蚤状溞caspase-3基因在不同环境条件下的表达分析
2019-02-15吴乐乐叶功照童巧琼王丹丽
吴乐乐,叶功照,童巧琼,王丹丽,2
(1.宁波大学 海洋学院,宁波 315211;2.宁波大学 应用海洋生物技术教育部重点实验室,宁波 315211)
蚤状溞(Daphniapulex)是一种常见的淡水枝角类,俗称红虫,属于节肢动物门(Arthropoda),甲壳纲(Crustacea),双甲目(Diplostraca),广泛分布于我国的淡水湖泊、河流中。蚤状溞因其易繁殖、适应性强、生长周期短、产仔量高、蛋白质含量高、易培养等特点,成为许多水产动物育苗中必不可少的关键活体饵料。近几年来,随着我国水产养殖业的蓬勃兴起及苗种生产的不断发展,对枝角类的需求量越来越大,故深入研究蚤状溞,有着重大的意义。在枝角类的分子水平研究中,蚤状溞是第一个基因组序列被测出来的溞类[1]。
枝角类生殖方式十分独特,在环境适宜条件下,其主要以孤雌生殖(单性生殖)方式繁殖,雌体所产卵子不需受精而直接发育成子代;而当其受到食物匮乏、温度骤变、种群密度过大等环境因素胁迫时,雄性个体出现,进而生殖方式转变为两性生殖[2]。然而,本实验室在蚤状溞培养过程中发现,即使保持稳定的最适环境,随着培养时间的延长,溞体也会逐渐进入衰老状态,同时伴随着更多的两性溞和卵鞍的出现。这一现象与Fontana等[3]认为衰老是随着时间的推移,伴随着分子、细胞和组织器官的损伤,生物体的机能和结构逐渐衰退、老化的复杂生物学过程,最终导致生物体死亡的观点相一致。因此我们推测枝角类进行有性生殖主要是由内部因素决定,环境变化仅在一定程度上起诱导作用,即在环境条件好的情况下随着溞年龄的增长,衰老进程的发展,最后从量变发生了质变,进入两性生殖模式。
Caspase(Cysteinylaspantate specific pnoteinase)半胱氨酸天门冬氨酸蛋白酶家族主要存在于动物细胞中,参与细胞因子的成熟、衰老、细胞生长和分化调控,在凋亡信号传导途径中发挥重要的功能[4]。Caspase家族与真核细胞有着密切的联系,无论是在低等的无脊椎动物,还是高等的哺乳动物中均高度保守,而在酵母和植物这类没有典型的细胞凋亡现象的生物中并没有发现,由此可见细胞凋亡的发生很大程度上依赖于Caspase家族蛋白酶的活化。而Caspase-3是迄今为止在该家族中研究较为透彻,且起主要效用的一种蛋白,在凋亡的有序级联反应中,Caspase-3处于下游,是多种凋亡刺激信号传递的汇聚点,通过信号触发蛋白底物裂解,细胞解体,最终完成细胞凋亡[5],可见Caspase-3是凋亡的关键执行者,它的活化也是细胞进入不可逆凋亡阶段的标志[6]。Caspase-3还可以激活促凋亡蛋白,拮抗抑凋亡蛋白[7],破坏细胞平衡状态及修复机制,加速细胞凋亡[8]。
