稻虾轮作模式下江汉平原土壤理化性状特征研究*
2019-02-26朱建强李继福吴启侠
蔡 晨 李 谷 朱建强† 彭 亮 李继福 吴启侠
(1 长江大学农学院,湖北荆州 434025)
(2 中国水产科学研究院长江水产研究所,湖北荆州 434000)
水稻作为我国主要的粮食作物,年种植面积已达3 000万hm2[1]。传统的水稻种植方式通过施用化肥、农药来保证水稻的高产稳产,然而这种生产方式常造成养分资源浪费、农业环境污染,还会降低水稻品质、品味,甚至威胁食品安全[2]。为了充分利用稻田水、肥、气、热资源,不同地区根据气候和生产特点将水稻种植与动物养殖结合起来形成了独具特色的稻田养鱼、稻田养鸭和稻田养虾等稻田综合种养模式[3-5]。研究表明,与水稻单作相比,水稻种植与鱼、虾、鸭等水产动物养殖组成的互利共生复合生态农业系统具有诸多优点[6-11]。稻渔共生系统在世界很多国家均有分布,其中,稻田养虾模式在美国已开展数十年[12-14],在我国湖北省的江汉平原也较为普遍。2016年,湖北省就有20万hm2以上的稻田养殖小龙虾[15]。生产实践中,稻田养殖小龙虾多在水稻收后进行,即稻虾轮作,这种模式易于操作而得到广泛推广和应用。近年来,国内外学者对稻田综合种养模式进行了较为深入的研究,但多集中在稻鱼模式和稻蟹模式[16-19]。目前,关于稻虾模式的研究侧重稻田小龙虾养殖的技术和经济效益评估,对稻田土壤自身演化规律方面的研究尚较为薄弱[20]。为此,本研究基于前期调研数据,在长期实行稻虾轮作的同一农渠范围内,选择中稻单作和不同年限中稻/小龙虾轮作的稻田,进行0~20 cm、20~40 cm和40~60 cm土层理化性质分析,旨在揭示长期稻虾轮作下稻田土壤理化性质的演变规律,为稻虾田管理提供参考依据。
1 材料与方法
1.1 试验区概况
调查取样稻田位于湖北省石首市调关镇敕王庙村(29°41′25.2″N,112°39′40.8″E),毗邻长江、属江汉平原(图1),该村从20世纪90年代中期开始稻虾轮作生产。20年来,稻虾轮作集中在仅能种植一季水稻的低湖田上。土壤类型为第四纪沉积物发育的沼泽型水稻土,质地为黏壤土。稻虾种养单元呈矩形,宽20~30 m、长约50~100 m。根据1982年湖北省土壤普查结果显示,该区域土壤基本理化性质为:pH 7.28,有机质24.05 g·kg-1,全氮1.54 g·kg-1,全磷0.33 g·kg-1,全钾17.24 g·kg-1,碱解氮78.0 mg·kg-1,有效磷2.59 mg·kg-1,速效钾78.75 mg·kg-1。
1.2 稻虾轮作生产管理
稻虾轮作因地制宜利用了低湖田水土资源,且经济效益显著,逐渐由滨湖稻田向湖区地势较高的常规稻田发展,由此形成不同稻虾轮作年限的稻田。稻虾轮作模式中,水稻一般于每年6月中旬进行整地、插秧,9月底收获。水稻所施肥料为:尿素(N 46%)、过磷酸钙(P2O512%)和氯化钾(K2O 60%)。年均施肥量一般为N 150 kg·hm-2、P2O550 kg·hm-2和K2O 60 kg·hm-2。虾苗于当年10月水稻收获后,按每公顷15~20万尾的标准进行投放,至翌年3月幼虾在稻田中自然生长,每年3—5月份投放虾饲料,投放量平均为2 000 kg·hm-2,6月上旬捕捞成虾。捕虾完毕后(6月中下旬)进行下一季的中稻种植,在9月底中稻收割完毕灌水,进行下一季小龙虾养殖,依次循环。
