15个小苍兰品种的花粉形态研究
2019-02-14丁苏芹史益敏唐东芹
孙 忆 丁苏芹 史益敏 唐东芹
(上海交通大学设计学院园林系,上海 200240)
花粉形态特征是由基因控制并在长期进化过程中不断演化而形成的,包含大量的遗传信息,在进化过程中具有保守性、可靠性和稳定性,对研究植物的起源、系统发育、系统分类、品种演化、亲缘关系等都具有重要意义[1~5]。此外,孢粉学标记分析操作简单,在杂交育种中进行花粉形态观察,可作为分析品种间遗传多样性的指标之一,可见其具有良好的实践指导意义。因其分析方法简便,近几十年来,孢粉学在众多有花植物中得到广泛应用,其中包括不少观赏植物。如通过观察木兰科4属7种植物[6]、木兰科16种含笑属植物[7]、牡丹13个品种[8]、莲31份种质间[9]、微型月季10个品种[10]的花粉形态,均发现不同种质之间具有一定差异,体现了不同分类等级种质间存在不同程度的遗传多样性与遗传分化。在球根花卉方面,对百合的孢粉学研究比较多。如张西丽等[11]对9种百合、吴祝华等[12]对12个百合野生种和6个栽培品种、张彦妮和钱灿[13]对百合属4个野生种和8个栽培品种、顾欣等[14]对中国西部四省15种野生百合等,分别进行了花粉形态观察并分析了种质间的亲缘关系。此外,学者们也先后报道了鸢尾[15~17]、郁金香[18]、石蒜[19]等其它球根花卉的花粉形态特征,并探讨了其系统学意义。
小苍兰(Freesiahybrida)是鸢尾科(Iridaceae)香雪兰属(Freesia)的多年生球茎花卉,现代意义上的小苍兰为其园艺杂交品种统称,品种资源丰富,因其具有丰富花色且香气浓郁的实出特点而深受世人喜爱,是著名的切花之一[20]。然而,迄今为止却没有关于小苍兰的花粉形态方面的报道。本研究选用多个小苍兰栽培品种,通过观察比较其花粉形态,进而利用孢粉学标记探讨小苍兰品种间的亲缘关系,旨在为将来对小苍兰的种质鉴定和利用杂交育种等手段开展种质创新等工作提供科学依据。
1 材料与方法
1.1 试验材料
以15个小苍兰园艺品种(Freesiahybrida)作为试验材料开展研究(表1),包括5个自育品种[21]和10个进口品种(均购自荷兰Van den bos公司,https://www.vandenbos.com/zh-cn),均为单瓣型品种,其中包括白色系品种2个、黄色系品种4个、红—紫红色系品种6个以及紫色系品种3个。所有品种选择健壮种球于2015年10月定植于上海交通大学农业与生物学院工程训练中心标准大棚中,进行常规水肥管理。2016年2~4月在小苍兰各品种开花期间采集花粉样品进行观测。