已有许多研究表明Caspase-3的活性与年龄相关,随着年龄增长,C57B/6J小鼠大脑初级听皮层中caspase-3的表达增多,凋亡抑制蛋白XIAP的表达量减少[9]。胡艳丽等人的研究结果显示在衰老模型动物大脑海马中,有大量caspase-3蛋白表达,且较对照组明显增多[10]。Zhao等[1]和Su等[12]也提出caspase-3在衰老和阿尔茨海默病(AD)的某些特定脑区异常高表达。还有学者研究发现caspase-3基因受环境因子的影响很大,是一种环境敏感型基因,环境的变化会给予细胞一定的刺激,从而产生凋亡信号[13-14]。多因素方差分析显示,养殖条件、温度变化及变化时间、非离子氨浓度和胁迫时间对凡纳滨对虾(Litopenaeusvannamei)血淋巴和肝胰腺Caspase-3 活性均具有显著影响(P<0.05)[14]。目前,caspase-3基因表达定量也被证实可用于测定细胞凋亡水平,表达量越高则细胞凋亡水平越高[13]。
本研究拟通过分析蚤状溞caspase-3基因在不同温度、种群密度和食物种类下的表达水平差异,以及3种环境因子对Caspase-3酶活变化的影响,结合整体原位杂交检测caspase-3基因在不同生殖状态下的表达位点,初步探索蚤状溞caspase-3基因在不同环境条件下的时空表达,为今后深入探究衰老相关的caspase-3基因是否参与调控枝角类生殖模式转换的分子机制提供一定的理论参考和依据。
1 材料与方法
1.1 实验材料驯化与培养
试验所用的蚤状溞是由本实验室鉴定并培养保种的,成体体长(3.0±0.3)mm。挑选其中健康并且活力较强的蚤状溞,在玻璃培养箱内进行培养,每2~3 d添加“Banta粪土培养液”(1.5 g兔子粪,2 g新鲜稻草,20 g沃土和1 L水,煮沸冷却取上清或过滤)进行驯化,pH值控制在7.0~7.5的范围内,培养温度(25±1)℃,光周期为12 h光照和12 h黑暗,并且每3 d更换1/3体积的新鲜培养液。驯化完成后,在500 mL的玻璃烧杯进行培养,每杯加入“Banta粪土培养液”500 mL,从驯化好的蚤状溞中挑选出大量怀卵的蚤状溞放入培养箱中进行培养,培养条件与之前相同。过1~2 d会有小溞出生,挑选同一批出生的小溞按实验条件进行培养。
1.2 不同温度下蚤状溞的培养
在15℃、20℃、25℃、30℃和变温(10℃~18℃)5组不同温度下,每组用6个500 mL的烧杯培养蚤状溞,每只烧杯接种150只活力好,且大小相似的同一天出生的蚤状溞。实验设3个平行。其中3个烧杯溞用于测定Caspase-3酶活,另外3个烧杯溞用于测定caspase-3 mRNA表达量)。
1.3 不同密度下蚤状溞的培养
设计5组不同密度,用500 mL的玻璃烧杯培养蚤状溞,分别接种50、100、150、200 和300只活力好,且大小相近的同一天出生的蚤状溞,培养液为“Banta粪土培养液”,温度控制为23℃~25℃。实验设3个平行。
1.4 不同食物投喂下蚤状溞的培养
设计2组不同食物,用500 mL的烧杯培养蚤状溞,每只烧杯接种150只活力好,且大小相似的同一天出生的蚤状溞,分别用“Banta粪土培养液”和浓度为1×105~1×106cell/mL小球藻(Chlorellavulgaris)进行培养,温度控制为23℃~25℃。实验设3个平行。
以上试验均培养8 d(绝大部分在第8天已经性成熟,性成熟的标志是怀卵),pH值控制在7~8之间,每天观察记录溞的生长情况,并剔除刚出生的小幼溞,在第4天时换1/3的培养液。
1.5 蚤状溞总RNA的提取和第一链cDNA的合成