稻虾轮作模式下,水稻收获后泡田养虾,稻草粉碎后全量还田,还田量约为6 750 kg·hm-2;普通中稻模式水稻收获后冬闲,不养虾。供试水稻品种为当地主推品种“黄花占”,小龙虾品种为克氏原螯虾。在4—5月稻田养虾期间,用生石灰进行水体消毒。按150 g·m-2用量在藻体聚集处撒生石灰,连续施用3次,每次间隔3~4 d,杀灭藻类,合计施用量约为340 kg·hm-2。
1.3 土样采集与测定方法

图1 采样点位置图Fig. 1 Location of sampling site
2017年9月水稻收获后,根据稻虾轮作年限,分别在中稻单作稻田(记为CK)以及稻虾轮作1 a、7 a、13 a、18 a和23 a的稻田(分别记为CR1、CR7、CR13、CR18、CR23)分层取土样。采用不锈钢土钻和环刀按S型5点法,分别采集0~20 cm、20~40 cm和40~60 cm土样,重复3次。土样带回实验室,剔除其中石块、根系等杂物,混匀、风干并分别过0.1 mm和0.149 mm筛备用。环刀采集的土样用于测定土壤容重和孔隙度,风干土样用于测定土样pH、全氮、碱解氮、有效磷、速效钾、阳离子交换量(CEC)和土壤易氧化有机碳(ROC)。
土壤pH测定采用电位法;土壤容重和孔隙度测定采用环刀法;土壤全氮测定采用凯氏定氮法;碱解氮测定采用碱解扩散法;有效磷测定采用0.5 mol·L-1NaHCO3浸提—比色法(UV-1600紫外可见分光光度计,Mapada,上海);速效钾测定采用1 mol·L-1醋酸铵浸提—火焰光度法(HG-3火焰光度计,北京检测仪器有限公司);阳离子交换量(CEC)测定采用乙酸钠-火焰光度法[21]。土壤易氧化有机碳(ROC)测定采用333 mmol·L-1高锰酸钾氧化法[22]。
1.4 数据分析
采用Microsoft Excel 2016处理数据和制图,DPS 15.10进行方差分析,最小显著差异(LSD)法进行显著性检验(P<0.05)。
2 结 果
2.1 不同年限稻虾轮作对土壤容重和孔隙度的影响
从空间尺度而言,各处理土壤容重自表层而下逐渐增加;从时间尺度而言,不同轮作年限各层土壤容重均较中稻单作的低,且随着轮作年限的延长而逐渐减小(图2)。从容重变幅而言,在0~40 cm土层中,随稻虾轮作年限的增加,土壤容重表现为先缓慢降低(0~1 a),随后显著降低(1~13 a),最后达到稳定,且土壤容重发生较大转折的时间为稻虾轮作的第7年与第13年之间。在40~60 cm土层中,土壤容重随轮作年限的延长逐渐降低,但降幅缓慢,表现为稻虾轮作13 a以内与中稻单作无差异。与中稻单作(CK)相比,稻虾轮作23 a后,0~20 cm、20~40 cm和40~60 cm土层的容重分别降低了12.2%、14.8%和1.26%。因此,长期稻虾轮作会显著降低土壤0~40 cm土层的容重,但对40~60 cm的土壤容重无显著影响。
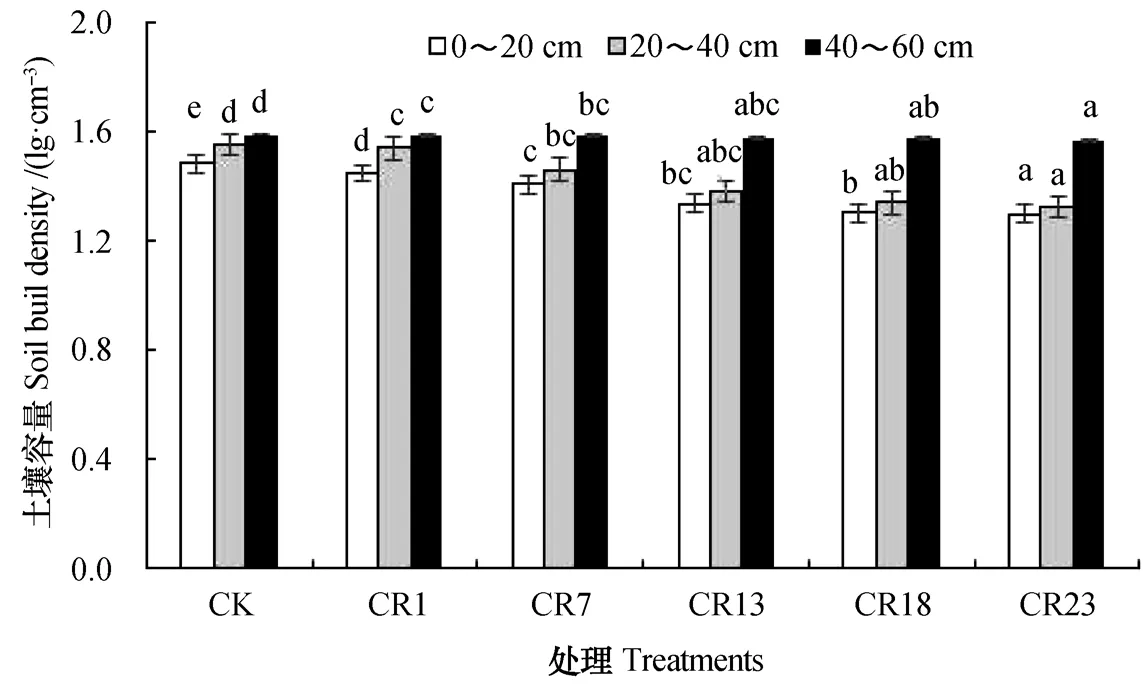
