杉木纯林和混交林林下草本层种群生态位特征
2019-02-28冯玉超郑晓阳王正宁兰思仁
冯玉超,郑晓阳,王正宁,刘 博,兰思仁
(福建农林大学林学院/ 国家林业局杉木工程技术研究中心,福建 福州 350002)
随着人工林经营管理从过去强调木材生产到现在注重生态功能发挥的转变,如何在保证人工林经济价值的同时发挥其生态功能,实现可持续经营成为目前林业经营管理中亟需解决的问题。目前,人工林林下植物物种多样性已成为森林经营管理的一个主要目标,探究人工林林下植物物种组成关系及其对环境的适应性是评估人工林生态系统的必要途径。作为森林生态系统的重要组成部分,林下植被对维持森林生物多样性、维护立地生产力和生态系统稳定性等方面具有不可忽视的作用[1],尤其是草本层因其所需的生态位相对较小,生长、发育和丰富程度容易受外界环境的影响,对生态系统环境变化最为敏感,因此对森林生态系统健康和立地指数具有重要的指示作用[1]。评估不同林分类型人工林林下草本层植物的生长发育特征、种间关系及种群在群落中功能地位,对促进人工林物种多样性和生态功能恢复,实现可持续经营管理具有重要的现实意义。
生态位研究是解决人工林林下植物共存和相互适应的主要手段,对森林生态系统的恢复与重建具有重要的理论和现实意义[2]。目前国内外研究主要集中于不同植物群落和生境条件下森林植物种群生态位研究,而基于不同林分类型杉木人工林林下草本层生态位相关研究仍较为缺乏。
杉木(Cunninghamialanceolata)是我国南方林区主要的用材树种,栽培历史久远[3],作为杉木人工林生态系统的重要组成部分,林下植物在物种多样性研究领域至关重要[4],但由于杉木人工林的集约经营影响林下植物的生长发育,造成地力衰退、物种多样性下降等诸多生态问题,直接威胁杉木人工林的可持续经营,由此林下植物在人工林生态系统中的重要地位和作用重新引起重视。有关不同林分类型对人工林林下植被的研究多限于生物多样性和生物量的变化[5]。探究杉木人工林林下植被各种群之间相互关系及在群落中的功能地位,对于正确理解杉木林林下植被物种多样性具有重要的参考价值。该研究运用生态位理论,对杉木纯林和混交林林下自然更新的草本层种群的3种生态位指标进行比较和定量研究,旨在探讨不同林分类型下群落内优势种群对环境的生理适应性和种间竞争水平,揭示不同林分类型草本层对资源的利用能力,探究其在群落中的功能地位及其内在规律性,进一步探讨何种林分类型更有利于杉木人工林林下植被的生存发展,以期为森林可持续经营措施的选择和生产实践提供科学依据。
1 材料与方法
1.1 研究区自然概况
研究区位于福建省三明市莘口教学林场(26°07′~26°10′ N,117°24′~117°27′ E),地势总体呈西南高,东北低的走向,属中低山、丘陵为主的地貌类型,海拔高度250~500 m。土壤以暗红壤为主,成土母质以粉砂页岩为主。气候属于中亚热带季风气候,年平均降水量1 400~2 000 mm,年平均气温17.0~19.4 ℃,平均日照时数1 727.1~1 897.5 h,无霜期216~304 d,气候温暖,光照充足,水分充沛,适合杉木人工林的生长。该林场森林群落类型多样,植物种类资源丰富,是典型的杉木人工林研究基地。主要乔木树种有杉木、格氏栲(Castanopsiskawakamii)、木荷(Schimasuperba)、米槠(Castanopsiscarlesii)、马尾松(Pinusmassoniana)等,主要灌草植物有毛鳞省藤(Calamusthysanolepis)、杜茎山(Maesajaponica)、华山姜(Alpiniachinensis)、狗脊(Woodwardiajaponica)、江南短肠蕨(Allantodiametteniana)等。