各实验组收集不同培养条件下蚤状溞样品150~200只,去水称重。用RNA提取试剂盒(RNA Extraction Kit,Axygen)分别提取总RNA,测定RNA的浓度,并用1%的琼脂糖凝胶电泳检测总RNA的完整性。每组各取1 μg RNA,以上述提取的总RNA为模版,用HiFiscript cDNA第一链合成试剂盒反转录合成cDNA第一链,立即实验或-20℃保存备用。
1.6 caspase-3基因荧光定量
以蚤状溞不同模板的cDNA作为模板,用特异性引物CaspRT-F(CGTTGAAGATGTTTGGATGCC)/CaspRT- R(GATGAACAGGAGCCACTGC)和18S-F(GTGCGTCGTTGTTGTATCTGC)/18S-R(TCCATGCTGCGATATTCAGG)进行荧光定量PCR。数据采集在LC480上完成,采用2-△△Ct法[15]进行引物的效率检测,若PCR产物的溶解曲线没有杂峰,则显示产物特异性好。
1.7 Caspase-3酶活的检测
收集不同实验培养条件下蚤状溞样品,裂解后离心获得上清。根据Caspase-3试剂盒说明书,通过酶标仪测定吸光度A405下样品数据,同时做空白对照。利用标准曲线换算样品催化产生pNA的量。酶活力=[吸光度A405值(2 h)-吸光度A405值(0 h)]/样品重量(mg)/ 1.5×105
1.8 数据处理
1.9 caspase-3 RNA探针的制备
首先根据已获得的caspase-3 cDNA全长(GenBank登录号:KY622004),设计合成探针的引物CaspPro-F(GATGAACAGGAGCCACTGC)/CaspPro-R(CTATTTCCTTGACAGCCACTG),构建pGEM-T-caspase-3重组质粒。将pGEM-T-caspase-3重组质粒酶切线性化,采用AxyPrep PCR纯化试剂盒回收线性化的模板质粒,用1.0%DEPG处理的去离子水中重悬于-20℃保存备用。
将纯化的线性化质粒作为体外转录的 DNA 模板,参照DIG RNA Labeling Mix和Riboprobe®System—SP6/T7实验程序,体外转录合成 DIG 标记的Caspase-3正反义RNA 探针。用无RNA酶活性的DNA酶处理除去DNA,然后加入2.5 μL的4 mol/L LiCl(DEPC水配制)和75 μL冰预冷的无水乙醇,轻轻混匀后于-20℃放置过夜。再在4℃,12 000 r/min下离心30 min,弃上清。加入100 μL预冷的70%乙醇(DEPC 水配制)清洗沉淀,4℃,12 000 r/min离心5 min,弃上清。向 RNA沉淀中加入30 μL RNase Free ddH2O和1.5 μL RNase inhibitor自然溶解,进行琼脂糖电泳检测和浓度测定后,将其放置在-80℃保存备用。
1.10 整体原位杂交检测caspase-3基因的表达定位