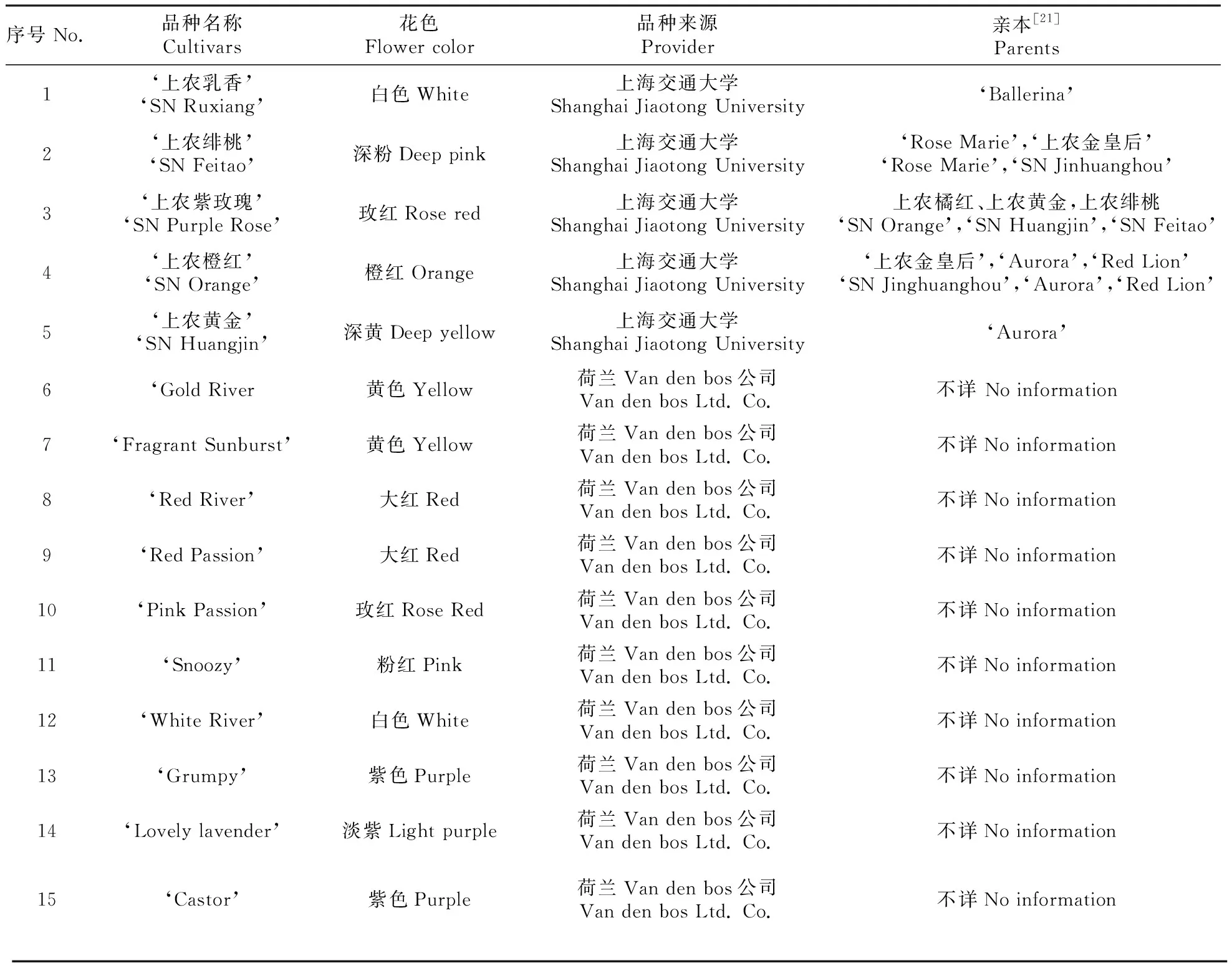
表1 供试小苍兰品种一览表
1.2 试验方法
天气晴朗的上午选择生长健壮,无病虫害的植株采集花粉,装入干燥器中并置4℃条件保存3d。将制备好的样品轻弹于粘有导电胶的样品台上,用离子溅射仪(E-1045)镀金属膜,在扫描电子显微镜(TESCAN-MIRA/WITEC)下测量花粉的极轴(P)、赤道轴(E)的长度并观察记录花粉的形状、外壁纹饰等。
扫描电镜的试验样品制备与观测照相在上海交通大学分析测试中心完成。
1.3 数据统计方法
每个品种观测10~15粒发育良好的花粉并选择有代表性的视野拍照,P、E值测量后取平均值,花粉大小以P×E值表示,花粉形状以P/E表示,其中P/E≥2为超长球形,1.14≤P/E<2为长球形[22]。针对所有定量数据根据标准差和平均值计算其变异系数CV(%)。
利用SPSS软件对花粉的外壁纹饰类型等定性数据以及极轴长(P)、赤道轴长(E)的平均值、P/E值等定量数据进行标准化转换后,定性数据采用二元数据(有记为1,无或极少记为0)表示,采用组间联接结合欧氏距离法(平方Euclidean)进行聚类分析。
2 结果与分析
2.1 小苍兰花粉的外部形态及外壁纹饰特征
用扫描电镜对15个小苍兰品种的花粉形态进行了观察。从外形上看,小苍兰的花粉整体形状为椭球体,所有品种花粉均呈单粒存在且两侧对称,极面观为舟形或心形,赤面观为超长球形或长球形(图1)。花粉具萌发孔,且所有品种均为长达两极的单沟,但不同品种萌发孔的开合状态有所不同,萌发孔一开始呈窄长线形,随着花粉成熟程度,逐渐由2头向中间加宽,有些品种沟内有不规则隆起,并且部分品种孔沟内可见小颗粒状突起(图2)。从花粉的外壁纹饰来看,所有品种均有小刺状突起,并且绝大多数具有穿孔和圆形斑纹,仅上农橙红和Red Passion没有明显穿孔,而Red River和Red Passion则没有圆形斑纹(图2)。