1.2 研究方法
1.2.1样方设置
于2017年6月在研究区分别选取2块林龄相同且海拔、坡度、坡向等基本一致的杉木纯林和混交林。杉木纯林和混交林均为1957年格氏栲天然林皆伐后营造。在营林初期,杉木人工纯林和混交林(杉木-木荷混交,针阔混交比例6:4;初植密度均为3 000株·hm-2。2017年调查杉木纯林林分密度为1 427株·hm-2;混交林林分密度为1 375株·hm-2,郁闭度分别为0.63和0.57。)每个林分按照上、中、下不同坡位各设置3块20 m×20 m的样方,记录各样方的海拔、坡位、坡向、坡度和郁闭度,调查记录各样方乔木树种的物种名,并进行每木检尺(起测径阶为5 cm),测定每一种类的株数以及该种每个个体的胸径、树高、冠幅和枝下高。在每个乔木样方分别设置4个5 m×5 m灌木样方和在样方4个顶角及中心选取5个1 m×1 m草本样方。调查记录样方内物种的种类、株数、高度、盖度和基径等数据。林分基本情况见表1。
表1林分基本情况
Table1Basicconditionsofforests
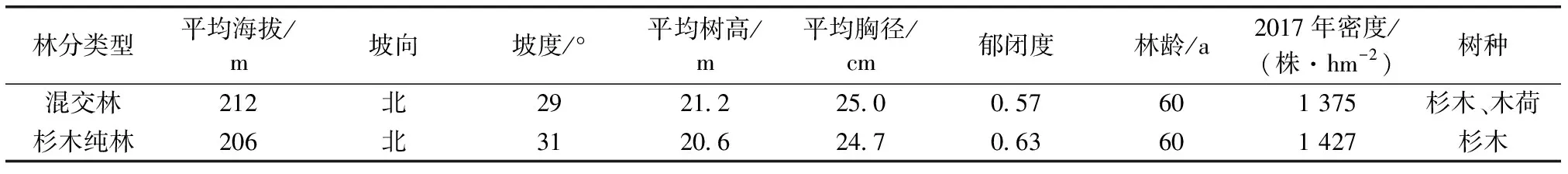
林分类型平均海拔/m坡向坡度/°平均树高/m平均胸径/cm郁闭度林龄/a2017年密度/(株·hm-2)树种混交林 212北2921.225.00.57601 375杉木、木荷杉木纯林206北3120.624.70.63601 427杉木
1.2.2计算方法
(1)重要值
重要值(importance value,V)是衡量物种在群落中的作用和地位的综合数量指标[6]。
V=(RC+RH+RD)/3。
(1)
式(1)中,RC为相对盖度;RH为相对高度;RD为相对密度。重要值的大小反映物种在群落中的优势程度,因此依据重要值大小筛选出不同林分类型下林下植物优势种群进行分析比较。
(2)生态位宽度
生态位宽度采用Shannon-Wiener生态宽度指数[7]和Levins生态宽度指数[8],计算公式分别为
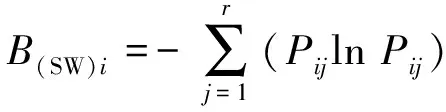
(2)
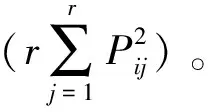
(3)
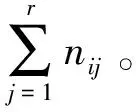
(4)
式(2)~(4)中,B(SW)i和B(L)i分别为物种的Shannon-Wiener生态位宽度指数和Levins生态宽度指数,值域分别为[0,lnr]和[1/r,1];Pij为物种i在第j资源位上的重要值与它在全部资源位上重要值的占比,nij为物种i在第j资源的重要值;r为资源位总位数(该文代表样方数)。
以上2个指数Bi值越大,说明生态位越宽。当一个种的个体以相等的数目利用每一资源状态时,Bi最大化,即该种具有最宽的生态位;当种i的所有个体都集中在某一个资源状态下时,Bi最小,该种具有最窄的生态位。
(3)生态位重叠
生态位重叠采用Levins生态位重叠指数[9],计算公式为
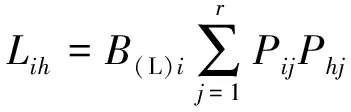
(5)
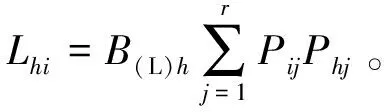
(6)
式(5)~(6)中,Lih和Lhi分别为物种i重叠物种h、物种i被物种h重叠的重叠指数,值域皆为[0,1];Phj为物种h在第j个资源位上的重要值与它在全部资源位上重要值的占比;B(L)h为物种h的Levins生态位宽度指数。
(4)生态位相似性
生态位相似性是指物种间对资源利用的相似程度,计算公式为