取在-20℃脱水保存的孤雌成溞和在高温和高密度实验中出现卵鞍的两性雌溞,在梯度浓度甲醇中逐步复水至PBST,每次洗脱5 min。在含100%1 ×PBST中漂洗10 min洗脱2次。用含10 μg/mL蛋白酶K的PBST在37℃下消化15 min。然后在4%PFA中固定20 min,PBST漂洗2次。经过1∶1预杂交液/PBST漂洗5 min,于68℃预杂交2 h(预杂交液∶RNA探针=1∶100,探针浓度约4 ng/μL)。预杂交液做空白对照,正义探针作阴性对照,反义RNA探针为实验组。70℃杂交过夜,不超过16 h。杂交后于72℃下,依次经过50% 4×SSC+50%MABT洗20 min,50%2×SSC+50% MABT洗20 min,2×SSC洗20 min,0.2×SSC洗40 min 2次,0.5×PBST室温洗20 min 2次。室温下用封闭液(MAB Block)封闭2 h。用Anti-DIG单抗按1∶5000将抗体加入到封闭液中,4℃下过夜处理。经过PBST漂洗8次,每次15 min。TSM1漂洗2次,每次20 min,TSM2漂洗2次,每次20 min。然后用NBT/BCIP室温下避光染色10~20 min。
用1×PBST漂洗,放在4%PFA中,4℃保存。结果用显微镜进行图像采集,拍照记录caspase-3基因在溞体中的表达定位情况。
2 结果
2.1 蚤状溞存活率和生长情况观察
试验期间各实验组的存活概率都在95%以上。
温度组:室温组和15℃实验组蚤状溞个体较小,且在8 d之内只有怀卵情况而没有小溞产生,其他各个实验组都有小溞产生。30℃组出现卵鞍。
密度组:200 ind/500 mL和300 ind/500 mL组的蚤状溞几乎都在杯底并挤成一团只有个别溞在其他水层游动,且密度越高蚤状溞个体也越小。50 ind/500 mL组在第5天就有部分蚤状溞生产,密度越高所产小溞越少。300 ind/500 mL组出现卵鞍。
食物组:第8天时以Banta培养液培养的蚤状溞溞体明显大于以小球藻培养的蚤状溞,同时小球藻组只产生了较少的幼溞,而Banta培养液组生产的幼溞明显较多。
2.2 蚤状溞caspase-3 mRNA在不同环境条件下的表达
根据测定的cDNA浓度值,选取10倍稀释度进行Real-Time PCR扩增反应,获得相对定量标准曲线,引物18S-F/R和CaspRT-F/R的扩增标准曲线分别如图1和图2所示。qPCR对扩增引物的特异性要求高,从引物扩增标准曲线图中得到,caspase-3相关系数(R2)为0.9986,18S(R2)为0.9976,越接近1可信度越高;标准曲线斜率分别为-3.48和-3.60,将其换算为引物的扩增效率(E)分别为1.05和1.08,越接近1越理想;两者的扩增效率应保持一致。R2、E及曲线斜率均在正常范围内,所以可拿来当实时荧光定量的引物。
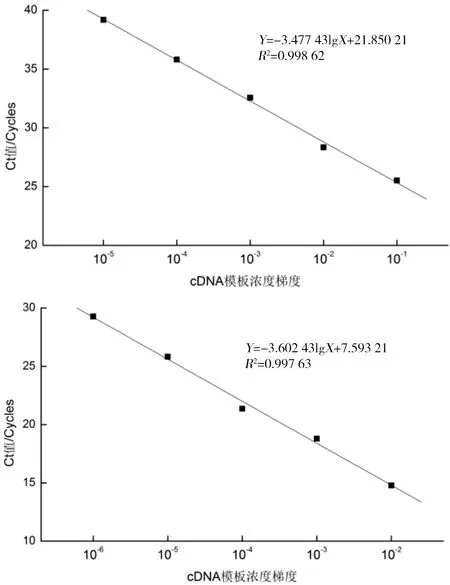
图1 目的引物caspase-3和参照18S的扩增标准曲线图Figure 1 Standard curves of target primer caspase-3 and 18S reference amplification
图2实时荧光定量检测图Figure 2 Real time fluorescence quantitative detection map
