Basics on the use of acid-sensing ion channels'inhibitors as therapeutics
2019-02-13JamilehDibasHoussamAlSaadAdnanDibas
Jamileh Dibas, Houssam Al-Saad, Adnan Dibas,
1 Faculty of Pharmacy, Applied University, Amman, Jordan
2 North Texas Eye Research Institute, Department of Pharmacology and Neuroscience, University of North Texas Health Science Center at Fort Worth, Fort Worth, TX, USA
Abstract Since the discovery of acid-sensing ion channels in 1997, their importance in the health of neurons and other non-neuronal cells has gained significant importance. Acid-sensing ion channels play important roles in mediating pain sensation during diseases such as stroke, inflammation, arthritis, cancer, and recently migraine. More interestingly, acid-sensing ion channels may explain the sex differences in pain between males and females. Also, the ability of acid-sensing ion channel blockers to exert neuroprotective effects in a number of neurodegenerative diseases has added a new dimension to their therapeutic value. The current failure rate of ~45% of new drugs (due to toxicity issues) and saving of up to 7 years in the life span of drug approval makes drug repurposing a high priority. If acid-sensing ion channels' blockers undergo what is known as “drug repurposing”, there is a great potential to bring them as medications with known safety profiles to new patient populations. However, the route of administration remains a big challenge due to their poor penetration of the blood brain and retinal barriers. In this review, the promise of using acid-sensing ion channel blockers as neuroprotective drugs is discussed.
Key Words: optic nerve; glaucoma; neurodegeneration; neuroprotection; acid; sensing; ion channel; calpain
Introduction
Under normal physiological conditions, extracellular pH (pHo)is maintained around ~7.3 while intracellular pH (pHi) is ~7.However, under pathological conditions such as an ischemic stroke, seizure, and inflammation, pH is greatly shifted towards acidification, a mechanism mediated in part by anaerobic glycolysis and the accumulation of lactic acid and protons. Neurons are mostly at risk of uncontrolled acidification leading to their death and permanent neurological impairment. As a defensive mechanism, cells including neurons express a family of proteins known as the acid-sensing ion channels (ASICs) that are sensitive to changes in extracellular pH reduction.
ASICs, a family of cation-selective, voltage insensitive, and proton-sensing channels belongs to a proton-gated family of the degenerin-epithelial channel family of cation channels,which are responsible for Na+influx (Waldmann et al., 1997).In the human genome, there are four ASIC genes that have been assigned the symbols of Accn1 (ASIC2), Accn2 (ASIC1),Accn3 (ASIC3), and Accn4 (ASIC4) (Lin et al., 2015). A novel Accn5 encodes a related channel, BLNaC/BASIC that is however different from ASICs (Lin et al., 2015). ASIC1 is the most abundant ASIC subunit in the mammalian central nervous system (Chen et al., 1998). Similar to ASIC1a, ASIC3 has equal sensitivity to H+however; it is mainly expressed in peripheral nervous system (Waldmann et al., 1997).
ASICs form trimeric ion channels composed of either identical (homotrimeric) or different (heterotrimeric) subunits.ASICs are permeable to monovalent cations (Na+> K+) and protons (Waldmann et al., 1997), and some ASICs (ASIC1a homomeric channels and ASIC1a/2b heteromeric channels) are also permeable to divalent cations such as calcium, suggesting that they could play a particularly important role in intracellular signaling as well as membrane excitability (Stankowska et al., 2018). Interestingly, while both ASIC1a and ASIC3 are active at neutral pH (7.1-7.3), ASIC2a is only active at low pH(~4.5) (Hesselager et al., 2004).