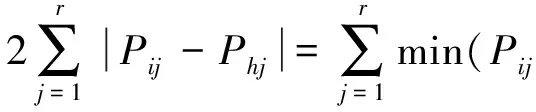
(7)
式(7)中,Cih为物种i与物种h之间的生态位相似程度,且Cih=Chi,值域为[0,1][10]。
1.3 数据处理
采用SPSS 19.0软件进行数据统计分析,在Excel 2010软件中计算生态位的各项指数,并构建生态位重叠指数和生态位相似性比例矩阵。
2 结果与分析
2.1 林下草本层优势物种的生态位宽度
样方调查表明,杉木的混交林和纯林林下植被种类分别有25种和22种,混交林种类比杉木纯林增加13.64%。由表2可知,无论是混交林还是纯林林下草本层优势种群的Shannon-Wiener和Levins生态位宽度指数的排序基本一致。2种经营方式下的林下共同优势植物有华山姜、狗脊、江南短肠蕨、杜茎山、扇叶铁线蕨(Adiantumflabellulatum)、团叶鳞始蕨(Lindsaeaorbiculata),其中华山姜和狗脊在混交林和纯林经营方式下的B(SW)和B(L)生态位宽度指数均最大;除了狗脊和江南短肠蕨外,同一物种在混交林下的B(SW)和B(L)生态位宽度指数均比纯林高。B(SW)生态宽度指数大于1的优势植物在混交林和纯林下分别占83.3%和58.3%;B(L)生态位宽度指数大于0.2的分别占66.6%和50.0%。从总体上看,混交林的林下草本层生态位宽度普遍比纯林高,资源利用率更大,相同物种在混交林中发展更具优势。
表2不同林分类型林下草本层优势物种生态位宽度
Table2Nichewidthofdominantspeciesinherbaceouslayerunderdifferentforesttypes
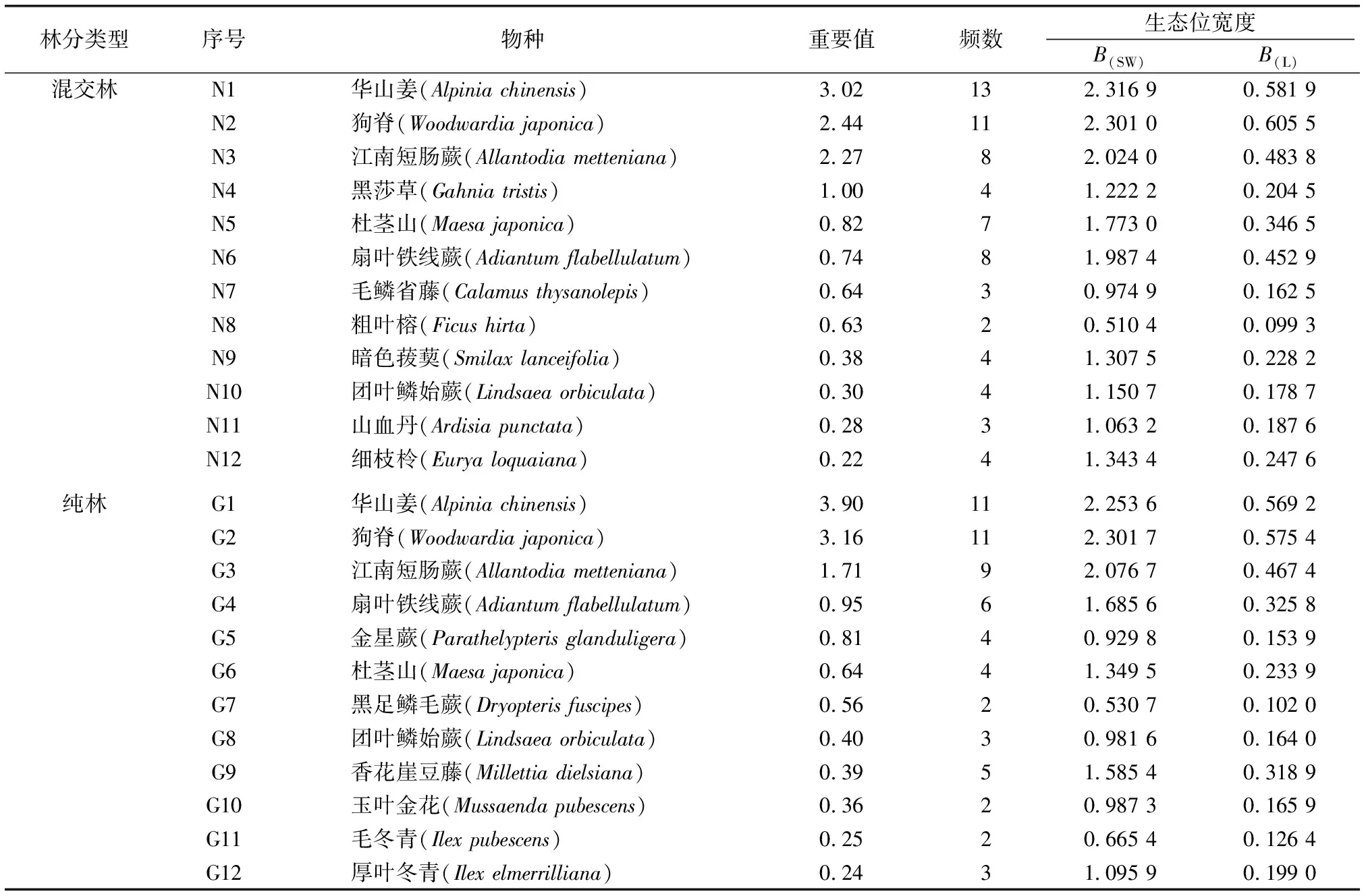
林分类型序号物种重要值频数生态位宽度B(SW)B(L)混交林 N1华山姜(Alpinia chinensis)3.02132.316 90.581 9N2狗脊(Woodwardia japonica)2.44112.301 00.605 5N3江南短肠蕨(Allantodia metteniana)2.2782.024 00.483 8N4黑莎草(Gahnia tristis)1.0041.222 20.204 5N5杜茎山(Maesa japonica)0.8271.773 00.346 5N6扇叶铁线蕨(Adiantum flabellulatum)0.7481.987 40.452 9N7毛鳞省藤(Calamus thysanolepis)0.6430.974 90.162 5N8粗叶榕(Ficus hirta)0.6320.510 40.099 3N9暗色菝葜(Smilax lanceifolia)0.3841.307 50.228 2N10团叶鳞始蕨(Lindsaea orbiculata)0.3041.150 70.178 7N11山血丹(Ardisia punctata)0.2831.063 20.187 6N12细枝柃(Eurya loquaiana)0.2241.343 40.247 6纯林G1华山姜(Alpinia chinensis)3.90112.253 60.569 2G2狗脊(Woodwardia japonica)3.16112.301 70.575 4G3江南短肠蕨(Allantodia metteniana)1.7192.076 70.467 4G4扇叶铁线蕨(Adiantum flabellulatum)0.9561.685 60.325 8G5金星蕨(Parathelypteris glanduligera)0.8140.929 80.153 