2.2.1 蚤状溞caspase-3 mRNA在不同温度下的表达
利用qPCR相对定量方法分析了蚤状溞caspase-3 mRNA在不同温度下蚤状溞体内的表达,基因的表达特征图谱,见图3-a、b所示表达电泳规律一致。caspase-3在30℃时的表达量最高,高于其他生长阶段(P<0.05),总体上caspase-3的mRNA表达量随着温度的增长呈上升趋势,室温组的表达量则介于20℃组与25℃组之间。
2.2.2 蚤状溞caspase-3 mRNA在不同种群密度下的表达
从图4可以看出,caspase-3 mRNA的表达量随着密度的上升总体是呈上升的趋势(P<0.05),在100 ind/500 mL的时caspase-3 mRNA的表达量为最低。
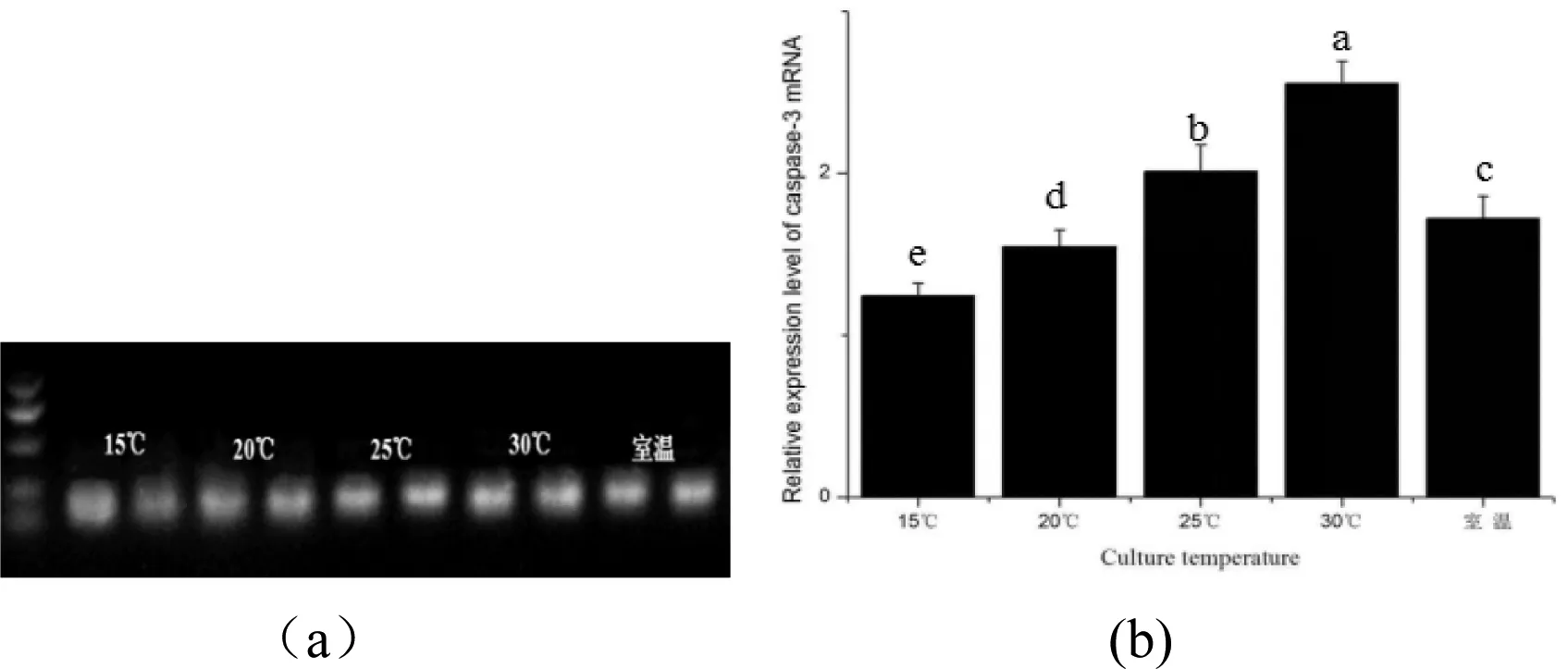
图3 蚤状溞caspase-3 mRNA在不同温度下的电泳图和表达情况Figure 3 Electrophores(a) and expression(b) of caspase-3 mRNA in D.pulex at different temperatures
注:图中不同字母表示有显著性差异(P<0.05);下同
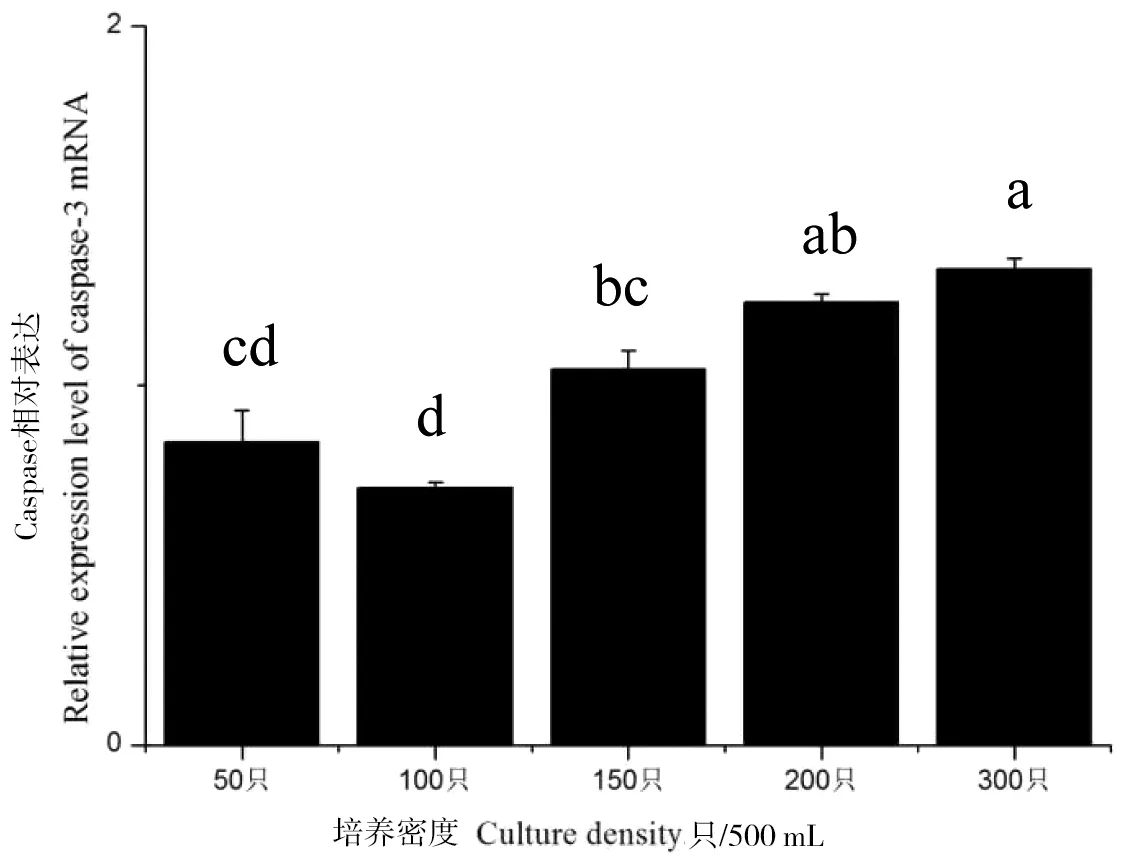
图4 蚤状溞caspase-3 mRNA在不同密度下的表达情况Figure 4 Expression of caspase-3 mRNA in D.pulex under different densities
2.2.3 蚤状溞caspase-3 mRNA在不同食物下的表达
从图5可以看出,在本实验中“Banta粪土培养液”组caspase-3 mRNA的表达量低于小球藻组,但无显著差异(P>0.05)。
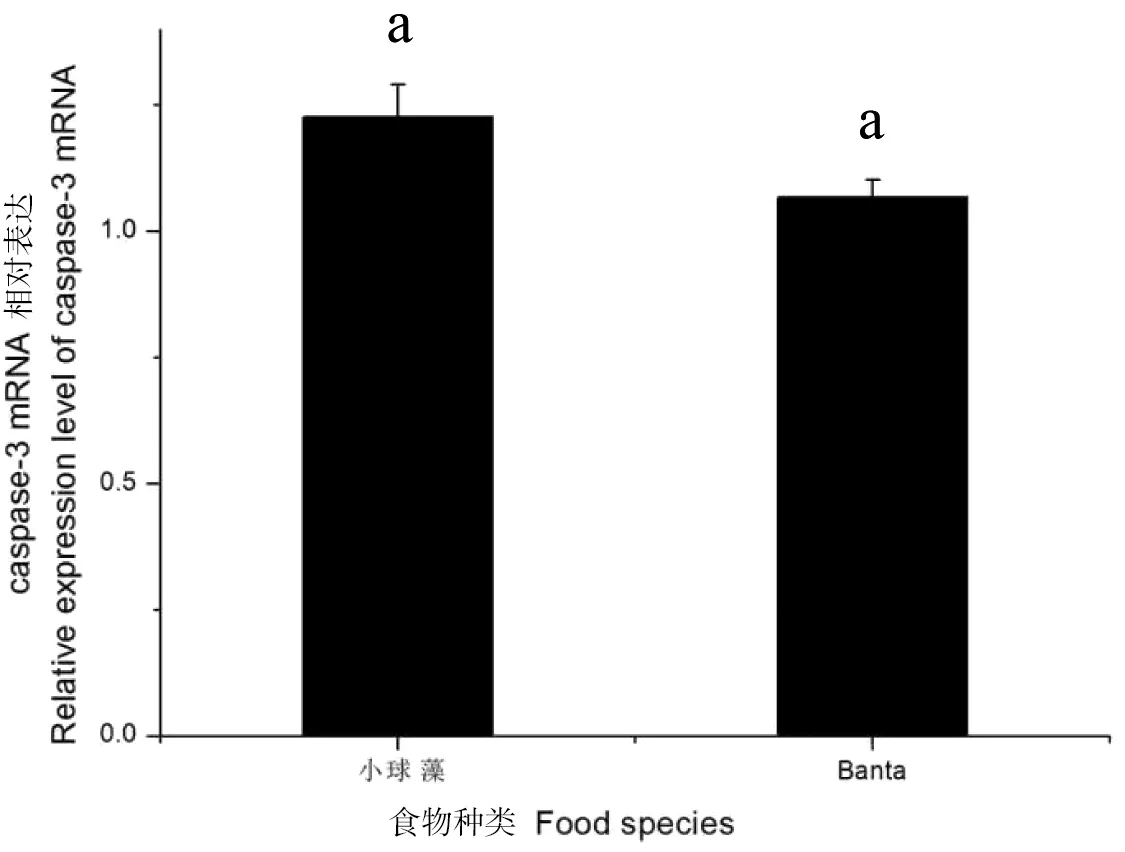
图5 蚤状溞caspase-3 mRNA在不同食物下的表达情况Figure 5 Expression of caspase-3 mRNA in D.pulex with different food specoes
2.3 蚤状溞Caspase-3酶活性在不同环境因子中的变化
2.3.1 蚤状溞Caspase-3酶活性在不同温度下的变化
为进一步研究蚤状溞Caspase-3蛋白酶在蚤状溞体内的表达情况,对Caspase-3的酶活性进行了测定,结果见图6。可以看出蚤状溞在15℃~20℃时Caspase-3酶活性较低,并在15℃时Caspase-3的酶活性最低为12.63 nmol/mg/h,30℃时最高为83.97 nmol/mg/h。除变温组总体随着温度的升高蚤状溞Caspase-3的酶活性也随之显著升高(P<0.05)。与caspase-3 mRNA在不同温度下的表达趋势一致。

图6 蚤状溞Caspase-3的酶活性在不同温度下的变化Figure 6 Changes of enzyme activity of Caspase-3 in D.pulex at different temperature
2.3.2 蚤状溞Caspase-3酶活性在不同密度下的变化
