基于行为及气味对中缅树鼩亲缘识别的研究
2019-02-15侯东敏王政昆朱万龙
侯东敏,王政昆,朱万龙
(云南省高校西南山地生态系统动植物生态适应进化及保护重点实验室,云南师范大学 生命科学学院,昆明 650500)
动物生态学研究的前提和重要内容之一是行为的辨别和分类。区分认识动物的活动、休息、取食、攻击、标记等行为能够更为系统地理解其生态功能。动物的社会行为在很大程度上受到化学信号的影响,而动物社会行为发生的首要条件是对化学信号中所包含信息的辨识。近年来对动物行为的研究较为广泛,如通过对长爪沙鼠(Merionesunguiculatus)的研究表明,受限制的传播和社会组织可能在精细的空间内决定啮齿类动物的种群遗传基因结构[1];基于布氏田鼠(Lasiopodomysbrandtii)的研究表明通过声音环境切换的行为机制能够控制小鼠的生育率[2],对小鼠(Musmusculus)的社会行为研究发现早期环境强化不会导致雄性大鼠减少社会等级[3]。气味作为一种重要的化学信号,对于动物在复杂环境中生存以及繁殖具有重要作用。近年来对动物气味的研究颇多,如对雪豹(Pantherauncia)[4]、棕色田鼠(Lasiopodomysmandarinus)[5]、根田鼠(Microtusoeconomus)[6]等。在动物个体中嗅觉往往是对物种进行识别的主要感觉形式,包括性选择、亲缘识别和个人识别等[7-10]。
中缅树鼩(Tupaiabelangeri)属攀鼩目(Scandentia)树鼩科(Tupaiidae),分布于南亚、东南亚以及中国南部和西南部地区,为东洋界特有的昼行性小型哺乳动物。中缅树鼩是一种进化程度相对较高的哺乳动物,其生长速度快、体型小、驯化繁殖成本低,且在生物学特性上是除灵长类动物外较其他动物更接近于人的哺乳动物,因此成为近年来被广泛应用于生物学和医学等诸多领域的新型实验动物。为满足实验需求,对树鼩进行规模化驯化饲养十分迫切。本实验基于行为和气味探讨中缅树鼩的亲缘识别,为揭示中缅树鼩的亲缘识别提供重要的科学理论依据。
1 材料与方法
1.1 样本来源
本实验所采用的实验样本为实验室繁殖(中国医学科学院医学生物学研究所医学灵长类中心提供,实验动物生产许可证号:SCXK(滇)K2013-0001;实验动物使用许可证号:SYXK(滇)K2013-0001。)中缅树鼩12窝,每窝3只,共36只,每窝包括亲代雌性、子代雌性及子代雄性各1只。根据Kawamichi[11]对中缅树鼩的社会结构研究可知,一般情况下,一个雄性中缅树鼩的领域往往和一个或几个雌性的领域重叠。这说明中缅树鼩在自然条件下具有一雄一雌或者一雄多雌的繁殖方式[12]。结合饲养条件以及动物存活率等实际情况,故暂未选取亲代雄性进行亲缘识别研究,此部分内容将在后续的实验研究中进行深入探讨。