9G6杜茎山(Maesa japonica)0.6441.349 50.233 9G7黑足鳞毛蕨(Dryopteris fuscipes)0.5620.530 70.102 0G8团叶鳞始蕨(Lindsaea orbiculata)0.4030.981 60.164 0G9香花崖豆藤(Millettia dielsiana)0.3951.585 40.318 9G10玉叶金花(Mussaenda pubescens)0.3620.987 30.165 9G11毛冬青(Ilex pubescens)0.2520.665 40.126 4G12厚叶冬青(Ilex elmerrilliana)0.2431.095 90.199 0
BSW为Shannon-Wiener生态位宽度指数,B(L)为Levins生态宽度指数。
2.2 林下草本层优势物种的生态位重叠
混交林和杉木纯林林下植被优势物种的生态位重叠矩阵见表3和表4。
表3混交林林下草本层优势物种的生态位重叠指数
Table3Nicheoverlapsofdominantspeciesinherbaceouslayerundermixedforest
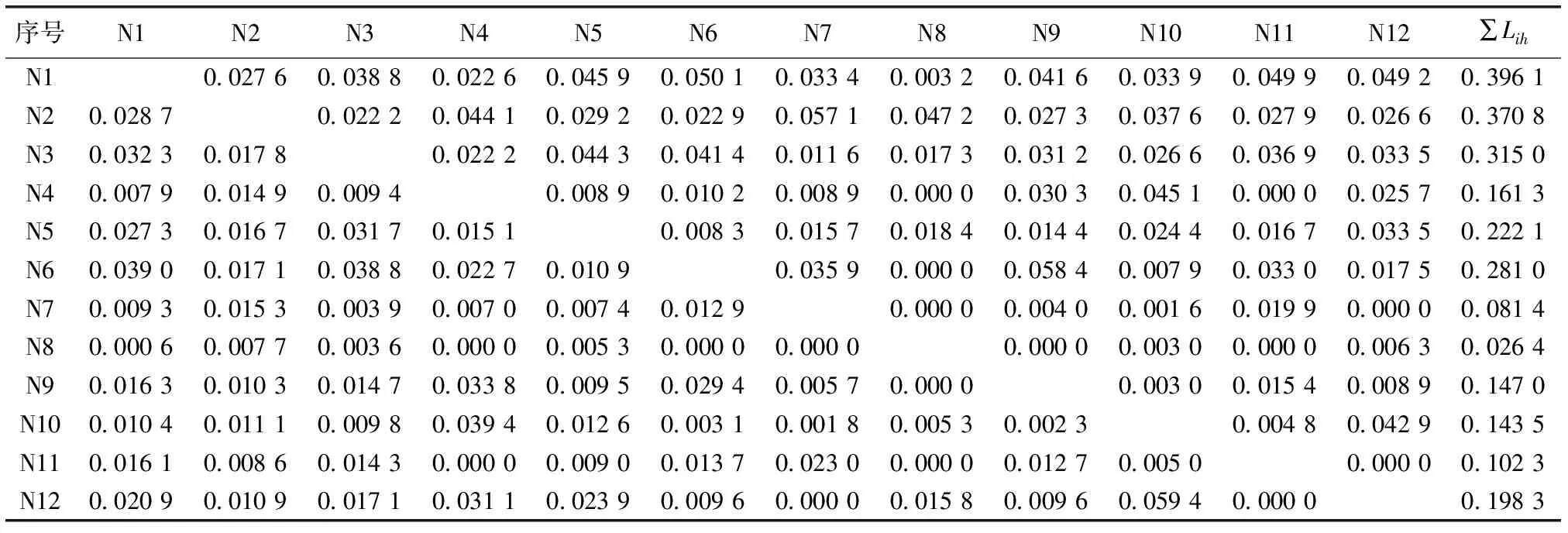
序号N1N2N3N4N5N6N7N8N9N10N11N12∑LihN10.027 60.038 80.022 60.045 90.050 10.033 40.003 20.041 60.033 90.049 90.049 20.396 1N20.028 70.022 20.044 10.029 20.022 90.057 10.047 20.027 30.037 60.027 90.026 60.370 8N30.032 30.017 80.022 20.044 30.041 40.011 60.017 30.031 20.026 60.036 90.033 50.315 0N40.007 90.014 90.009 40.008 90.010 20.008 90.000 00.030 30.045 10.000 00.025 70.161 3N50.027 30.016 70.031 70.015 10.008 30.015 70.018 40.014 40.024 40.016 70.033 50.222 1N60.039 00.017 10.038 80.022 70.010 90.035 90.000 00.058 40.007 90.033 00.017 50.281 0N70.009 30.015 30.003 90.007 00.007 40.012 90.000 00.004 00.001 60.019 90.000 00.081 4N80.000 60.007 70.003 60.000 00.005 30.000 00.000 00.000 00.003 00.000 00.006 30.026 4N90.016 30.010 30.014 70.033 80.009 50.029 40.005 70.000 00.003 00.015 40.008 90.147 0N100.010 40.011 10.009 80.039 40.012 60.003 10.001 80.005 30.002 30.004 80.042 90.143 5N110.016 10.008 60.014 30.000 00.009 00.013 70.023 00.000 00.012 70.005 00.000 00.102 3N120.020 90.010 90.017 10.031 10.023 90.009 60.000 00.015 80.009 60.059 40.000 00.198 3