图2 不同年限稻虾轮作土壤容重变化Fig. 2 Changes in soil buik density in the paddy field with history of the rotation
由表1可知,土壤总孔隙度、毛管孔隙度和非毛管孔隙度均表现为随土层深度加深而减小的趋势。在0~20 cm土层中,与中稻单作相比,轮作1 a的毛管孔隙度和非毛管孔隙度缓慢增加,但总孔隙度显著增加;轮作1~13 a的毛管孔隙度、1~18 a期间的非毛管孔隙度和总孔隙度显著增加,随后趋于稳定。在20~40 cm土层中,土壤毛管孔隙度和总孔隙度的变化表现为:在轮作1~13 a期间缓慢增加,随后(18~20 a)趋于稳定。在40~60 cm土层中,土壤毛管孔隙度和总孔隙度的增加趋势更为缓慢,基本无变化。土壤非毛管孔隙度在20~60 cm土层中随年限的延长变化不明显。

表1 不同年限稻虾轮作对土壤毛管孔隙度、非毛管孔隙度和总孔隙度的影响Table 1 Effects of rice-crawfish rotation on soil capillary porosity, non-capillary porosity, and total porosity, relative to rotation history
2.2 不同年限稻虾轮作对土壤pH和阳离子交换量的影响
图3A结果显示,不同年限稻田土壤的pH介于7.28~8.29之间,且随土层深度增加呈升高的趋势。中稻单作和稻虾轮作1 a的稻田,其表层(0~20 cm)和下层土壤pH差异显著,轮作7 a以后,各层土壤pH差异不明显。与中稻单作相比,稻虾轮作可显著提高土壤表层pH。在稻虾轮作模式下,土壤pH随轮作年限增加而缓慢增加,各层土壤pH均表现为:23 a >18 a >13 a >7 a >1 a>CK。与CK相比,轮作23 a的3个土层pH分别提高了12.8%、5.8%和4.8%。
图3B结果显示,土壤阳离子交换量(CEC)由表层向下逐渐降低,不同土层CEC随轮作年限表现出不同的变化趋势。在0~20 cm土层,当轮作年限在13 a以内时,CEC 随轮作年限缓慢增加,之后趋于稳定;在20~40 cm土层,土壤CEC 随轮作年限的变化不显著;在40~60 cm土层,轮作7 a内CEC随轮作年限有缓慢增加趋势,随后变化不显著。
2.3 不同年限稻虾轮作对土壤全氮的影响

图3 不同年限稻虾轮作稻田土壤pH(A)和阳离子交换量(B)变化Fig. 3 Changes in soil pH (A) and cation exchange capacity (B) in the rice-crawfish field with history of the rotation
由图4可知,中稻单作时,土壤全氮含量自表层而下逐渐降低,且差异显著;而稻虾不同轮作年限各处理本身的20~40 cm和40~60 cm土壤全氮含量差异不显著。就时间尺度而言,各层土壤的全氮含量均随稻虾轮作年限延长而增加。与CK相比,轮作23 a稻田0~20 cm、20~40 cm和40~60 cm土层的全氮含量依次提高了128.6%、124.14%和96.2%。此外,从图4可以看出,土壤全氮含量发生较大转折的时间介于稻虾轮作的第13年和第18年之间。