1.2 实验动物的处理
动物从中国医学科学院医学生物学研究所医学灵长类中心取回后于云南师范大学生命科学学院实验动物房(昆明)单笼室温分房间饲养,保证彼此间气味隔绝,避免气味交流。饲养笼规格为:120 mm×200 mm×300 mm,室温控制在20℃±2℃,光周期12L∶12D,用充足的饲料进行喂养。
1.3 行为观测
行为观察箱规格为:400 mm×400 mm×450 mm,材质为亚克力板,内部装有食槽(80 mm×80 mm×50 mm)以及休息暗盒(180 mm×80 mm×80 mm),并装有实时监控摄像头(名称:9路高清数字硬盘录像机;型号:ES-N6009),行为观察箱安置房间内装有15 W的暖色白炽灯用以照明。
实验前在行为观察箱内放置充足食物,包括苹果与面包虫等。将雄性动物尾巴根部剃毛,以便在实验观察时区分雌雄,实验时分别将各组亲属与非亲属动物个体两两置于观察箱内适应15~20 min,并通过实时监控录像设备采集动物生活视频信息。一次观测实验周期为24 h,期间保证实验动物不受外界环境影响。实验结束后,对行为观察箱进行消毒。
获取行为视频信息后,根据动物行为的表达特征,对动物在一定环境条件下出现的行为动作进行的辨认和记录。通过对24 h内动物个体的行为观察统计,计算某一行为产生的频次。
某时段内某行为产生频次百分比(%)=
1.4 气味识别
Y-迷宫由镀锡铁皮(厚16 mm)和透明亚克力板组成。Y-迷宫包括适应箱、刺激源箱(400 mm×300 mm×300 mm)、公共臂(700 mm×300 mm×300 mm)、选择臂(600 mm×300 mm×300 mm)及空气泵(名称:电磁式空气泵,型号:ACO-001,平均风速:38 L/min)组成。Y-迷宫安置房间内装有实时监控摄像头采集视频信息,15 W的白炽灯用以照明。实验后随机选取10 min进行统计,结果以s为单位进行统计分析。
1)尿液气味识别。实验前采集动物个体的新鲜尿液置于-20℃冰箱贮存;以规格为10 mm×10 mm×2 mm的方块脱脂棉作为尿液载体,保存于密封袋内避免污染,使用时用镊子夹出。
实验时先将被试动物个体置于Y-迷宫适应箱内,适应20 min,此时间段内解冻尿液样本,将方块脱脂棉置于由透明亚克力板制成的长方体小盒(规格为100 mm×50 mm×40 mm)上层中部位置,取1 mL尿液置于方块脱脂棉上使之充分吸收;后迅速将长方体小盒放置于刺激源箱内,将通风塑料管对准方块脱脂棉;开启空气泵。实验进行30 min,通过实时监控摄像头采集视频信息。实验完成后对Y-迷宫各部件消毒,对Y-迷宫安置房间进行消毒通风。