表3中序号所对应物种见表2。∑Lih为该物种占用其他物种资源量的生态位重叠之和。
生态位宽度较大的华山姜、狗脊与其他物种间具有相对较高的生态位重叠,如混交林的华山姜-扇叶铁线蕨(N1-N6,0.050 1)、华山姜-山血丹(Ardisiapunctata)(N1-N11,0.049 9)、狗脊-毛鳞省藤(Calamusthysanolepis)(N2-N7,0.057 1)、狗脊-黑莎草(Gahniatristis)(N2-N4,0.044 1)和杉木纯林的华山姜-香花崖豆藤(Millettiadielsiana)(G1-G9,0.041 7)、华山姜-杜茎山(Maesajaponica)(G1-G6,0.041 5)、狗脊-黑足鳞毛蕨(Dryopterisfuscipes)(G2-G7,0.045 7)、狗脊-毛冬青(Ilexpubescens)(G2-G11,0.078 3);生态位宽度较大的物种在这2种不同经营方式下均具有相对高的∑Lih值(该物种占用其他物种资源量的生态位重叠之和),如华山姜、狗脊和江南短肠蕨;少数生态位宽度较小的物种出现较大的生态位重叠,如混交林的团叶鳞始蕨-细枝柃(Euryaloquaiana)(N10-N12,0.042 9 )以及杉木纯林的厚叶冬青(Ilexelmerrilliana)-香花崖豆藤(Millettiadielsiana)(G12-G9,0.041 3),但团叶鳞始蕨和厚叶冬青占用其他物种资源量的生态位重叠之和仍较小。
表4纯林林下草本层优势物种的生态位重叠指数
Table4Nicheoverlapsofdominantspeciesinherbaceouslayerunderpureforest
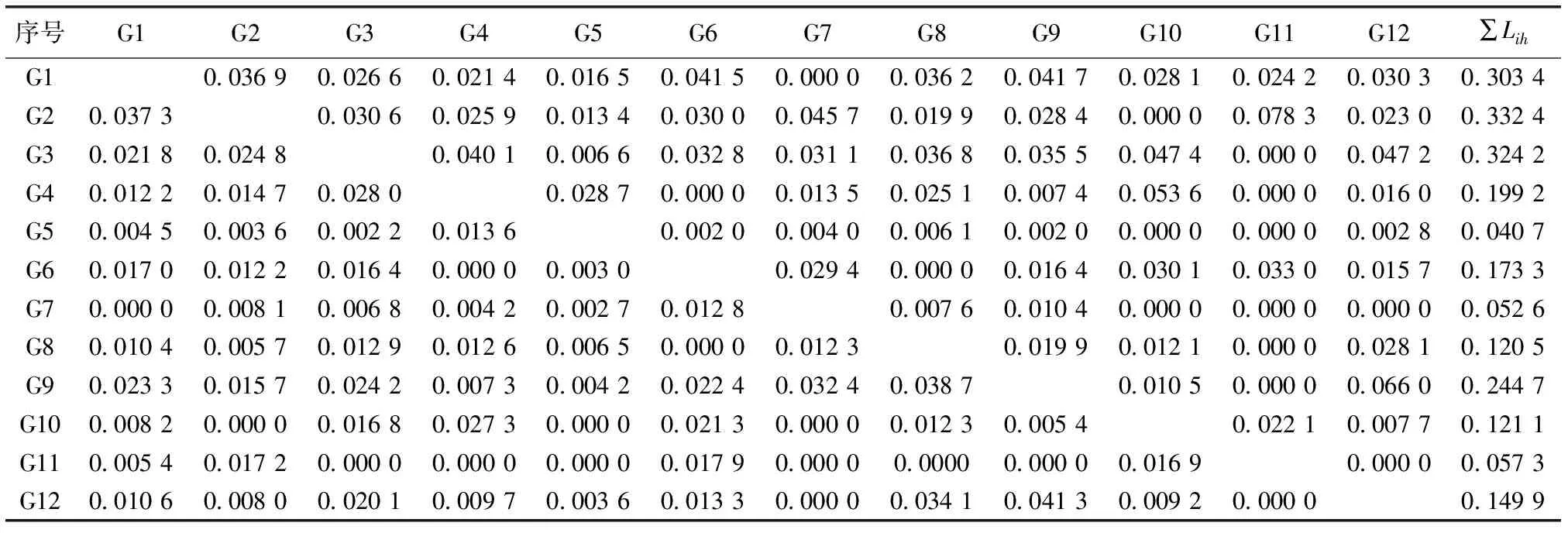
序号G1G2G3G4G5G6G7G8G9G10G11G12∑LihG10.036 90.026 60.021 40.016 50.041 50.000 00.036 20.041 70.028 10.024 20.030 30.303 4G20.037 30.030 60.025 90.013 40.030 00.045 70.019 90.028 40.000 00.078 30.023 00.332 4G30.021 80.024 80.040 10.006 60.032 80.031 10.036 80.035 50.047 40.000 00.047 20.324 2G40.012 20.014 70.028 00.028 70.000 00.013 50.025 10.007 40.053 60.000 00.016 00.199 2G50.004 50.003 60.002 20.013 60.002 00.004 00.006 10.002 00.000 00.000 00.002 80.040 7G60.017 00.012 20.016 40.000 00.003 00.029 40.000 00.016 40.030 10.033 00.015 70.173 3G70.000 00.008 10.006 80.004 20.002 70.012 80.007 60.010 40.000 00.000 00.000 00.052 6G80.010 40.005 70.012 90.012 60.006 50.000 00.012 30.019 90.012 10.000 00.028 10.120 5G90.023 30.015 