小鼠Na+-K+-2Cl-共转运蛋白基因启动子克隆及20-HETE对其转录活性的影响
2019-02-13武晶晶孔令慧贾茹
武晶晶,孔令慧,贾茹
(中国医科大学生命科学学院 1. 医学遗传学教研室; 2. 生物科学系,沈阳 110122)
Na+-K+-2Cl-共转运蛋白(Na+-K+-2Cl-cotransporter,Nkcc2)基因编码的呋塞米敏感的Na+-K+-2Cl-共转运蛋白是溶质转运体超家族成员之一,因此编码该蛋白的基因也被称为SLC12A1[1-2]。NKCC2蛋白主要分布在肾脏髓袢升支粗段,可将2个Cl-、1个Na+和1个K+同向转运到细胞内,参与肾脏近30% Na+的重吸收[3],是重要的高血压候选基因之一。
细胞色素P450 4F2编码的ω-羟化酶,可将花生四烯酸代谢生成二十羟基二十炭四烯酸 (20-hydroxyeicosatetraenoic acid,20-HETE)[4],是重要的血压调节因子。前期本课题组成功地构建了CYP4F2转基因小鼠,并证明转基因小鼠肾脏CYP4F2蛋白表达增加,20-HETE合成增多,是研究20-HETE对血压调控机制的重要动物模型[5]。在对转基因小鼠的研究[6]中发现,转基因小鼠肾脏Nkcc2蛋白和mRNA表达均显著低于野生型小鼠,推测20-HETE在基因转录水平对Nkcc2存在调控作用。本研究通过构建小鼠Nkcc2基因启动子萤光素酶报告基因载体,利用20-HETE刺激转染了Nkcc2启动子报告基因载体的HEK-293T细胞,检测该报告基因的转录活性,初步分析20-HETE对Nkcc2基因启动子活性的影响,为阐明20-HETE对Nkcc2基因调节作用的分子机制奠定实验基础。
1 材料与方法
1.1 材料与试剂
大肠杆菌JM109感受态细胞、pMD18-T Vector、Taq DNA polymerase、LA Taq DNA polymerase、DNA marker、T4 DNA连接酶、KpnⅠ和Hind Ⅲ限制性内切酶、琼脂糖均购自日本TaKaRa公司,DMEM高糖培养基、胎牛血清购自美国HyClone公司,人类胚胎肾细胞系HEK293T细胞购自中国科学院上海细胞生物学研究所,胰蛋白酶购自美国Biosharp公司,萤光素酶报告基因载体pGL3-Basic、海参萤光素酶载体pRL-TK、Dual-GloTM双萤光素酶检测系统购自美国Promega公司,胰蛋白胨、酵母提取物、琼脂粉购自美国Sigma-Aldrich公司,胶回收、质粒制备试剂盒购自美国Axygen公司,脂质体LipofectamineTM2000购自美国Invitrogen公司,20-HETE购自美国Cayman公司,引物合成由中国南京金斯瑞生物科技有限公司完成,DNA测序由中国沈阳优唯生物科技有限公司完成。
1.2 方法
1.2.1 生物信息学分析:应用生物信息学软件ALi-Baba 2.1和TRANSFAC TESS,对小鼠Nkcc2基因启动子区-1 474 bp~+1 bp序列进行预测,比对2个软件的分析结果,寻找潜在的顺式作用元件。