ASIC1 Blockers are Potential Therapeutics
ASICs have been associated with many neurodegenerative disorders and recent observations suggest that ASICs are major contributors to axonopathy in such pathologies. The inhibition of ASICs is neuroprotective in stroke, Huntington's disease(Walker, 2007), multiple sclerosis (Vergo et al., 2011), and Parkinson's disease (Arias et al., 2008). List of diseases where ASIC blockers exerted neuroprotective effects is shown in Figure 1A.Using optic nerve crush in rats (Stankowska et al., 2018) and ischemia/reperfusion (retinal stroke model) in mice (Dibas et al., 2018), we demonstrated that blockers of ASIC1 such as amiloride (nonselective) and psalmotoxin-1 (selective ASIC1a blocker), exerted neuroprotective effects on retinal ganglion cells. In both models, an upregulation of ASIC1 and ASIC2 was observed (ASIC1 significantly increased in optic nerve extracts and ASIC1 and ASIC2 increased in retinal ganglion cells in ischemia/reperfusion model). Not surprisingly, acidification increased intracellular Ca2+([Ca2+]i) in isolated primary rat retinal ganglion cells, an effect attenuated by psalmotoxin-1(Stankowska et al., 2018). We have shown and others that ASIC upregulation and activation can induce extracellular Ca2+influx and increase in intracellular Ca2+(Stankowska et al.,2018). ASIC1a-inducd influx of extracellular Ca2+mediating ischemic neuronal death, can be prevented by either reducing extracellular Ca2+or ASIC1a inhibitors (psalmotoxin-1 and amiloride) (Xiong et al., 2004; Yermolaieva et al., 2004; Gao et al., 2005). Elevated intracellular Ca2+is a known activator of the calcium-dependent calpains. Activation of calpain 1 via autolysis plays a key role in mediating neuronal apoptosis as it cleaves essential proteins in neurons such as fodrins. Myelin basic proteins and axonal neurofilament protein are also known substrates for calpain and their loss greatly affects structure of neurons (Li and Banik, 1995; Das et al., 2013). Calpains are involved in retinal neurodegeneration in a number of injury models (Wu et al., 2004; Oka et al., 2006; McKernan et al., 2007;Huang et al., 2010).
In optic nerve crush in rats and ischemia/reperfusion (retinal stroke model) in mice, calpain-1 activation was evident as increased fodrin cleavage was detected. Heat shock protein(HSP)70, a known neuroprotective protein and a substrate for calpain, was degraded following retinal ischemic stroke (Dibas et al., 2018). HSP70 induction or overexpression has been shown to be neuroprotective in retinal ganglion cells in an Intraocular pressure-model and during optic nerve crush (Ueda et al., 1998; Park et al., 2001a). We have shown that ischemia/reperfusion induced-upregulation of ASIC1 correlated with degradation of HSP70 and the appearance of a lower band of 55 kDa.While psalmotoxin-1 pretreatment did prevent fodrin cleavage,a well-known calpain substrate, unfortunately it failed to block HSP70-induced cleavage. This inability to affect HSP70 may explain the partial rescue of retinal ganglion cells by psalmotoxin-1. HSP70 protein is considered a master switch in neuroprotection. Its mechanism of protection involves different pathways.One mechanism involves enhancement of autophagy as HSP70 stabilized lysosomes by binding to endolysosomal phospholipid,bis(monoacylglycero)phosphate, and acted as a chaperone recycling damaged proteins to lysosomes for elimination (Kirkegaard et al., 2010). HSP70 also blocked maturation of apoptosome by binding to key elements preventing the recruiting of caspases and their activation (Beere et al., 2000). Furthermore, Matsumori et al. (2005) have shown that HSP70 attenuated caspase-unrelated cellular death pathways as well. Also, HSP70 blocked c-Jun N-terminal kinase-dependent and p38 mitogen-activated protein kinase signaling pathways (Gabai et al., 1997; Park et al., 2001b).HSP70 suffered oxidization by free radicals-induced carbonylation of its Arg469 amino-acid leading to its degradation by calpains. HSP70 depletion causes lysosomal destabilization and autophagy failure that lead ultimately to neurodegeneration(Zhu et al., 2002; Oikawa et al., 2009; Sahara and Yamashima,2010; Furukawa and Koriyama, 2016). Our data is consistent with that reported by Nakajima et al 2006, who have shown that monkey retinas subjected to hypoxia suffered from neuronal degeneration via calpain activation and degradation of fodrin and HSP70 (Nakajima et al., 2006).