70.024 20.007 30.004 20.022 40.032 40.038 70.010 50.000 00.066 00.244 7G100.008 20.000 00.016 80.027 30.000 00.021 30.000 00.012 30.005 40.022 10.007 70.121 1G110.005 40.017 20.000 00.000 00.000 00.017 90.000 00.00000.000 00.016 90.000 00.057 3G120.010 60.008 00.020 10.009 70.003 60.013 30.000 00.034 10.041 30.009 20.000 00.149 9
表4中序号所对应物种见表2。∑Lih为该物种占用其他物种资源量的生态位重叠之和。
由图1可以看出,混交林和杉木纯林下生态位重叠指数为0的物种分别占总数的12.1%和21.2%,说明杉木纯林林下物种不发生重叠的比重大于混交林;生态位重叠指数在(0,0.02]、(0.02,0.04]、(0.04,0.06]、(0.06,0.08]区间的分别占49.3%和43.9%、28.0%和27.3%、10.6%和6.1%、0%和1.5%;生态位重叠平均值分别为0.018 5 和0.016 1。从总体分布格局来看,无论是混交林还是杉木纯林的生态位重叠指数普遍不高,种间竞争强度和物种对环境资源的共享能力不强,但总体上混交林林下植被物种间生态位重叠指数大于杉木纯林。
2.3 林下草本层优势物种的生态位相似性
通过比较生态位相似性比例和生态位重叠矩阵发现(表5~6),生态位重叠不仅与生态位宽度相关联,还与生态位相似性比例有一定的相关性。分析结果显示,生态位相似性比例较大的物种其生态位重叠也较大,如生态位相似性比例较大的物种华山姜-扇叶铁线蕨(N1-N6,0.5373)、江南短肠蕨-杜茎山(N3-N5,0.524 9)、 华山姜-狗脊(G1-G2,0.551 3)、狗脊-江南短肠蕨(G2-G3,0.434 3)具有较大的生态位重叠。
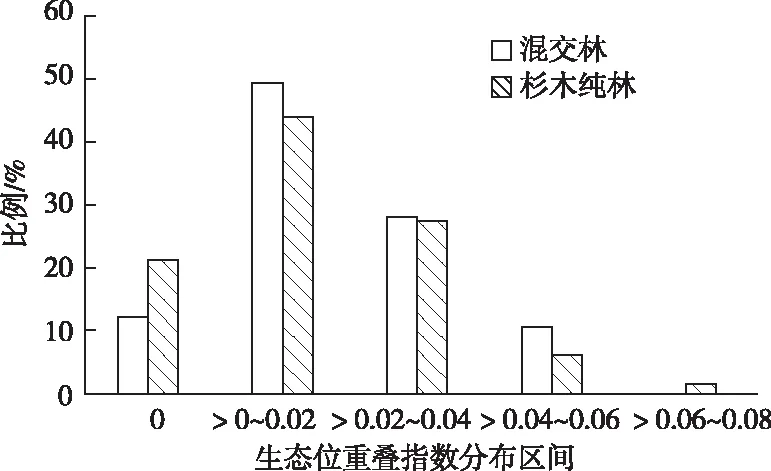
图1 不同林分类型生态位重叠指数分布格局
在林下植被12个主要优势物种的66个种对中,混交林和杉木纯林草本层生态位相似性比例为0的分别占总数的12.1%和15.2%;生态位相似性比例在(0,0.2]、(0.2,0.4]和(0.4,0.6]区间的分别占42.4%和39.4%、31.8%和40.9%、13.7%和3.0%;少数分布在(0.6,0.8]区间,分别占0%和1.5%(图2);混交林和杉木纯林草本层生态位相似性比例的总体平均值分别为0.2161和0.189 7,说明无论是混交林还是杉木纯林,物种间对环境资源的要求均存在较大的差异,但混交林总体的生态位相似性比例高于杉木纯林。
表5混交林林下草本层优势物种的生态位相似性比例
Table5Nicheproportionalsimilarityofdominantspeciesinherbaceouslayerundermixedforest
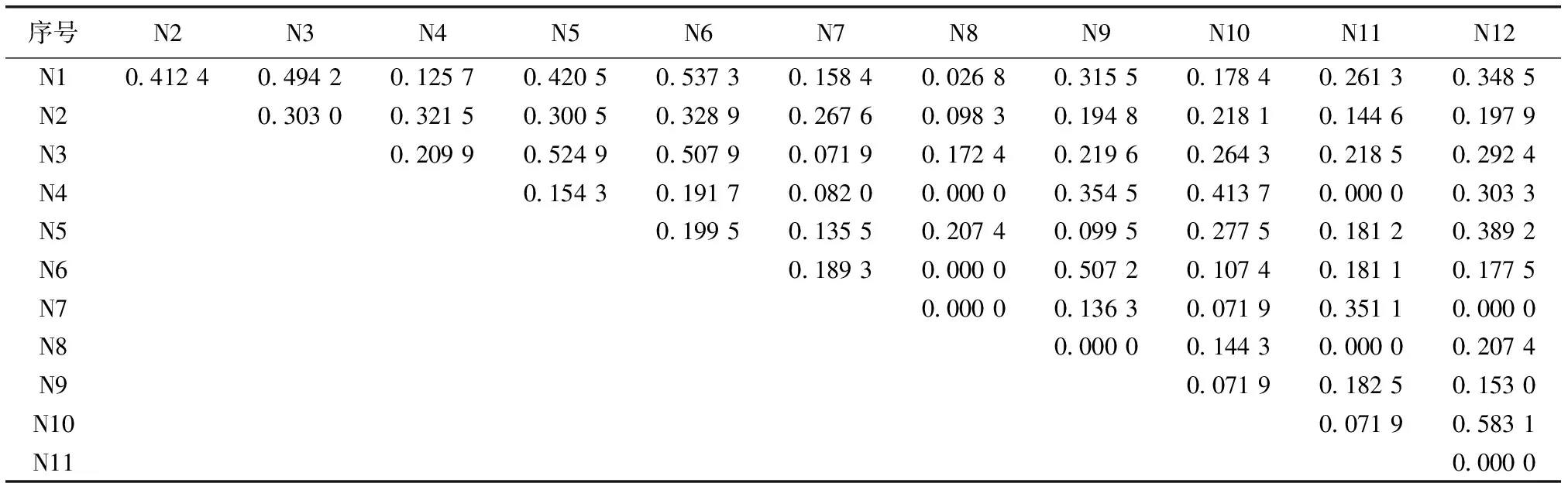