从图7可以看出,蚤状溞在密度为50~150 ind/500 mL时Caspase-3酶活性最低,300 ind/500 mL时最高达33.85 nmol/mg/h,在密度大于150 ind/500 mL时酶活性提高迅速。总体上看随着密度的升高蚤状溞Caspase-3的酶活性也随之显著升高(P<0.05)。与caspase-3 mRNA在不同密度下的表达趋势一致。
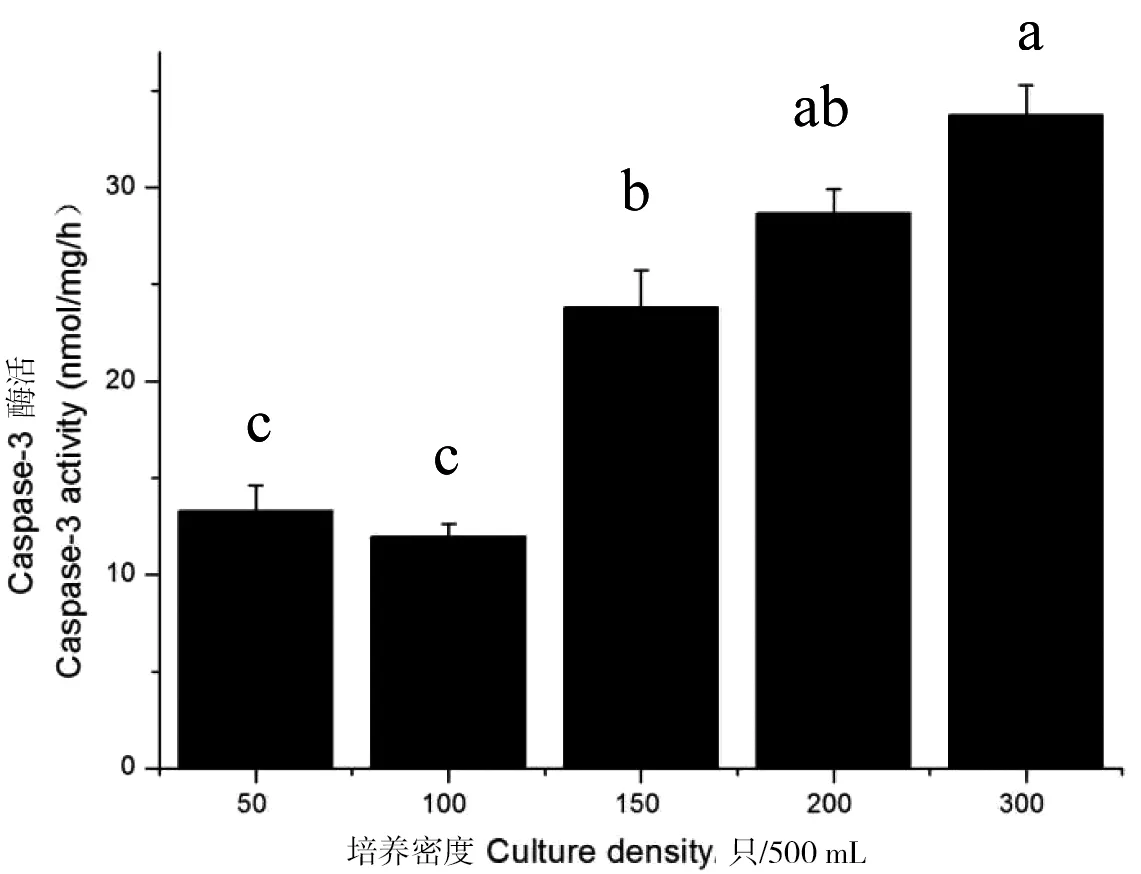
图7 蚤状溞Caspase-3的酶活性在不同密度下的变化Figure 7 Changes of enzyme activity of Caspase-3 in D.pulex at different densities
2.3.3 蚤状溞Caspase-3酶活性在不同食物条件下的变化
从图8中可以看出,在本实验中“Banta粪土培养液”组Caspase-3酶活性低于小球藻组,但无显著差异(P>0.05),结果与caspase-3 mRNA在不同食物条件下的表达趋势一致。
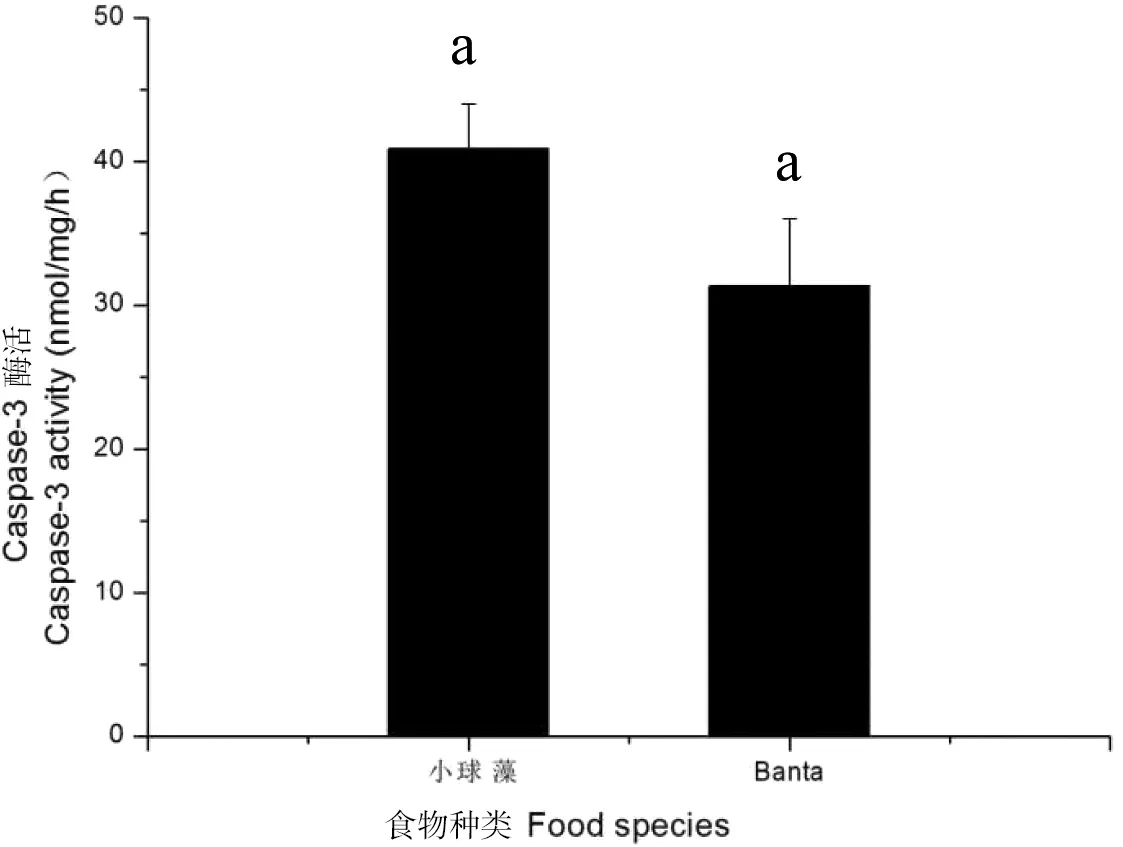
图8 蚤状溞Caspase-3的酶活性在不同食物条件下的变化Figure 8 Changes of enzyme activity of Caspase-3 in D.pulex with different food species
2.4 整体原位杂交检测caspase-3基因在蚤状溞不同生殖状态下的表达
原位杂交检测caspase-3基因,用探针对孤雌溞和两性溞进行整体原位杂交定位和表达分析。蓝色的深浅表示反义探针在对应部位上阳性信号的强弱,正义探针对照组则没有阳性信号。表达定位结果显示caspase-3基因在蚤状溞活体细胞几乎均有表达,但在孤雌成溞中表达量较低,在两性溞中表达量较高。定位结果显示除肠道和卵鞍外caspase-3 mRNA在溞体全身基本都有表达,广泛定位在胸肢、第一触角、第二触角和性腺等部位(图9)。
3 讨论
3.1 温度对蚤状溞caspase-3 mRNA表达和酶活的影响
对于温度对枝角类生长发育的影响,已经有了很多相关的研究。其寿命因温度升高而缩短[16]。在正常细胞中Caspase-3活性被严格调节,激活的Caspase-3是较少的,即使在凋亡细胞内也只有一部分Caspase-3蛋白被切割参与凋亡过程,少量的增加即可检测到显著差异[17-18]。
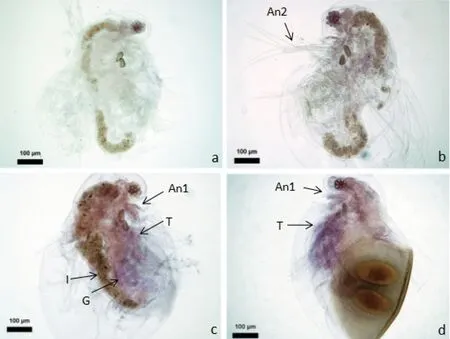
图9 caspase-3基因在蚤状溞中原位杂交位点Figure 9 In situ hybridization site of caspase-3 in D.pulex
注:An1:第一触角;An2:第二触角;T:胸肢;G:性腺;I:肠。 a为空白对照;b为阴性对照,正义探针;c-d为实验组,反义探针(c为孤雌成溞,d为两性雌溞)