2)动物个体气味识别。实验时先将被试动物个体置于Y-迷宫适应箱内,适应20 min,随后将刺激源动物置于刺激源箱内,开启空气泵。后续步骤同1.4(1)。
3)生活暗盒气味识别。实验时先将被试动物个体放置于Y-迷宫适应箱内,适应20 min,随后将生活暗盒与刺激源箱对接,开启空气泵。后续步骤同1.4(1)。
1.5 统计分析
数据采用SPSS 13.0软件包统计,采用SigmaPlot 12.5软件包作图,并采用独立样本t检验(Independent-Samples T Test)完成。结果均以平均值±标准误表示,P<0.05表示差异显著,P<0.01表示差异极显著。
2 结果
2.1 亲属中缅树鼩行为节律分析
正常条件下,亲属中缅树鼩共同生活时的24 h行止具有明显的节律性,雄性与雌性个体的行为节律大致相同。
活动时间为白天,活动高峰为早晨和黄昏,活动呈双峰型。运动型活动开始时间为05:00,08:00—09:00达到活动的第一个高峰期,随后活动减少,持续到13:00出现活动的第二个高峰。20:00 后,几乎没有活动量(图1-A)。休息时间为18:00—07:00,尤其20:00—05:00处于休息高峰期(图1-B)。取食节律伴随活动节律增减,07:00—09:00和13:00—15:00为取食高峰(图1-C)。修饰行为发生于07:00—14:00,取食、休息后都会伴随修饰行为的发生。雌性修饰行为略多于雄性,对面部和四肢的修饰较多,对尾巴和身体其他部位修饰较少(图1-D)。
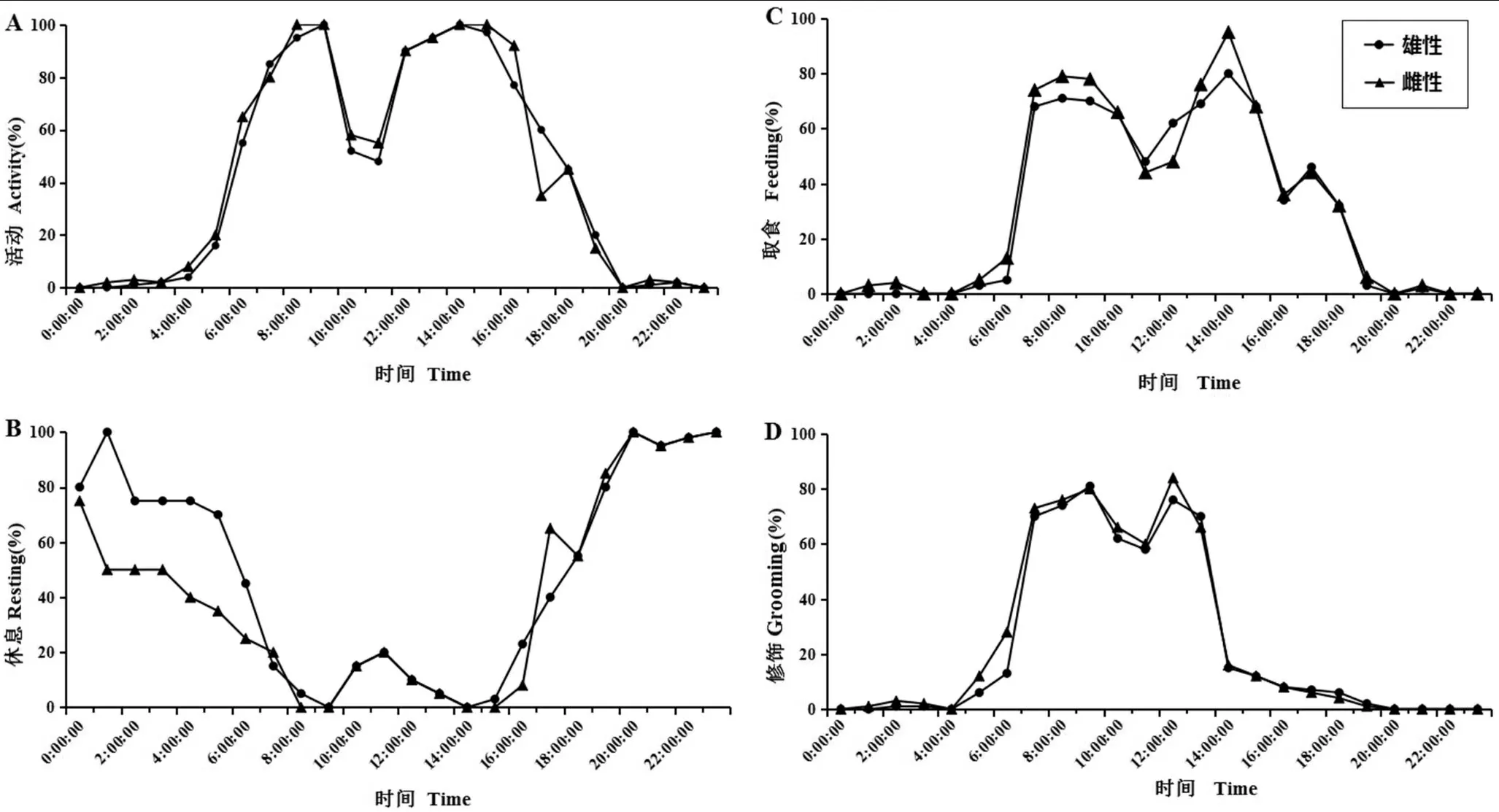
图1 亲属中缅树鼩行为日节律图(A:活动;B:休息;C:取食;D:修饰)Figure 1 A daily rhythm chart of the common life of Tupaia belangeri in a related relationship(A:Activity;B:Resting;C: Feeding;D:Grooming)

图2 非亲属中缅树鼩行为日节律图(A:活动;B:休息;C:取食;D:修饰)Figure 2 Daily rhythm chart of the common life of Tupaia belangeri in non-kin relationship(A:Activity;B:Resting;C: Feeding;D:Grooming)
2.2 非亲属中缅树鼩行为节律分析