就不同土层全氮含量变幅而言,在0~20 cm土层,随稻虾轮作年限的增加,土壤全氮含量有一个明显增加的过程。在20~40 cm土层,随稻虾轮作年限的增加,土壤全氮含量表现为先显著增加(0~1 a),随后缓慢增加(1~13 a),最后达到稳定的趋势。在40~60 cm土层,土壤全氮含量在轮作的0~1 a、7~13 a、18~23 a 3个时段增幅最为明显。

图4 不同年限稻虾轮作土壤全氮含量变化Fig. 4 Changes in soil total N content in the rice-crawfish field with history of the rotation
2.4 不同年限稻虾轮作对土壤速效养分的影响
由表2可知,土壤速效养分表层最高,向下逐渐降低;同时,土壤速效养分随着轮作年限延长均表现出增加的趋势。与中稻单作相比,轮作23 a后,0~20 cm、20~40 cm和40~60 cm土层中碱解氮、有效磷和速效钾均有显著增加。其中,0~20 cm土层中碱解氮、有效磷和速效钾分别增加35.6%、71.7%和117.1%,这表明长期稻虾轮作有利土壤速效养分含量提高,具体变化表现为:
1)0~20cm土层:随着轮作年限的增加,土壤速效养分均表现出先缓慢增加,随后显著增加的趋势。轮作7 a内,土壤碱解氮和有效磷含量缓慢增加,而速效钾在轮作1 a后显著增加。土壤碱解氮和有效磷在轮作13 a时趋于稳定,但速效钾一直呈显著增加趋势。
2)20~40cm土层:与对照相比,轮作7 a以上碱解氮才有显著增加;土壤有效磷含量在轮作1 a时显著增加,7~13 a时缓慢累积,轮作18 a以上又显著增加;土壤速效钾含量变化趋势与土壤有效磷类似。
3)40~60cm土层:与对照相比,土壤有效磷和速效钾含量在轮作1 a后均缓慢增加,13 a以上有显著增加的趋势,但不同轮作年限的土壤碱解氮含量之间差异较小,无明显规律。
2.5 不同年限稻虾轮作对土壤易氧化有机碳的影响
图5结果显示,土壤易氧化有机碳(ROC)由表层向下逐渐减少,且随轮作年限延长而逐渐增加。与中稻单作相比,稻虾轮作23 a后,0~20 cm和20~40 cm土层的ROC分别达到3.19 g·kg-1和3.13 g·kg-1,分别提高了95.7%和311.8%(P<0.05),40~60 cm土层处理间差异不显著。
3 讨 论
3.1 稻虾轮作下土壤物理性状变化及原因
土壤容重和孔隙度是土壤的重要物理性质,受到气候、土壤动物、微生物及耕作方式等的影响,对土壤透气性能、持水量、抗侵蚀能力和根系生长阻力有非常大的影响[23]。本研究结果显示,长期稻虾轮作会显著降低土壤0~20 cm和20~40 cm土层的容重(图2),并增加土壤孔隙度(表1);而对40~60 cm土层容重无明显影响。这是由于在稻虾种养过程中,会投喂水草和秸秆还田,这些有机物料在微生物和土壤酶的作用下逐渐腐解,且腐解产物的密度小于土壤自身密度,因此,能够与土壤颗粒结合形成稳定疏松态的团粒结构。此外,小龙虾在生长过程中排泄的粪便和掘洞行为也会加速这种团粒结构的形成和向较深层(20~40 cm)土壤迁移、转化的速率,从而改善土壤紧实程度。