图1 小苍兰花粉外形 A.花粉极面观;B.花粉赤道面观;C.萌发孔Fig.1 Pollen shape of Freesia A.Aperture in pollen;B.Pollen shape in equatorial view;C.Pollen exine ornamentation



图2 小苍兰15个品种的花粉形态及表面纹饰 A.花粉萌发孔;B.花粉赤道面观;C.花粉外壁纹饰特征 1.上农乳香;2.上农绯桃;3.上农紫玫瑰;4.上农橙红;5.上农黄金;6.Gold River;7.Fragrant Sunburst;8.Red River;9.Red Passion;10.Pink Passion;11.Snoozy;12.White River;13.Grumpy;14.Lovely Lavender;15.CastorFig.2 Pollen morphology and exine ornamentation of 15 Freesia cultivars A.Aperture in pollen;B.Pollen shape in equatorial view;C.Pollen exine ornamentation 1.SN Ruxiang;2.SN Feitao;3.SN Purple Rose;4.SN Orange;5.SN Huangjin;6.Gold River;7.Fragrant Sunburst;8.Red River;9.Red Passion;10.Pink Passion;11.Snoozy;12.White River;13.Grumpy;14.Lovely Lavender;15.Castor
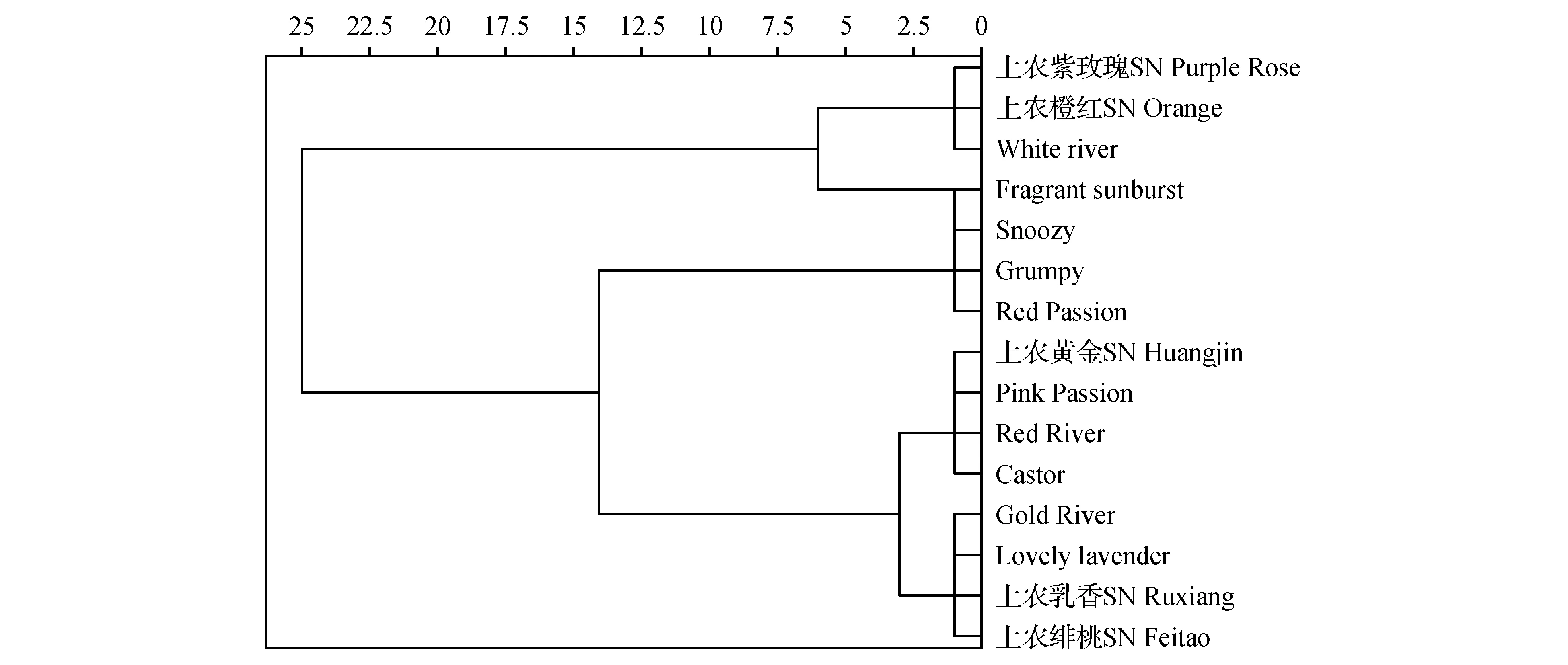
图3 小苍兰15个品种的花粉形态聚类图Fig.3 Pollen morphology clustering map of 15 Freesia cultivars
测定发现,15个小苍兰品种的极轴长(P值)为90~112.6 μm不等,变异系数为6.37%,其中只有4个品种的P值低于100.0 μm;赤道轴长(E值)变化范围为37.0~47.7 μm,变异系数为6.27%,Grumpy最短且为唯一一个E值低于40.0 μm的品种(表2)。进而计算P/E值(表2),按文献划分标准,发现所观测小苍兰的花粉外形绝大部分为超长球形(P/E≥2),仅有Castor一个品种为长球形(P/E为1.96),其变异系数为8.65%。