非亲属中缅树鼩共同生活时的24 h行止与亲属中缅树鼩的结果大致相同,其具有明显的节律性,雄性与雌性个体的行为节律大致相同。
活动时间为白天,活动高峰为早晨和黄昏,活动呈双峰型。运动型活动开始时间为04:00,07:00—09:00达到活动的第一个高峰期,随后活动减少,持续到13:00出现活动的第二个高峰。20:00 后,几乎没有活动量(图2-A)。休息时间为18:00—07:00,尤其为18:00—05:00处于休息高峰期(图2-B)。非亲属中缅树鼩的取食节律与亲属中缅树鼩有所不同,未伴随活动节律增减,而在07:00~18:00间断性持续取食(图2-C)。修饰行为在07:00—10:00和12:00—14:00两个时段内较多发生(图2-D)。
2.3 亲属与非亲属中缅树鼩的行为比较
亲属中缅树鼩攻击行为的发生时间极显著小于非亲属中缅树鼩(t=-6.65,P<0.01);亲属中缅树鼩友好行为的发生时间显著大于非亲属中缅树鼩(t=3.13,P<0.05);亲属中缅树鼩标记行为的发生时间显著小于非亲属中缅树鼩(t=-2.44,P<0.05)。结果见表1。

表1 亲属与非亲属中缅树鼩的行为比较Table 1 Comparisons of the behaviors of kin and non-kin Tupaia belangeri
注:数值为平均值±标准误。P<0.05表示两者之间差异显著;P<0.01表示两者之间差异极显著
2.4 中缅树鼩尿液气味识别
亲属中缅树鼩的尿液与对照(不做任何处理的空刺激源箱)做刺激源时,被试子代雄性(t=5.36)、子代雌性(t=4.09)、亲代雌性(t=25.19)中缅树鼩均极显著(P<0.01)的选择中缅树鼩的尿液;非亲属中缅树鼩的尿液与对照做刺激源时,被试子代雄性(t=5.07)、子代雌性(t=3.4)中缅树鼩均极显著(P<0.01)选择中缅树鼩的尿液,被试亲代雌性(t=2.73)中缅树鼩显著(P<0.05)选择中缅树鼩的尿液;亲属与非亲属中缅树鼩的尿液做刺激源时,被试子代雄性(t=4.27)、子代雌性(t=4.88)、亲代雌性(t=7.19)中缅树鼩均极显著(P<0.01)选择亲属中缅树鼩的尿液。结果见图3。
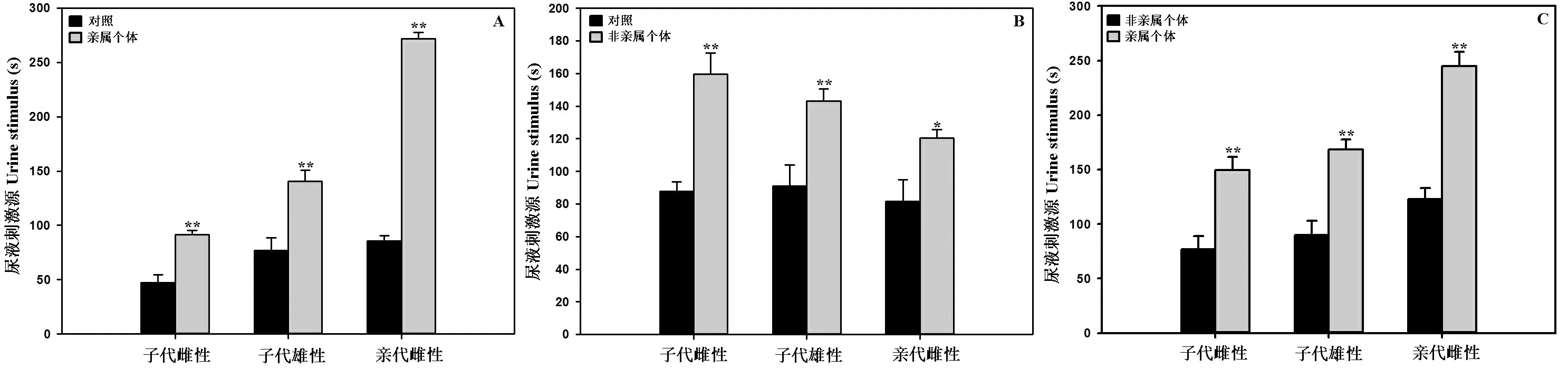
图3 Y-迷宫中中缅树鼩对亲属与对照(A)、非亲属与对照(B)、亲属与非亲属(C)尿液的选择
Figure 3 Selection of the urine in the Y-type labyrinth with kin and control (A),non-kin and control (B),kin and non-kin (C) of theTupaiabelangeri