Finally, at least three clinical trials (NCT01802489,NCT01879527 and NCT01910259) are currently underway or finished assessing blocking ASICs using amiloride (nonselective blocker) for the treatment of optic neuritis. The results from first clinical trial (NCT01902489) was recently published and although it appears that amiloride did not demonstrate any neuroprotective effects based on the scanning laser polarimetry(which measures the thickness of the peripapilliary retinal fiber layer), visual evoked potentials measures were enhanced in the amiloride group. However, newer trials have been suggested with changed window of opportunities (McKee et al., 2017).The results from NCT01879527 and NCT01910259 clinical trials have not been released yet although both supposedly ended.
The Anti-Nociceptive Effects of ASIC Blockers
Peripheral ASICs are involved in cutaneous pain. Acidification of human skin was accompanied by cutaneous pain (Steen et al., 1995), an effect that was blocked by amiloride, a nonselective ASIC blocker (Ugawa et al., 2002; Jones et al., 2004). Amiloride also exerted analgesic effects following topical application in pain models of inflammatory and postoperative pain in rodents (Kuduk et al., 2009). Interestingly, ASIC inhibition may be helpful for migraine patients as amiloride reduced headache symptoms in a human clinical trial (Holland et al., 2012). Elevated ASIC proteins have been also detected in human degenerated intervertebral discs compared to healthy tissues suggesting a role for ASICs in inflammation and ostheoarthritis-induced joint pain (Cuesta et al., 2014). Not surprisingly, intra-articular administrations of anthopleura elegantissima toxin 2 (APETx2),an ASIC3 blocker, reduced pain-related behavior and inflammation-induced cartilage damage in animals (Izumi et al.,2012). Interestingly, the known non-steroid anti-inflammatory drugs (ibuprofen, aspirin and diclofenac) directly inhibited the activity of both ASIC1a and ASIC3 channels with ASIC2a being partially blocked by diclofenac (0.5 mM) (Voilley et al., 2001).
The Neuroprotective Properties of Psalmotoxin-1 and Related Peptides
Various ASIC inhibitors have also been isolated from venoms.Psalmotoxin-1, a toxin isolated from the venom of tarantula spider, is specific to ASIC1a homomeric and ASIC1a/2b heteromeric channels has exerted neuroprotective effects in several injury models including; ischemic stroke (Xiong et al.,2004), porcine models of cerebral ischemia (Yang et al., 2011),ischemic stroke in conscious hypertensive rats (McCarthy et al., 2015), optic nerve crush in rats (Stankowska et al., 2018),and retinal stroke model in mice (Dibas et al., 2018). Hi1a is a recently discovered ASIC1a blocker with almost double the size of PcTx1 (psalmotoxin-1; 75 amino acids) (Chassagnon et al., 2017), is more potent at inhibiting ASIC1a than PcTx1 with an estimated half maximal inhibitory concentration (IC50) of~400 pM. Chassagnon et al., 2017 have shown that intracerebroventricular injection of Hi1a (2 ng/kg) exerted significant neuroprotective effects as late as 8 hours post-injury. Also, novel peptides known as mambalgins 1, 2, and 3 isolated from the venom of mamba snakes are also potent inhibitors of ASIC1a as homotrimer or heterotrimers. Mambalgin-1 inhibited currents from ASIC1a and ASIC1b homomers (ASIC1a and ASIC1b are isoforms of ASIC1) as well as from heteromers of ASIC1a-ASIC2a, ASIC1a-ASIC2b or ASIC1a-ASIC1b (ASIC2a and ASIC2b are isoforms of ASIC2) (Diochot et al., 2012; Mourier et al., 2016). APETx2, on the other hand, inhibits ASIC3 containing channels (Chagot et al., 2005). However, to date, there are no known selective ASIC2a/2b inhibitors.