Caspase-3在凋亡早期即可检测到,因此半胱天冬蛋白酶-3活性作为细胞凋亡的标记[19]。Caspase-3活性与年龄正相关[20-21],随着年龄的增长,细胞死亡和促进器官凋亡加剧[22]。而Caspase-3酶原的表达量水平高低对衰老进程没有直接影响[23],也就是说Caspase-3低水平活化可使患者更好地康复或存活更久,而与Caspase-3酶原则无关系[24],所以可以认为除了蚤状溞在分化和蜕变时,caspase-3 mRNA的表达量增高表明衰老进程加剧。
在15℃、20℃、25℃和30℃和室温下培养的蚤状溞中,qPCR检测结果显示caspase-3 mRNA表达量与酶活的变化趋势相一致,即随着温度的上升而呈显著的增加(P<0.05)。当温度低的时候,溞体的循环生理和营养代谢水平较低,当温度高的时候则其水平相对会增加,细胞分化、生长、衰老以及凋亡的速率都会增快,从而缩短生殖周期以致衰老加快。但室温处于10℃~18℃,平均温度小于20℃时,其mRNA表达量却比20℃培养下的高,这可能是当温度降低时蚤状溞的生理活动会骤然减慢,从而损伤溞体,以致于加快了细胞的凋亡,caspase-3 mRNA表达量上升。这和贾旭颖等[14]研究淡水养殖的南美白对虾,得出其肝胰腺caspase-3活性在低温突变后显著升高的结论相同。室温组caspase-3 mRNA表达量高于20℃组而Caspase-3酶活与20℃组无显著差异有可能是两组的上限温度接近所致。
3.2 种群密度对蚤状溞caspase-3 mRNA表达和酶活的影响
种群密度对枝角类的生长繁育有重大的影响。一般认为,密度增高使枝角类种群增长速度变慢,冬卵发生率增大,是由于高密度造成代谢产物积累,饵料缺乏所致。陆开宏[25]通过实验发现密度与隆线溞冬卵发生率、产仔率及个体增长速度都有明显相关关系。
在培养密度为50只/500 mL、100只/500 mL、150只/500 mL、200只/500 mL和300只/500 mL蚤状溞中,qPCR检测结果显示caspase-3 mRNA表达量以及酶活随密度上升呈增加趋势,在低培养密度50~100只/500 mL时的表达量无显著差异(P>0.05),之后呈显著上升(P<0.05)。与蚤状溞的种群密度越高,其衰老程度越高的规律相一致[26]。结合蚤状溞的生长情况的观察,密度为50~100 ind/500 mL的蚤状溞在第8天的时候大多已产仔过,甚至有少部分溞怀卵第二胎,而高密度组的蚤状溞大多只是刚开始产第一胎,且密度越高蚤状溞个体越小,所产幼溞也越少。当密度较低的时候,蚤状溞之间的碰撞率低,生存所需要的资源比如食物、氧气等都有富足,所以caspase-3 mRNA的表达无显著变化。当密度过高时,蚤状溞之间需要抢夺生存资源,密度越高每只蚤状溞能分得资源越少,对溞体的损伤越大,细胞凋亡水平升高,caspase-3 mRNA表达水平也会随之升高,部分转入两性生殖,出现卵鞍。
3.3 食物种类对蚤状溞caspase-3 mRNA表达和酶活的影响