数值为平均值 ± 标准误。*:P<0.05表示两者之间差异显著;**:P<0.01表示两者之间差异极显著。下同
2.5 中缅树鼩个体气味识别
亲属中缅树鼩与对照(不做任何处理的空刺激源箱)做刺激源时,被试子代雄性(t=5.36)、亲代雌性(t=26.12)中缅树鼩均极显著(P<0.01)的选择中缅树鼩,被试子代雌性(t=2.24)中缅树鼩显著(P<0.05)选择中缅树鼩;非亲属中缅树鼩与对照做刺激源时,被试子代雄性(t=14.54)、子代雌性(t=4.74)及亲代雌性(t=13.85)中缅树鼩均极显著(P<0.01)选择中缅树鼩;亲属与非亲属中缅树鼩做刺激源时,被试子代雄性(t=6.72)、子代雌性(t=5.33)中缅树鼩均极显著(P<0.01)选择亲属中缅树鼩,被试亲代雌性(t=2.11)中缅树鼩显著(P<0.05)选择亲属中缅树鼩。结果见图4。
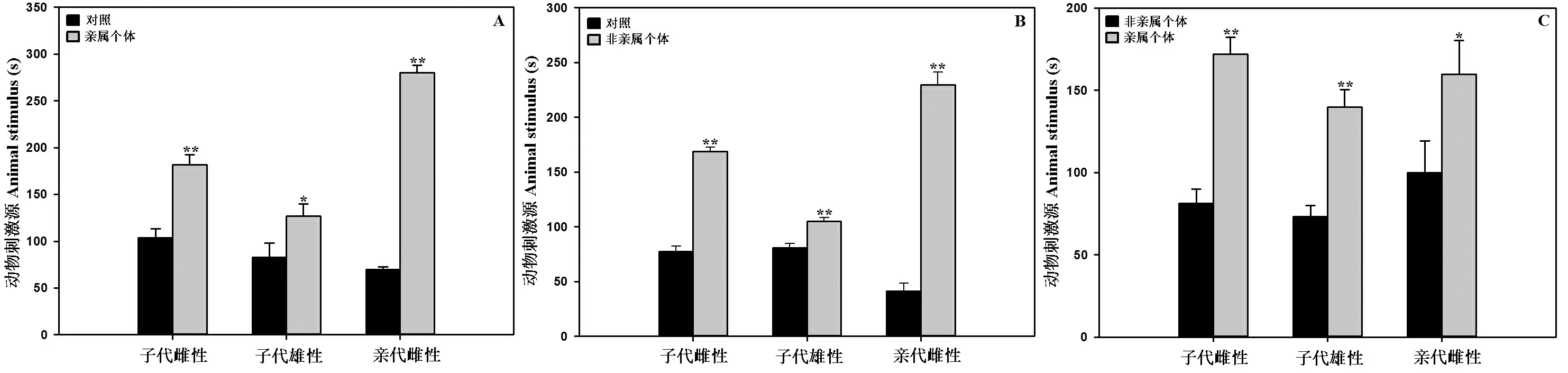
图4 Y-迷宫中中缅树鼩对亲属与对照(A)、非亲属与对照(B)、亲属与非亲属(C)身体气味的选择
Figure 4 Selection of the body odor in the Y-type labyrinth with kin and control (A),non-kin and control (B),kin and non-kin (C) of theTupaiabelangeri
2.6 中缅树鼩暗盒气味识别
亲属中缅树鼩的居住暗盒与对照(不做任何处理的空刺激源箱)做刺激源时,被试子代雄性(t=26.51)、亲代雌性(t=16.47)中缅树鼩均极显著(P<0.01)的选择中缅树鼩的居住暗盒,被试子代雌性(t=2.82)中缅树鼩显著(P<0.05)的选择中缅树鼩的居住暗盒;非亲属中缅树鼩的居住暗盒与对照做刺激源时,被试子代雄性(t=11.40)中缅树鼩极显著(P<0.01)选择中缅树鼩的居住暗盒,被试子代雌性(t=2.12)、亲代雌性(t=2.73)中缅树鼩显著(P<0.05)选择中缅树鼩的居住暗盒;亲属与非亲属中缅树鼩的居住暗盒做刺激源时,被试子代雄性(t=4.20)中缅树鼩极显著(P<0.01)选择亲属中缅树鼩的居住暗盒,被试子代雌性(t=2.50)及亲代雌性(t=2.14)中缅树鼩显著(P<0.05)选择亲属中缅树鼩的居住暗盒(见图5)。
3 讨论
近年来,动物行为的研究逐渐进入国内外学者的视线,成为当下研究的热点问题,但大多数是对啮齿类动物和大型兽类社群行为[13-14]、繁殖行为[15]、攻击行为[16]的研究,而对中缅树鼩的行为研究鲜有报道。