Does ASICs' Differential Gene Expression Explain Female Over Sensitivity
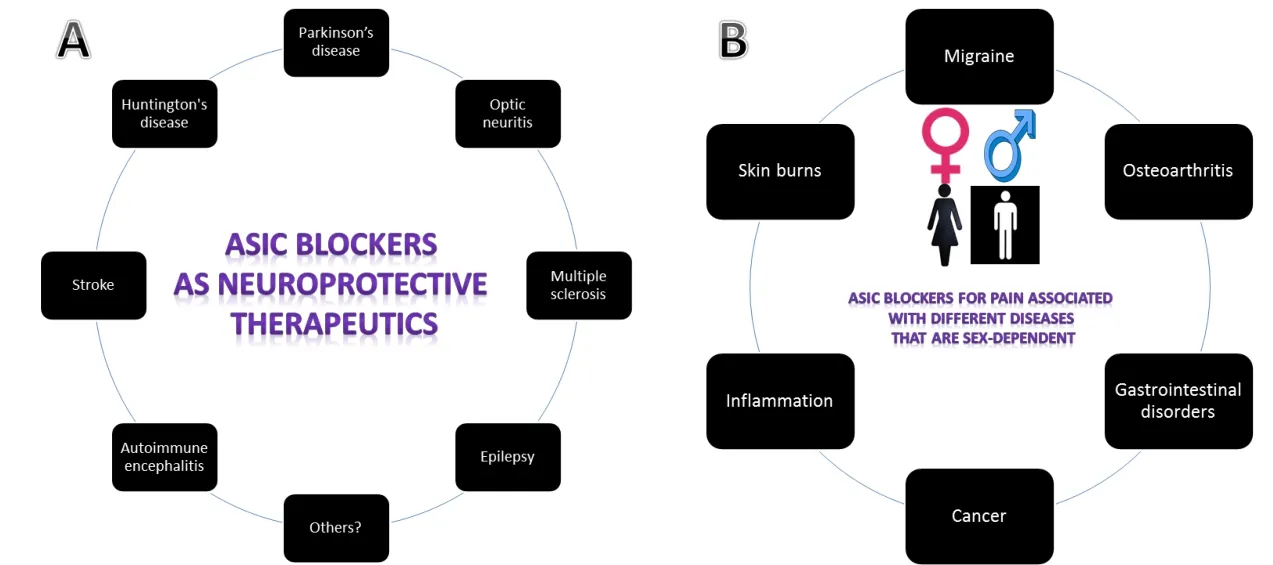
Figure 1 The promising role of ASIC blockers as therapeutics.
Overwhelming evidence is suggesting sex-based disparities between males and females with a clear majority of chronic patients being females (Mogil, 2012). Women with knee osteoarthritis have greater pain than men (Sluka et al., 2012). Women also demonstrate higher pain sensitivity and prevalence of chronic visceral pain conditions such as fibromyalgia, migraine,and functional gastrointestinal disorders (Unruh, 1996). The exact mechanisms for why women constitute a large majority of chronic patients are not known. While psychological, hormonal, and genetic differences have been suggested to explain reasons making women more sensitive to pain than men and why women endure more pain post operationally and experience different outcomes to therapeutical interventions, ASICs's differential expression between both genders may play a role.
Studies have shown that female bladders are more sensitive to acidic injuries than male bladders. ASIC2 (which is only activated at acidic pH of ~4.5), is expressed at almost double the levels in female muscle bladder compared to male mouse bladders (Kobayashi et al., 2009), which could possibly be a contributor to the increased susceptibility in females. Male mice injected with 2-guanidine-4-methylguanidine (an ASIC3 agonist), in their paws licked their 2-guanidine-4-methylguanidine-injected paw ~34% less than female mice suggesting a clear difference in pain sensitivity between genders (Izurieta Munoz et al., 2018).However, there are no current studies on ASICs's expression in different tissues in males and females although this could help explain the substantial sex differences in clinical and experimental pain outcomes for both genders. A list of diseases where ASIC blockers may exert sex-based anti-nociceptive effects is shown in Figure 1B.