序号N2N3N4N5N6N7N8N9N10N11N12N10.412 40.494 20.125 70.420 50.537 30.158 40.026 80.315 50.178 40.261 30.348 5N20.303 00.321 50.300 50.328 90.267 60.098 30.194 80.218 10.144 60.197 9N30.209 90.524 90.507 90.071 90.172 40.219 60.264 30.218 50.292 4N40.154 30.191 70.082 00.000 00.354 50.413 70.000 00.303 3N50.199 50.135 50.207 40.099 50.277 50.181 20.389 2N60.189 30.000 00.507 20.107 40.181 10.177 5N70.000 00.136 30.071 90.351 10.000 0N80.000 00.144 30.000 00.207 4N90.071 90.182 50.153 0N100.071 90.583 1N110.000 0
表5中序号所对应物种见表2。
表6纯林林下草本层优势物种的生态位相似性比例
Table6Nicheproportionalsimilarityofdominantspeciesinherbaceouslayerunderpureforest
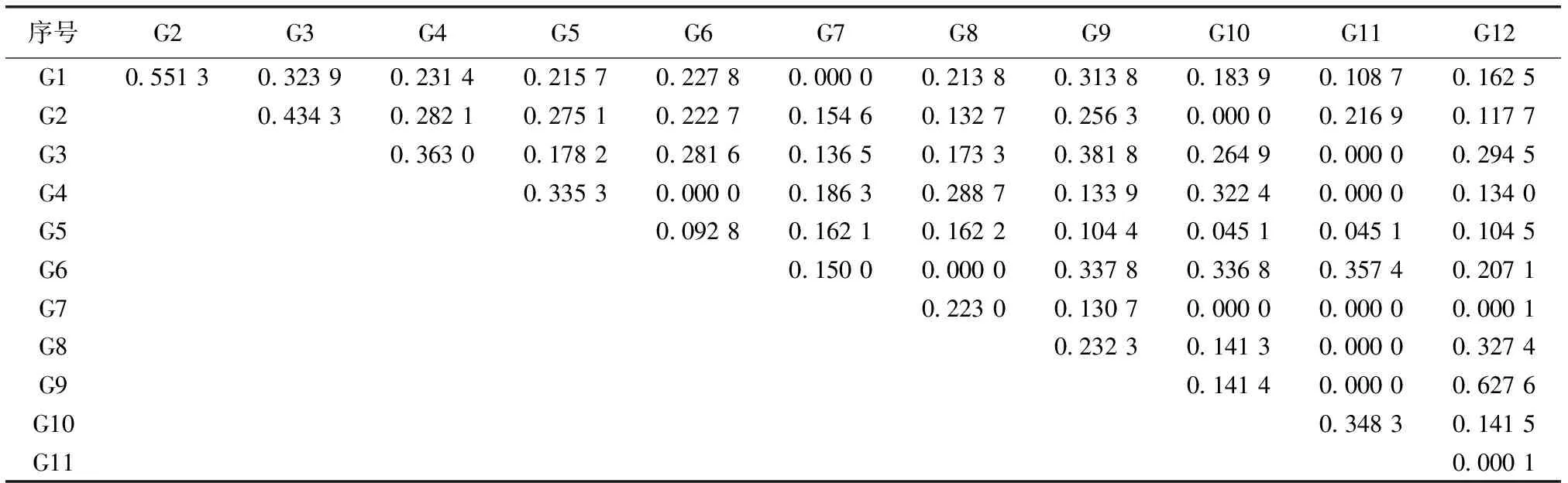
序号G2G3G4G5G6G7G8G9G10G11G12G10.551 30.323 90.231 40.215 70.227 80.000 00.213 80.313 80.183 90.108 70.162 5G20.434 30.282 10.275 10.222 70.154 60.132 70.256 30.000 00.216 90.117 7G30.363 00.178 20.281 60.136 50.173 30.381 80.264 90.000 00.294 5G40.335 30.000 00.186 30.288 70.133 90.322 40.000 00.134 0G50.092 80.162 10.162 20.104 40.045 10.045 10.104 5G60.150 00.000 00.337 80.336 80.357 40.207 1G70.223 00.130 70.000 00.000 00.000 1G80.232 30.141 30.000 00.327 4G90.141 40.000 00.627 6G100.348 30.141 5G110.000 1
表6中序号所对应物种见表2。
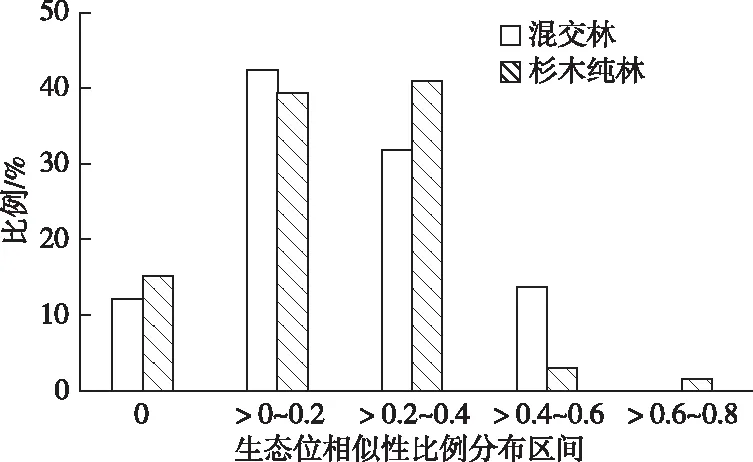
图2 不同林分类型生态位相似性比例分布格局
3 讨论
生态位宽度是一个物种对不同资源利用状况的衡量指标,其大小取决于物种对环境资源利用和适应能力。生态位宽度与环境的变化密切相关,物种对资源的利用及对环境适应性随环境的改变而发生变化[11]。研究结果发现,物种在群落中出现的频率越高,生态位宽度越大,其在群落中处于明显的优势地位,比生态位宽度窄的种群能更好地利用资源,具有极强的环境适应能力和竞争力,更倾向于泛化种,这与山西辽东栎群落优势种群生态位研究结果相似[12]。同时,生态位宽度的涵义在于体现物种利用资源的幅度[13],该研究中杉阔混交林林下草本层植物种类多于杉木纯林且生态位宽度普遍比杉木纯林高,说明植物对资源的利用程度更明显,种群在群落发展中更具优势,更趋于多物种均衡发展,资源空间不再仅为少数优势种群所占据,而是为多物种生长发育提供了更优的生长空间。种群在群落中具有更大资源利用幅度,对林下环境具有极强的生态适应性和竞争力,对群落的改造和构建起着重要作用。有研究发现,杉木混交林水稳性团聚体含量和枯落物归还量及枯落物分解速度高于杉木纯林,土壤肥力更高[14]。由于杉木纯林枯落物组成的单一性造成土壤微生物少,枯落物难以分解,而杉木混交林林分树种组成结构优于纯林,林地上保存了当地乡土阔叶树种,林地阔叶枯落物的增加为林地提供了更多的有机物,为改善土壤肥力提供了更佳的物质条件[15],丰富了林地植物的物种多样性,并为物种提供了更多可利用的资源空间。合理栽植混交林可以有效改善土壤肥力状况和杉木林地生态环境[16],而杉木纯林林分结构单一、林地养分较低等因素可能是导致生态位宽度总体比混交林窄的原因。