本研究中亲属中缅树鼩个体取食行为呈双峰型,说明亲属中缅树鼩雌雄个体不产生食物竞争关系,能够和睦相处,这与本研究组对单独笼养中缅树鼩取食行为的研究相符[17];而非亲属中缅树鼩的雌雄个体产生食物竞争,因此争相取食,致使取食行为呈单峰型。因此推测导致此差别的原因是中缅树鼩能够进行亲缘识别,从而分辨亲属与非亲属中缅树鼩。

图5 Y-迷宫中中缅树鼩对亲属与对照(A)、非亲属与对照(B)、亲属与非亲属(C)居住暗盒的选择
Figure 5 Selection of the cassette in the Y-type labyrinth with kin and control (A),non-kin and control (B),kin and non-kin (C) of theTupaiabelangeri
通过对中缅树鼩行为频次的统计分析发现,亲属中缅树鼩雌雄个体之间友好行为的发生时间显著多于非亲属雌雄个体,而攻击行为和标记行为却显著低于非亲属雌雄个体,尤其攻击行为达到极显著状态。相关研究证实动物个体间亲缘关系越近,友好行为发生频次越高[18-19],这说明个体能够通过与亲缘合作提高广义适合度。大量研究表明个体间的友好行为不仅可以作为缓和攻击之后社会关系和减弱竞争的方式,它也能增加种群内个体获得资源的概率[20]、维持社会地位[21]以及增加繁殖成功率[22]等。
小型哺乳动物利用气味进行化学通讯主要是通过对其他个体的气味差异进行嗅觉识别从而完成个体识别,尿液、粪便及腺体分泌物等在识别过程中起到了气味源的作用[23-24]。本研究气味识别实验中的刺激源为尿液和暗盒时,子代雄性对其亲属个体和对照组的选择明显低于非亲属和对照组,可能说明雄性中缅树鼩对其亲属个体尿液及暗盒的气味识别能力相比雌性中缅树鼩较弱;而刺激源为动物个体时子代雄性对其亲属个体和对照组的选择却明显高于非亲属和对照组,由此推测动物个体作为刺激源时雄性中缅树鼩除嗅觉通讯外,还可能通过视觉及听觉等方式进行通讯,且此能力比雌性个体较强。嗅觉气味源信号在小型哺乳动物的化学通讯中具有重要作用,通过嗅觉对气味信号的识别和记忆的能力影响动物的社会等级、亲缘识别、领域等社会行为[25-27]。本研究无论以尿液、暗盒、个体中任意一种方式作为刺激源时,中缅树鼩均能显著识别亲属个体,这说明中缅树鼩能够通过气味物质进行识别。
综上所述,亲属与非亲属中缅树鼩的活动、休息、取食、修饰行为表现出昼夜节律性;亲属与非亲属的取食行为频次趋势有所不同。中缅树鼩更显著偏向于选择与其具有亲属关系动物的尿液、个体及暗盒气味。以上结果表明亲属和非亲属中缅树鼩的行为模式存在差异,尿液等气味因素会对中缅树鼩的亲缘识别产生影响。