1.2.2 小鼠Nkcc2基因启动子的克隆:
1.2.2.1 小鼠基因组DNA提取与引物设计 取FVB/N小鼠鼠尾,常规方法提取小鼠基因组DNA,-20℃保存。根据Gen Bank数据库获得小鼠Nkcc2基因启动子的DNA序列,利用primer 5.0设计一对特异性引物P1和P2,PCR产物的长度为1 502 bp (-1 462 bp ~+40 bp) 。设计引物:上游引物5’-GTCACCAATTTACCTTT CCCTT-3’;下游引物5’-ACTTACCGCCCCATCCAG-3’。
1.2.2.2 启动子片段扩增及TA克隆 以小鼠基因组DNA为模板进行PCR扩增。PCR产物经电泳鉴定,并利用胶回收纯化试剂盒回收纯化,纯化后的PCR产物连接到pMD-18T载体,16 ℃连接过夜。连接产物转化JM109型感受态细胞,涂布于LB琼脂板,进行蓝白斑筛选。挑取白色克隆于含有氨苄西林的LB培养液中扩增,经菌液PCR鉴定DNA片段插入方向,质粒提取试剂盒提取质粒,经DNA双向测序验证,正确的重组载体命名为pMD18-Nkcc2。
1.2.2.3 小鼠Nkcc2启动子萤光素酶报告基因载体的构建 pMD18-Nkcc2重组载体经KpnⅠ和Hind Ⅲ双酶切,酶切产物经电泳鉴定,并利用胶回收纯化试剂盒回收纯化后,插入到pGL3-Basic载体,16 ℃连接过夜,转化JM109型感受态细胞,挑取单克隆菌落于含有Amp的LB培养液中扩增,试剂盒提取质粒经双酶切及双向测序鉴定,正确的重组载体命名为pGL3-Nkcc2。
1.2.3 小鼠Nkcc2基因启动子活性检测:
1.2.3.1 细胞培养及处理 人类胚胎肾细胞系HEK293T细胞常规培养于含10%胎牛血清,100 U /mL青霉素,100 U /mL链霉素的DMEM高糖培养基中,37 ℃、5% CO2及饱和湿度的条件下培养,用含0.1%胰蛋白酶的消化液消化传代。
1.2.3.2 转染 转染前1 d,用无抗生素培养基接种HEK293T细胞于24孔培养板中,接种密度以24 h后细胞增长至50%~70%为宜,分别用无血清无抗生素的培养基配置质粒DNA (小鼠Nkcc2启动子萤光素酶报告基因质粒及海参萤光素酶对照质粒) 和脂质体LipofectamineTM2000,两者混合后室温孵育20 min,加入24孔培养板培养基中转染HEK293T细胞。细胞转染24 h后,用终浓度为1 μ mol/L 20-HETE刺激2 h。
1.2.3.3 Dual-GloTM双萤光素酶检测系统检测荧光素酶活性 依据试剂盒说明书PBS洗涤细胞后,加入100 μ L 1×PLB裂解细胞,室温震荡15 min,将10 μ L细胞裂解液移入萤光素酶检测管中,每管加入35 μ L LAR Ⅱ,利用生物发光仪检测萤火虫萤光强度值,每管再加入35 μ L Stop-reagent,检测海参萤光强度值,计算相对荧光值。
1.3 统计学分析
实验数据均由3次独立实验结果的表示,采用SPSS 17.0软件进行统计学分析,P< 0.05为差异有统计学意义。
2 结果
2.1 小鼠Nkcc2基因启动子区的生物信息学分析