比较15个小苍兰的花粉大小(P×E)发现,Grumpy、Snoozy、Red Passion体积较小(<4 000 μm2),而Fragrant sunburst、上农紫玫瑰、上农橙红的体积较大(>4 800.00 μm2),其中Grumpy最小(3 761.10 μm2),而Fragrant sunburst最大(5 130.59 μm2),两者相差约36%,从该指标的变异系数来看,是所有数量指标中变化最大的,达到9.15%。
2.2 花粉形态聚类分析
根据以上观测的花粉形态特征进行聚类分析,结果发现,在遗传距离5以上的水平,可以将15个小苍兰品种分为5个类群,除Fragrant Sunburst单独成一个类群外,每个类群均含3~4个品种(图3)。
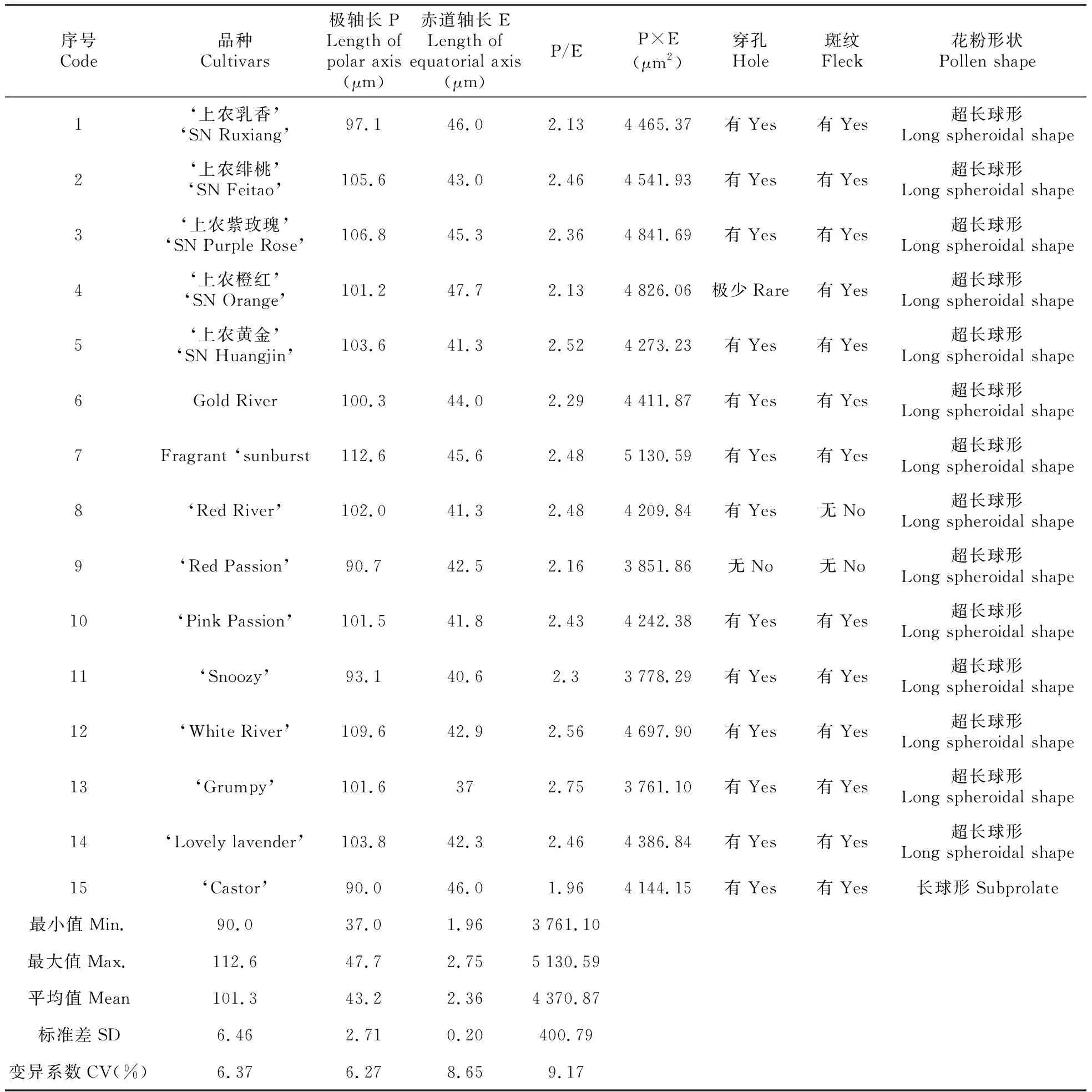
表2 小苍兰的花粉形态特征
第一类群包括上农紫玫瑰、上农橙红、White River和Fragrant Sunburst共4个品种。花粉形状均为超长球形,花粉表面均有圆形斑纹,此外,除上农橙红观察到极少穿孔外,其它品种均有明显穿孔。3个品种的极轴长在101.2~112.6 μm,赤道轴长在42.9~47.7 μm,P/E在2.13~2.56,花粉大小范围为4 697.9~5 130.59,是所有品种中花粉第二大的一个类群。
第二类群包括Snoozy、Grumpy、Red Passion共3个品种。花粉形状均为超长球形,Snoozy和Grumpy的花粉表面明显穿孔和圆形斑纹,而Red Passion则两者饰纹均无。极轴长在90.7~101.6 μm,赤道轴长在37.0~42.5 μm,P/E在2.16~2.75,花粉大小范围为3 761.1~3 851.86,是所有品种中花粉最小的一个类群。
第三类群包括上农黄金、Pink Passion、Red River、Castor共4个品种,前3个品种的花粉形状均为超长球形,而Castor为长球形。花粉表面均有穿孔,此外,除Red River没有观察到圆形斑纹外,其它品种均有明显圆形斑纹。该类群的极轴长在90~103.6 μm,赤道轴长在41.3~46 μm,P/E在1.96~2.52,花粉大小范围为4 144.15~4 273.23。