本研究还发现,土壤容重的降低和孔隙度的增大并未随着稻虾轮作年限的延长呈现无限下降和增长的趋势,而是表现出随着轮作年限延长趋于稳定的特点。如0~20 cm土层中,土壤容重和毛管孔隙度在轮作1~13 a期间呈现逐渐减小和增加的趋势,当轮作达到13 a之后基本保持稳定。在20~40 cm土层中,土壤容重在轮作1~18 a期间呈现逐渐减小,毛管孔隙度和总孔隙度在轮作1~18 a期间呈现逐渐增加的趋势,当轮作达到18 a之后保持稳定。这可能是每年秸秆摄入量和土壤有机物质消耗量达到动态平衡的结果,尚需要进一步进行试验研究。
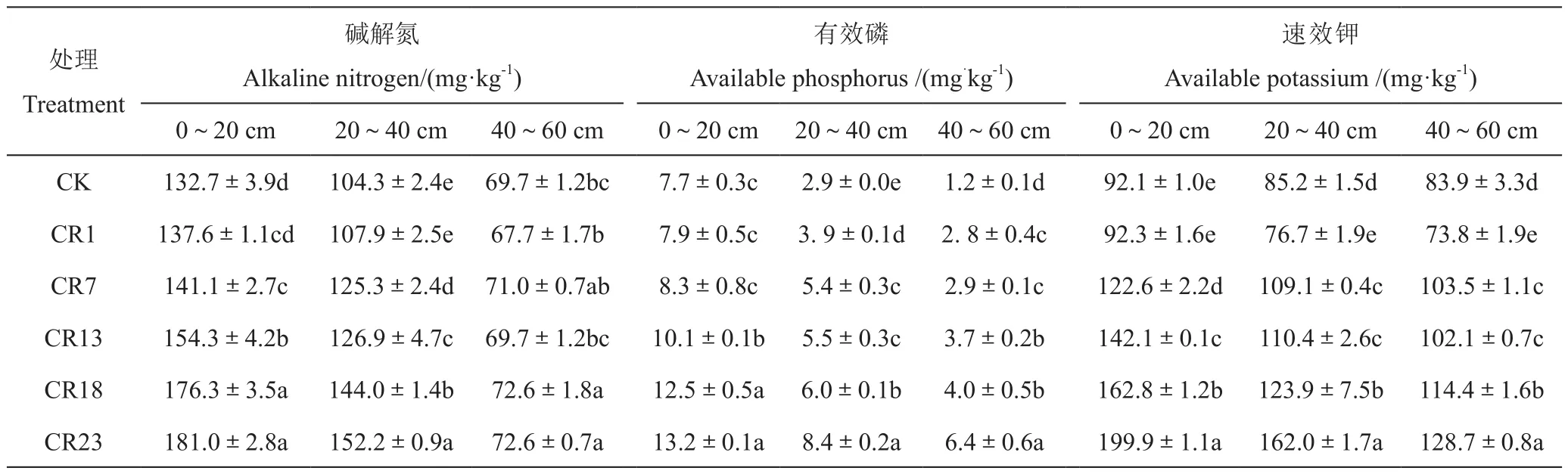
表2 不同年限稻虾轮作下土壤速效养分的变化Table 2 Change in content of soil available nutrients in the rice-crawfish field with history of the rotation
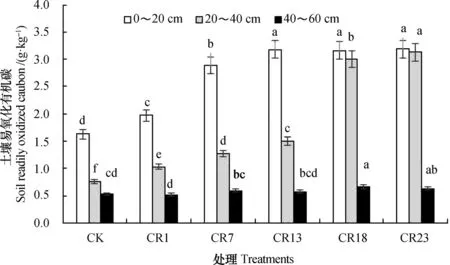
图5 不同年限稻虾轮作土壤易氧化有机碳含量变化Fig. 5 Change in soil ROC content in the rice-crawfish field with history of the rotation
3.2 稻虾轮作下土壤化学性状变化及原因
土壤pH是衡量土壤酸碱性的重要指标,受到土壤盐基状况、成土母质和人为活动等综合因素的影响,其中,土壤盐基状况主要取决于淋溶过程和复盐基过程的强弱[24]。本研究区,地下水位较高、成土过程稳定,故而,本研究区域土壤pH的变化主要受生物过程和人为活动的影响。水稻土是受人为活动作用影响剧烈、土壤质量变异最为显著的土壤。本研究表明,与第二次土壤普查数据相比,长期中稻单作对土壤pH无显著影响,而长期稻虾轮作可使土壤pH增加,稻虾轮作23 a时可使0~20 cm、20~40 cm和40~60 cm土层的pH较中稻单作依次增加12.8%、5.8%和4.8%(图3A)。这与佀国涵等[25]的研究基本一致。这一方面是在稻虾种养过程中,需要施用大量石灰来进行田间消毒,另一方面,小龙虾脱壳含有大量钙质,长期的生石灰和含钙较多的壳灰、贝壳粉等施入农田可中和田间H+,引起土壤pH缓慢升高,尤其是表层土壤表现更为明显[26-27]。
土壤阳离子交换量是指土壤胶体所能吸附和交换的各种阳离子的容量,直接反映土壤的保肥、供肥性能和缓冲能力,其量值与土壤胶体种类和含量有关。土壤有机质中的有机胶体为两性胶体,是土壤固相中阳离子交换量最大的部分,随着有机质含量的增加,土壤阳离子交换量也增加[24]。本研究表明,在表层(0~20 cm)土壤中,随着稻虾轮作年限的增加,土壤阳离子交换量随之增加,但对20~60 cm土层的土壤阳离子交换量无显著影响(图3B)。长期稻虾轮作模式下秸秆还田和小龙虾粪便进入了土壤表层,增加了土壤有机质的含量[25],从而使土壤表层阳离子交换量随之增加,而对深层土壤无显著影响,因此,深层土壤的阳离子交换量无明显变化。