登录美国国家生物技术信息中心Gen Bank下载小鼠Nkcc2基因启动子区1 474 bp序列,应用生物信息学软件AliBaba2.1和TRANSFAC TESS分析小鼠Nkcc2基因启动子区结构 (-1 474 bp~+1 bp) ,该区域存在多个顺式作用元件,其中NF-κ B、AP-1等均与20-HETE调控靶基因转录活性相关,是20-HETE调控Nkcc2基因的潜在作用元件,见图1。

图1 小鼠Nkcc2基因启动子区潜在转录因子结合位点Fig.1 Transcription factor binding site of the murine Nkcc2 gene promoter region
2.2 Nkcc2基因启动子萤光素酶报告基因载体的构建
以小鼠基因组DNA为模板,利用PCR扩增小鼠Nkcc2基因启动子片段 (-1 462 bp ~ +40 bp) ,PCR产物经琼脂糖凝胶电泳检测,片段大小与预期相符(图2) 。将PCR产物克隆至中间载体pMD18-T vector中,经PCR、双向测序及BLAST比较分析,鉴定出与GenBank序列一致的pMD18-Nkcc2重组载体,经限制性内切酶KpnⅠ和HindⅢ双酶切,回收目的短片段,插入到经KpnⅠ和HindⅢ双酶切的无启动子萤光素酶报告基因载体pGL3-Basic上。重组载体pGL3-Nkcc2经Kpn I和Hind Ⅲ双酶切,琼脂糖凝胶电泳分析,可见1 502 bp的阳性插入片段 (图3) ,且测序结果 (图4) 与GenBank序列一致,证明用于分析Nkcc2启动子活性的萤光素酶报告基因载体构建成功。
2.3 20-HETE对Nkcc2基因启动子活性的影响

图2 小鼠Nkcc2基因启动子PCR产物电泳图Fig.2 Agarose gel electrophoresis of the PCR-amplified murine Nkcc2 gene promoter fragmen
为验证20-HETE对Nkcc2基因启动子转录活性的调控作用,将pGL3-Nkcc2萤光素酶报告基因载体瞬时转染到HEK293T细胞24 h后,用1 μ mol/L 20-HETE刺激2 h,并利用萤光素酶报告基因系统检测Nkcc2基因启动子转录活性。与对照组相比,20-HETE处理组Nkcc2基因启动子转录活性下降了27.5% (P< 0.05) ,说明20-HETE可有效下调Nkcc2基因启动子转录活性,初步验证了假设。

图3 重组质粒双酶切鉴定电泳图Fig.3 Agarose gel electrophoresis of the recombinant plasmid digested by restriction endonuclease
3 讨论
原发性高血压是当今医学领域最具挑战性的心血管疾病之一,也是全球范围内重大公共问题。血压调节是一个多系统多环节的复杂过程,其中肾小管Na+重吸收作用通过调节机体水钠代谢,参与血压调节。Nkcc2基因编码的Na+-K+-2Cl-共转运体主要分布在肾脏髓袢升支粗段,参与肾小管Na+的重吸收,因此该基因是重要的高血压候选基因。