第四类群包括Gold River、Lovely Lavender、上农乳香、上农绯桃等4个品种,花粉形状均为超长球形,花粉大小居于前2个类群之间而成为第3大的类群。同时,它们的花粉表面均有明显穿孔和圆形斑纹。4个品种的极轴长在97.1~105.6 μm,赤道轴长在42.3~46.0 μm,P/E在2.13~2.46,花粉大小范围为4 386.84~4 541.93。
3 讨论
花粉形态受到基因控制,遗传性状基本稳定且受外界环境条件的影响很小,故孢粉分析已经应用于许多植物的分类和品种亲缘关系分析[23]。作为较高分类依据的科与亚科之间,花粉形态微观特征差异较大,同科不同属之间差异相对较小,而品种间的差异更小,共同点多。简单萌发孔在演化水平上是原始的,复合萌发孔在演化水平上是进化的[24]。本研究首次观测了小苍兰的花粉形态,发现所有品种均具有远极单沟且属于简单萌发孔,与同属于鸢尾科的鸢尾属植物[15~17]以及其它单子叶球根花卉如石蒜、百合、郁金香等的萌发孔相似[13,18~19],同时,与在分类地位上属于较原始的被子植物木兰科植物[6~7]也是一致的。而如牡丹、莲、蔷薇属等萌发孔则表现为更复杂的类型,分别为三拟孔沟、三沟花粉兼具部分散沟花粉和三孔沟[8~9,25],这些植物应属于更进化的类型。因此从花粉萌发孔的演化角度来看,包括小苍兰在内的鸢尾科植物在植物类群中应该属于较原始的类群。此外,根据Erdtman[26]对花粉大小的划分标准(大粒花粉的极轴平均长度在50 μm以上,中粒花粉在39.0~49.9 μm),而大粒的花粉属于原始的类型。因此,小苍兰属于典型的大粒花粉,在进化上应属于较原始的类群。
花粉由无结构层向穿孔演化发展,再继续演化成条纹状类型,是孢粉学家达成共识的植物花粉表面纹饰进化规律[27~28]。花粉外壁纹饰的差异可以从条嵴的有无、宽窄以及深浅等指标反映出来,而且也可衡量被子植物进化程度[22]。观察发现,小苍兰花粉均呈单粒存在,因此属于比较原始的类群,但有趣的是,在Pink Passion这个品种中发现有部分花粉存在条纹化、网状化的趋势,说明该品种相对其它品种而言比较进化。而观测小苍兰花粉外壁纹饰发现,上农橙红和Red Passion没有明显穿孔,可以推测这两个品种不如其它13个品种进化。从文献来看,小苍兰与同科的鸢尾属植物相比较,两者不仅在花粉外形存在差异,更在表面纹饰上存在明显差异,鸢尾属植物多数花粉呈现为扁球形并且有网状纹饰[15~16,29],小苍兰花粉的E值及P/E值明显大于鸢尾属植物。因此,从这个角度来看,鸢尾属植物可能比香雪兰属植物更为进化。
