AIC values of genetic model flag leaf sheath length,the number of main stem segments and average stem length of sorghum
在旗叶鞘长的4种备选模型中,Smirnov检验和Kolmogorov检验均达到显著水平(P<0.05),均匀性检验均未达到显著水平,其中以Model A_0的AIC值最小。因此,选择Model A_0作为该群体旗叶鞘长的最适遗传模型,说明旗叶鞘长是仅受微效多基因控制而不受主基因控制的性状。
在主茎茎节数的4种遗传备选模型中,均匀性检验、Smirnov检验和Kolmogorov检验均未达到显著水平,其中,Model A_1相对于另外3种备选模型来说AIC 值最小;因此,选择Model A_1为该群体主茎茎节数的最适遗传模型,说明主茎茎节数受1对主基因控制,为加性-显性混合遗传模型。
在平均茎节长的4种遗传备选模型中,除均匀性检验未达到显著水平外,Smirnov检验和Kolmogorov检验均达到了显著水平(P<0.05),且Model A_4在4种模型中AIC值最小。因此,选择Model A_4作为该群体平均茎节长的最适遗传模型,说明该群体的平均茎节长受1对主基因控制,为负向完全显性的遗传模型。

表5 高粱各农艺性状备选模型的适合性检验Tab.5 Sorghum suitability test for alternative models of agronomic traits
2.5 遗传参数估计
根据已经确定的最优模型和IECM[22-24]的估算方法,对F2群体农艺性状进行遗传参数估计(表6)。由于穗长和旗叶鞘长都是微效多基因控制的数量性状遗传,故无法计算二者的主基因遗传率。
该群体的株高遗传符合Model B_1模型,F2群体的分离符合加性-显性-上位性的混合遗传模型;其中,株高的平均值m为120.289 0,偏向于植株相对较高的亲本忻粱52,控制株高的2对主基因的加性效应之和为29.662 3,显性效应之和为-6.275 4,加性效应显著大于显性效应,说明控制该群体株高性状的2对主基因以加性效应为主,主基因遗传率为88.65%。
穗柄长的遗传模型符合Model A_1,F2分离群体符合加性-显性混合遗传模型,受1对主基因控制;穗柄长的平均值35.968 200,偏向于穗柄长相对较长的亲本美引-20,控制穗柄长的主基因的加性效应为4.643 999,显性效应为-0.154 951,说明控制穗柄长的基因以加性效应为主,主基因遗传率为61.58%。
主茎茎节数的遗传模型符合Model A_1,F2分离群体的主茎茎节数受1对主基因控制,符合加性-显性混合遗传模型;控制主茎茎节数的主基因的加性效应为2.033 050,显性效应为-0.571 172,加性效应显著大于显性效应,说明控制该群体主茎茎节数的基因以加性效应为主,主基因遗传率为68.94%。
平均茎节长的数量遗传模型符合Model A_4,F2分离群体的平均茎节长符合1对主基因遗传的负向完全显性遗传模型,主基因遗传率为76.99%;控制该群体平均茎节长的主基因加性效应为1.012 063 5,说明控制该群体平均茎节长的主基因以加性效应为主,主基因遗传率为49.24%。

表6 高粱各农艺性状遗传参数估计Tab.6 Estimation of genetic parameters of agronomic traits in sorghum


3 结论与讨论
本试验表明,株高符合Model B_1,受2对主基因控制的加性-显性-上位性的混合遗传模型。该性状的加性效应显著大于显性效应,以加性效应为主,主基因遗传率为88.65%,后代遗传也比较稳定,不易受环境影响。而蒋峰等[25]的试验表明,株高符合Model B_2,结果并不一致,造成这种现象的原因可能是所进行试验的亲本及群体大小不同。
穗长与穗柄长、旗叶鞘长存在一定相关性;同时,穗柄长与旗叶鞘长具有极显著的正相关关系,即穗柄长越长,旗叶鞘长也越长。通过主-多基因混合遗传模型分析可知,穗长和旗叶鞘长的遗传均符合A_0模型,两者都不受主基因的控制,而是受微效多基因的控制。穗柄长的遗传符合A_1模型,即加性-显性的遗传模型,受1对主基因的控制,以加性效应为主,主基因遗传率为61.58%,说明不易受到环境的影响,可以在后代中稳定遗传。因此,在进行株型选择时,不仅要考虑基因对该性状的作用,还要尽可能地去消除环境因素造成的影响[26]。高士杰等[27]对穗长的试验研究表明,加性效应占70.43%,显性效应占20.51%,上位效应占9.07%,因此,控制该性状的遗传以加性效应为主。孙守钧等[11]指出,旗叶鞘长以加性效应为主,占92.8%;穗柄长除加性效应外另有35.7%非加性效应,二者都易受环境影响。显然,与本课题试验的研究结果不一致。试验材料所选亲本的不同、人为因素的存在以及外界环境的影响都是造成试验结果存在差异的重要因素[28]。
主茎茎节数符合Model A_1,受1对主基因控制,属于加性-显性混合遗传模型。且该性状的加性效应显著大于显性效应,以加性效应为主,其遗传率为68.94%,遗传力较高,说明该农艺性状不易收到环境的影响。平均茎节长符合Model A_4,受1对主基因控制的负向完全显性模型,它的遗传率为49.24%,遗传率较低,在后代遗传中不稳定,容易受到环境的影响。
综上所述,本试验所用高粱群体的株高、穗柄长和主茎茎节数的遗传率较高,受环境因素影响较小,在后代中遗传比较稳定,可以在育种早代直接进行选育;平均茎节长的遗传率较低,说明该性状在后代遗传中受环境影响较大,遗传不稳定,需在高代中进行选育。在高粱育种中,数量性状不仅受遗传因子控制,同时在很大程度上也受环境因素的支配。数量性状遗传上除具有“一因多效”和“多因共效”特性外,同时,外界环境也会对其产生一定程度的影响[29]。因此,在高粱育种中,尤其应注意环境因素对数量性状产生的影响。