食物种类也是影响蚤状溞生长发育的重要因素之一。在之前的研究中,以小球藻为食的蚤状溞生长繁育状况优于以Banta粪土培养液培养的溞[27]。但在本实验中却出现了相反的情况,在qPCR的检验中Banta组caspase-3 mRNA的表达量小于小球藻组溞,意味着小球藻组的溞的衰老情况略高于Banta组溞。酶活测定结果也与此相同。虽然两者之间无显著差异(P>0.05),但在平时的观察中,小球藻组的蚤状溞生长状况是劣于Banta培养液组的,具体表现为溞体较小,怀卵量较少。究其原因,有可能是小球藻组中小球藻的生长状况过于旺盛,以至于蚤状溞对小球藻的消耗量小于小球藻本身的生长速度,导致水体的含氧量、光线等有较大的变化,恶化了蚤状溞的生存环境,也有可能是平时驯化和保种的时候都用的是Banta培养液,可能使蚤状溞更加适应Banta培养液。因条件有限,关于这些推测还有待于以后的研究。
3.4 caspase-3基因在蚤状溞不同生殖状态下的定位表达
虽然目前研究caspase-3基因表达调控机制已有不少报道,但关于caspase-3基因的定位研究的报道仍然较少[28]。本研究用DIG标记的RNA探针检测caspase-3 mRNA在蚤状溞不同生殖状态下的定位表达。整体原位杂交结果显示,caspase-3基因的定位表达广泛,在蚤状溞活体细胞几乎都有表达,在胸肢、第一触角、第二触角上和性腺等部位均有表达,符合Caspase-3在细胞中合成为无活性的酶原,普遍分布在细胞质中[29]。caspase-3基因在两性溞中表达量较高,孤雌成溞中表达量较低。在张萌萌[30]蚤状溞的研究中,caspase-3 mRNA表达量随年龄(1、10、15、20和25 d)增长呈增加趋势(P<0.05),酶活力测定也显示Caspase-3活性随年龄增长而显著增长(P<0.05),意味着随溞年龄的增长,细胞凋亡加剧。即表明除了各种环境因子刺激外,衰老也是一种凋亡的启动因素。虽然环境因子与蚤状溞的生殖模式转换有密切关系,但在不同的培养环境中,随着溞龄的增长,也会产生两性溞和卵鞍,即在培养环境条件好的情况下随着溞年龄的增长,衰老进程的发展,最后从量变发生了质变,进入两性生殖模式。因此我们推测枝角类进行有性生殖主要是由内部因素决定,环境变化仅在一定程度上起诱导作用。
caspase-3基因为环境敏感型基因,caspase-3基因的表达与环境有关,蚤状溞的衰老和caspase-3 mRNA的表达及Caspase-3的酶活有关,而寿命和两性溞的产生相关,但环境因子如何影响caspase-3基因的表达进而通过衰老相关基因调控枝角类生殖转换还需今后进一步的研究。
4 结论
通过Real-Time PCR检测结果发现,caspase-3 mRNA与Caspase-3酶活性增长规律相一致。蚤状溞caspase-3 mRNA 的表达量和酶活整体随着温度和种群密度的增长而增加。以Banta培养液培养的蚤状溞caspase-3 mRNA表达量无显著差异。原位杂交检测结果显示,caspase-3基因在蚤状溞活体细胞几乎都有表达,且在不同生长环境条件下均有表达,在两性溞中表达量较高,孤雌成溞中表达量较低。caspase-3 mRNA的表达广泛分布在溞的胸肢、第一触角、第二触角和生殖腺等部位上。研究认为高温度和高种群密度能够提高蚤状溞caspase-3 mRNA 的表达量并产生卵鞍。蚤状溞的衰老和caspase-3的表达有关,caspase-3的表达与环境因子有关,但环境因子如何影响caspase-3的表达需今后进一步的研究。该研究可为今后探索衰老相关基因调控蚤状溞生殖转换的分子机制提供一些理论依据和参考。