Future Directions and Limitations
Although ASICs's inhibitors are known for their pain-attenuation or control as well diuretics (in the case of amiloride), their promising neuroprotective effects exerted by ASIC1a selective blockers are opening a new exciting field in the fight against neurodegenerative diseases. The introduction of ASIC1a targeted agents (e.g., psalmotoxin-1, H1a, and mambalgins) has raised the question whether alternate clinical trial designs and window trials, are better suited to evaluate such new drugs. However,there are limitations due to lack of delivery of such inhibitors without invasive techniques due to the inability of penetrating the blood-brain barrier and the retinal blood barrier. Bloodbrain barrier is impermeable to almost 98% of small molecules(> 600 Daltons) and impermeable to large molecules. Current procedures of delivering drugs to brain such as intracerebroventricular, intracerebral, and intrathecal injections, require specialized experience and not only carry high risk of infections but also cannot be repeatedly used in patients. Similarly, intravitreal injection although successful in delivering drugs to the retina, has the risk of side effects and limitation on the number of times to be used in the human eyes. However, intranasal administration provides a non-invasive route for central nervous system drug delivery that successfully bypassed the blood-brain barrier. There are 102 completed clinical trials that assessed the intranasal administration delivery of numerous drugs with promising results (clinicaltrials.gov). There is a great difference in brain deposits of psalmotoxin-1 when injected intravenously vs. intranasal administration. Following intravenously administration of radiolabeled psalmotoxin-1, data clearly indicated that psalmotoxin-1 is unlikely to cross the blood-brain barrier in any appreciable amount. To the contrary, using intranasal administration of 5 μg psalmotoxin-1, ~1.3 ng (~0.03%) was detected in the olfactory bulb and many brain compartments(Er, 2017). McCarthy et al. (2015) reported that intracerebroventricular administration of psalmotoxin-1 at 1 ng/kg (~25 pg for a 25 g mouse) 2 hours post stroke exerted neuroprotective effects in mice. More interestingly, Pignataro et al. (2007) administered psalmotoxin-1 (intranasal administration) at 25 ng/mouse, also provided neuroprotection when given within the first 4 hours post brain ischemic stroke. Finally, it remains of high importance to evaluate any role for ASICs in gender-based pain sensation and the potential of development of masculinity/feminine-dependent therapy.
Author contributions:Article writing: JD, HAS and AD.
Conflicts of interest:None declared.
Financial support:This work was supported by the BrightFocus Foundation and intramural grant from University of North Texas Health Science Center at Fort Worth (to AD).
Copyright license agreement:The Copyright License Agreement has been signed by all authors before publication.
Plagiarism check: Checked twice by iThenticate.
Peer review: Externally peer reviewed.
Open access statement:This is an open access journal, and articles are distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 License, which allows others to remix, tweak, and build upon the work non-commercially, as long as appropriate credit is given and the new creations are licensed under the identical terms.
Open peer reviewer:Steven Levy, MD Stem Cells, USA.
Additional file:Open peer review report 1.
杂志排行

中国神经再生研究(英文版)的其它文章
- The role of Epstein-Barr virus in multiple sclerosis:from molecular pathophysiology to in vivo imaging
- The metabolome identity: basis for discovery of biomarkers in neurodegeneration
- Neuroinflammation as a target for glaucoma therapy
- Role of axon resealing in retrograde neuronal death and regeneration after spinal cord injury
- Rehabilitation following spinal cord injury: how animal models can help our understanding of exerciseinduced neuroplasticity
- The pig as a preclinical traumatic brain injury model:current models, functional outcome measures, and translational detection strategies