前人研究认为,越进化的花粉其调节功能越强,并且认为花粉进化程度与P/E值存在对应关系,认为(超)长球形(高P/E值)花粉较扁球形(低P/E值)进化[30]。本试验中的15个小苍兰品种花粉粒有超长球形与长球形2种类型,极轴长/赤道轴长比值(P/E)变化幅度为1.96~2.75,从这点上看,小苍兰总体上似乎又属于较进化的类群,这个结论与上述得出小苍兰属于较原始类群正好相反。可见,利用花粉特征分析植物的进化关系需要综合多方面指标来考虑。
当前市场上以切花四倍体小苍兰品种居多[31],但也不乏一些二倍、三倍体品系混淆其中,这导致了现有品种遗传背景变得更为复杂,给小苍兰种质资源的分类和遗传关系的研究造成了更多困难。有研究表明,植物的倍性与花粉大小等特征存在一定关系。如庄东红等[32]研究发现,木槿气孔器的长度、宽度和花粉粒的大小与其染色体基数和倍性呈显著正相关;王振江等[33]观察广东桑的花粉形态,也认为花粉大小及结构特征可作为其倍性鉴定的重要标准。本文中测定小苍兰品种花粉粒大小后发现,其范围为3 761.10~5 130.59 μm2,总体差异较大,是所有观测数量指标中变异系数最大的一个指标,可见现代小苍兰品种存在较丰富的遗传多样性。进而利用花粉特征进行聚类分析发现,花粉大小在划分大类群时具有明显的作用,有一定程度上意味着花粉大小可能与品种亲缘关系存在较为密切的关系。因此,如若花粉粒大小与倍性成正比关系,我们可以在不便计数染色体时,根据成熟花粉粒的大小来估计植物的倍性水平,从而为种质资源的分类和研究提供依据。
上述研究表明,花粉形态在小苍兰不同品种间存在较多的共性,但不同品种间花粉形态特征也具有一定差异,尤其是外壁纹饰细部特征和萌发器官的差别,体现了不同小苍兰品种间存在一定的遗传多样性和遗传分化,可为品种间亲缘关系及种质鉴定等提供有价值的依据。根据花粉特征推测其进化关系,总体上来看,小苍兰属于较原始的类群。然而,单凭孢粉学的证据推测其进化关系、判定品种间亲缘关系可能会不够全面,要进行更全面的分析只有同时结合形态学、分子生物学、细胞学、遗传学及生物化学等多方面的结果,方可得出相对客观的结论。