生态位重叠被视为种群对相同资源的共同利用或对生态空间区域的共同占有[17],有学者认为它可能是物种多样性和群落结构的决定因素之一[18]。物种对环境的适应能力取决于种群生态位宽度和生态位重叠度大小。如果某一种群生态位宽度越大,那么该种群与其他种群发生生态重叠的可能性越大,这与前人的研究结果一致[19]。但生态位宽度较小的物种也可能发生较大的生态位重叠,生态位宽度与生态位重叠并没有绝对的正相关关系,这与金佛山巴山榧树灌丛群落主要木本植物种群生态位特征的研究结果[20]相似。这可能是由于林区内人类干扰频繁无序,森林生态系统受到一定的影响,使得个别环境资源的空间异质性高,种群更偏向于集中分布在适宜生长的资源位,而在其他空间分布较少,导致某些种群在整个群落内的生态位宽度较窄。其次,资源量、资源的供求比、资源对生物需求的满足程度等因素共同决定着种群间生态位重叠程度,在资源丰富的情况下,种间竞争并不激烈,即使2种生态位宽度较大的物种共同生活在同一环境中,也不一定会出现高的生态位重叠。同时也发现,某物种在占用其他物种资源量的生态位重叠之和在排序上基本与生态位宽度一致。这是由于生态位宽度较大的种群一般分布范围较广、环境适应能力较强,与其他物种在群落中的生态位重叠之和较大。而生态位宽度较小的物种即便在某些资源位上拥有较大的生态位重叠,在整个群落中的生态位重叠总和依然较小。
物种间对资源利用能力随着生态位宽度的增大而提高,生态位重叠和相似性比例较高的物种之间可能存在着竞争关系[21]。不管在混交林还是杉木纯林中占重要值较大比重的华山姜和狗脊与其他物种间的生态位重叠指数和生态位相似比例均较大,说明华山姜、狗脊与其他物种间的竞争可能更加激烈。奚为民[22]在怀柔山区灌丛群落优势种群生态位的研究中认为,生态位重叠不仅是种间生态位相似性的重要参照指标,还与种间竞争水平密切相关。当资源丰富时,种间生态位重叠主要反映物种间因生态位相似性而占据相似的生态空间;当资源缺乏时,种间生态位重叠是表征物种间竞争关系和生态位相似性的共同指标。在该研究中,混交林和杉木纯林的生态位重叠指数普遍不高,主要集中在[0,0.02)区间,种间竞争强度和物种对环境资源的共享能力不强,种群存在分化现象,说明林地资源较为丰富。除部分物种存在激烈竞争关系外,大部分物种可能由于物种间生态位相似性而占据着相似的生态空间。但混交林的生态位重叠度和生态位相似性比例的平均值均比杉木纯林高,这是由于在一定的环境资源中,混交林群落中物种多样性高于纯林,物种间对资源的竞争程度较大,导致混交林物种间竞争能力大于杉木纯林。符饶等[21]在研究青衣江中游河滨草本植物生态位和物种多样性中也发现生境的物种多样性越丰富,则其平均生态位宽度和重叠越大。
种群生态位的这3个重要指标揭示了种群生态幅度、种群对资源利用和环境适应能力的差异以及种群之间的相互关系。杉阔混交林相比杉木纯林,其林下植被对资源的利用能力更强,森林资源结构更为合理,在经营模式上优于杉木纯林,为生物营造了一个良好的生态系统,因此探讨何种经营方式更有利于杉木人工林下植被的生存发展对于杉木人工林生物多样性资源保护和可持续利用具有一定的参考意义,也为森林经营措施的选择和优化群落结构提供了借鉴依据。