本研究结果显示,土壤全氮、速效养分、易氧化有机碳含量和阳离子交换量均随土层深度的增加而呈递减趋势,表现为:0~20 cm > 20~40 cm >40~60 cm(图4,表2,图5)。这种变化趋势体现土壤养分含量的垂直分布,与生物富集、表聚作用密切相关。随着稻虾轮作年份的延长,土壤深层的养分含量也随之增加。这是因为在长期稻虾轮作过程中,小龙虾以还田稻草和投喂的饲料为食,食物残渣及小龙虾排泄物进入土壤,有效补充和提高了土壤养分含量[6,16]。小龙虾在稻田的掘穴行为改变了土壤的通气特性,加快了有机物质在有氧和厌氧界面的交换过程,从而使土壤深层中相对难降解的有机物逐渐被好氧微生物降解,显著影响了土壤中有机碳的转化[28]。此外,与中稻单作相比,短期稻虾轮作模式下20~60 cm土壤速效钾含量有一个明显的下降,之后随轮作时间的延长逐渐上升的过程。原因在于:(1)水稻根系从深层土壤中吸收钾素,然后再通过秸秆还田等形式将部分钾素归还于表层土壤,短期内深层土壤中的钾素得不到及时补充而减少;(2)周年淹水状态造成土壤部分溶液流失,引起养分损失[17,25,29]。
土壤易氧化有机碳的变动主要发生在土壤表层,40 cm以下深度变化不明显,且随着土壤深度的增加而显著降低[30-34]。Fanjul等[34]研究表明,螃蟹的掘穴行为可提高沼泽地沉积物中有机质含量及其生物有效性,增加易分解有机碳的比例和加快有机碳的矿化速率。曹凑贵等[35]研究表明,不同养虾年限的稻田土壤易氧化态碳含量显著高于常规水稻单作田,水溶性有机碳含量低于常规水稻单作田。本研究表明,长期稻虾轮作能够显著增加0~20 cm和20~40 cm土层中土壤易氧化有机碳含量,但对40~60 cm土层的土壤易氧化有机碳含量影响不明显(图5)。这与两方面的因素有关:(1)稻虾轮作期间,小龙虾排泄物在一定程度上增加了表层土壤易氧化有机碳的来源[25,33];(2)小龙虾活动对稻田起到中耕的效果,促进水稻根系正常生长,进而增加根系有机分泌物量和土壤稳定态腐殖质向易分解态转化[30,33,35]。
4 结 论
长期稻虾轮作会显著降低土壤0~20和20~40 cm土层的容重并提高其非毛管孔隙度、毛管孔隙度和总孔隙度,而40~60 cm土层无明显变化。稻虾轮作模式下,土壤pH随轮作年限增加而缓慢增加,各层土壤pH均表现为:23 a >18a >13 a >7 a>1 a >0(CK)。在0~20 cm、20~40 cm土层中,当轮作小于13 a时,CEC随轮作年限缓慢增加,之后趋于稳定;在40~60 cm土层中,轮作大于7 a后,CEC趋于稳定。各层土壤中的全氮含量随稻虾轮作年限延长而增加,与CK相比,稻虾轮作23 a后,0~20 cm、20~40 cm和40~60 cm土层的全氮含量分别增加128.6%、124.14%和96.15%。土壤速效养分随着轮作年限延长均表现出逐渐增加的趋势,这表明长期稻虾轮作有利提高土壤速效养分。稻虾轮作下,0~20 cm的土壤易氧化有机碳含量随轮作年限增加较快,13 a后变化不明显;在20~40 cm土层,土壤易氧化有机碳呈现平稳增加趋势,轮作23 a后保持平衡。因此,与中稻单作相比,稻虾轮作模式有助于改善土壤通透性、提高土壤肥力和增加农业产出效益。