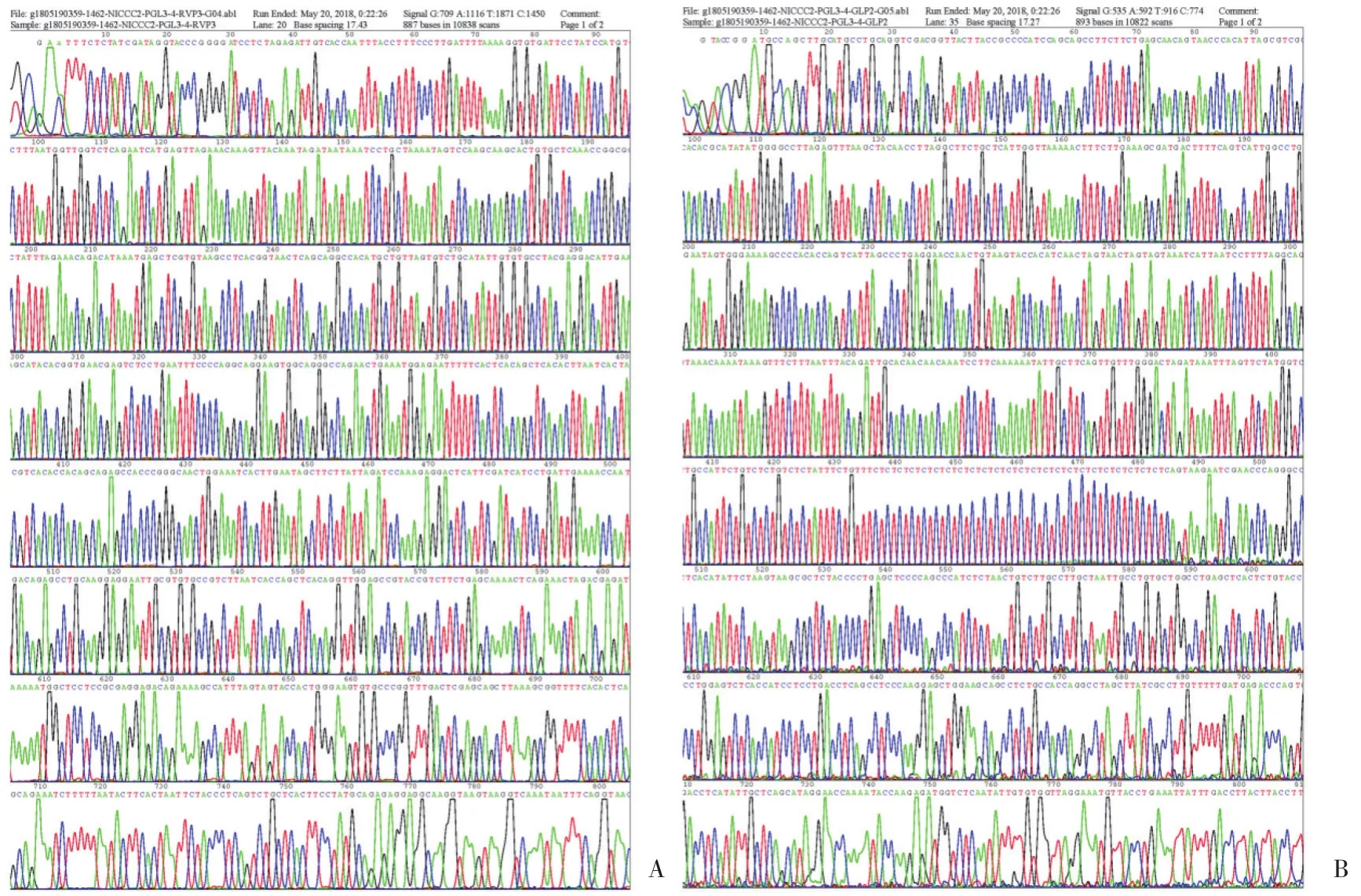
图4 重组质粒双向测序结果Fig.4 Bidirectional sequencing of the recombinant plasmid
CYP4F2是另一个重要的高血压候选基因,该基因编码的ω-羟化酶是人类肾脏产生20-HETE最重要的酶之一。20-HETE主要由髓袢升支粗段生成,通过抑制Nkcc2发挥利尿钠作用,然而其具体机制尚不清楚。有报道[7-8]称20-HETE通过抑制ROMK活性,减少Nkcc2转运所需K+浓度差的势能,间接抑制Nkcc2的活性。本课题组前期利用4% NaCl的高盐饲料喂养具有高20-HETE水平的CYP4F2转基因小鼠,发现20-HETE与高盐有协同作用,通过经典的泛素-蛋白酶体系统降解肾脏Nkcc2蛋白,证明了20-HETE通过翻译后调控机制参与肾脏Nkcc2蛋白表达的调控。本研究中还发现,正常饮食CYP4F2转基因小鼠肾脏Nkcc2蛋白和mRNA表达均低于同组野生型小鼠,推测20-HETE在基因转录水平对Nkcc2表达亦存在调节作用。虽然,目前尚未有直接证据证明20-HETE对Nkcc2基因表达存在调控作用,但仍有一些报道从侧面支持本研究的结论。在对盐敏感性高血压动物模型Dahl盐敏感性大鼠的研究[9-12]中发现,Dahl盐敏感性大鼠内源性20-HETE水平较低是引起盐敏感性高血压的主要原因;对Dahl盐敏感性大鼠肾脏水钠代谢相关通道蛋白表达谱的研究[13]发现,Dahl盐敏感性大鼠肾脏Nkcc2蛋白表达及活性均高于其他种类大鼠;而利用Clofibrate外源性诱导20-HETE合成增加可恢复Dahl盐敏感性大鼠肾脏Nkcc2至正常水平[9-10],以上结论均间接说明了20-HETE对Nkcc2的表达存在调控作用。
结合本课题组的前期工作基础和其他实验室关于20-HETE对Nkcc2作用机制的研究,推测20-HETE在转录水平存在对Nkcc2基因表达的调节作用。为探讨20-HETE对Nkcc2基因转录调控的作用机制,利用生物信息学软件分析小鼠Nkcc2基因启动子区-1 474 bp ~+1 bp序列,发现该区域存在多个20-HETE调控基因转录的潜在作用元件,如NF-κ B、AP-1等。进一步构建了小鼠Nkcc2基因启动子区萤光素酶报告基因载体pGL3-Nkcc2。20-HETE刺激转染了pGL3-Nkcc2载体的HEK293T细胞后,通过萤光素酶报告基因系统检测Nkcc2基因启动子转录活性,结果显示,20-HETE能有效减少pGL3-Nkcc2重组载体转录活性,初步判断20-HETE在转录水平下调Nkcc2基因的表达。
综上所述,本研究成功构建了小鼠Nkcc2基因启动子区重组报告基因载体pGL3-Nkcc2,为进一步探究20-HETE对Nkcc2基因的调控机制提供了有利的研究工具。同时,本研究发现20-HETE对pGL3-Nkcc2重组载体转录活性存在抑制作用,为进一步鉴定20-HETE对Nkcc2基因表达调控的分子机制奠定了实验基础。
