猕猴桃POD基因的克隆和表达分析
2019-02-13陈义挺赖瑞联冯新高敏霞程春振陈文光吴如健
陈义挺, 赖瑞联, 冯新, 高敏霞, 程春振, 陈文光, 吴如健
猕猴桃基因的克隆和表达分析
陈义挺1, 赖瑞联1, 冯新1, 高敏霞1, 程春振2, 陈文光1, 吴如健1*
(1. 福建省农业科学院果树研究所,福州 350013; 2. 福建农林大学园艺植物生物工程研究所,福州 350002)
为了解猕猴桃基因的表达调控功能,采用RT-PCR技术从‘米良1号’猕猴桃(‘Miliang-1’)克隆了2个POD家族成员基因(和)。结果表明,和开放阅读框分别为984和957 bp, 预测分别编码327和318个氨基酸,GenBank登录号分别为MF774100和MF774101。AdPOD27和AdPOD64为亲水性碱性蛋白,属于植物亚铁红素依赖Ⅲ型POD超家族成员,含有信号肽、跨膜螺旋结构和磷酸化位点,亚细胞定位预测分别定位于线粒体和细胞外。qRT-PCR结果表明,在脱落酸(ABA)和4℃处理时表达量急剧上升,而只在ABA处理时表达量显著提高。此外,表达量与POD活性、表达量均存在显著相关性。因此,和可能在猕猴桃果实软化、低温响应和ABA诱导等过程中发挥重要的调控功能。
猕猴桃;过氧化物酶;果实软化;低温;脱落酸
猕猴桃()是猕猴桃科(Actini- diaceae)猕猴桃属落叶藤本果树,果实中富含维生素C、亚麻酸、氨基酸和微量元素等,被认为是20世纪野生果树驯化最成功的果种之一[1]。然而,猕猴桃果实耐贮性差,采后易软化腐烂,严重影响猕猴桃可食性和商品价值,如何延长猕猴桃贮藏保鲜期成为猕猴桃产业发展亟待解决的难题[2]。
过氧化物酶(peroxidase, POD)是由单一肽链与卟啉组成的一种血红素蛋白,能够以铁卟啉为辅基,以H2O2为电子受体催化底物氧化,在植物体内发挥抗氧化作用。POD是多基因家族编码的同工酶,功能广泛且丰富多样,在植物生长、发育和衰老过程中,POD活性水平均有强烈变化[3–4]。此外,POD还参与植物激素调控、活性氧代谢、抗病虫、耐干旱胁迫等生理过程,是植物响应逆境胁迫的关键酶之一。过表达过氧化物酶基因明显提高番茄对苯酚胁迫的耐受性[5];基因能够响应缺硼胁迫改变柑桔砧木中木质素含量[6];POD活性会影响温州蜜柑贮藏过程中果皮和果肉中的枯水发生,进而影响果实采后衰老[7]。
已有研究表明,POD在猕猴桃采后贮藏和保鲜过程中发挥重要作用。Zolfaghari等[8]认为,低温下猕猴桃POD活性均提高,长时间冷藏于(1±1)℃, 相对湿度(80±5)%有利于维持猕猴桃最佳品质。猕猴桃花后不同时间采收的果实中POD活性也存在显著差异[9]。在猕猴桃采后贮藏效果影响因素研究中,一氧化氮、1-甲基环丙烯、氮气、硫化氢和槲皮素等均能上调猕猴桃果肉中POD活性,维持风味品质和较高的抗病能力,减少果实冷藏冷害,延缓果实衰老,但也可能引起部分品种果心木质化[10–17]; 而草酸能够显著抑制猕猴桃果肉中POD等木质素合成相关酶的活性,减轻因木质化引起的果实品质劣变[18]。此外,微波、高压和热处理也会影响猕猴桃果汁和果酱中POD活性,从而改变其贮藏效果[19–20]。
低温贮藏是延长猕猴桃果实货架期的有效途径,而脱落酸(abscisic acid, ABA)在猕猴桃后熟衰老过程中能够促进水解酶活性增强,参与猕猴桃果实成熟过程的软化启动[21]。目前,猕猴桃POD相关研究主要集中在POD活性改变对果实贮藏、衰老和品质等生理方面影响,其分子机理尚不明确。低温和ABA作为目前已知的猕猴桃软化两个重要相关因素,其在软化过程中与POD的相互作用关系及其调控机制也未见报道。而在前期研究中,通过高通量转录组测序发现,和在猕猴桃不同贮藏处理组中存在明显的转录水平差异。鉴于此,试验以‘米良1号’猕猴桃为材料,对猕猴桃基因组中预测的这2个基因家族成员进行验证和生物信息学分析,同时研究了猕猴桃贮藏软化、低温和ABA处理下POD活性和表达模式的相关性,以期为猕猴桃基因的调控功能研究提供理论依据。
1 材料和方法
1.1 材料
试验所用‘米良1号’猕猴桃(‘Miliang-1’)采自福建省农业科学院果树研究所建宁猕猴桃基地。2017年9月份,采集大小相近,成熟度一致,无病虫害和机械损伤的果实,参考陈义挺等[22]的方法进行处理,将猕猴桃随机分为6组(编号1~6),每组10个果实,组1为25℃不贮藏(对照),组2为25℃中贮藏1周,组3~5分别为4℃下贮藏1、2和3周,组6为50 mg L–1的ABA浸泡2 min后于25℃中贮藏1周。每处理重复3次,经液氮速冻后保存于-80℃超低温冰箱中用于POD活性测定和RNA提取。
1.2 POD活性测定
根据猕猴桃基因组数据库对和序列进行验证,分别获得长度为984和957 bp的ORF序列,预测分别编码327和318个氨基酸,起始密码子均为ATG,终止密码子分别为TAA和TAG (图2, 3)。POD27和POD64与‘红阳’猕猴桃(‘Hongyang’)基因组相应CDS序列(GenBank登录号: Achn356831和Achn132211)的相似性分别达到99.39%和99.16%,而和间的核苷酸和预测蛋白序列的相似性分别为49.45%和40.00%。通过NCBI比对,猕猴桃与烟草()和杨树()的相似性分别为74%和73%,与葡萄()和向日葵()的相似性分别为82%和79%,表明所获得的序列为猕猴桃相应基因,分别命名为和,GenBank登录号分别为MF774100和MF774101。
1.3 RNA提取和cDNA逆转录
以猕猴桃为内参基因进行qRT-PCR试验。将各cDNA样品等体积混样,随后按10–1、30–1、90–1和270–1进行梯度稀释用于绘制标准曲线,根据扩增效率筛选适宜的退火温度检测不同样品中基因的相对表达量。采用Eppendorf Realplex4荧光定量PCR仪设置qRT-PCR扩增程序:94℃预变性30 s; 94℃变性10 s,TM退火15 s,72℃延伸10 s,循环40次;94℃ 15 s,60℃ 15 s,再以0.11 ℃ s–1的速度升温至94℃保持15 s,绘制融解曲线。qRT-PCR反应体系为2×TransStart®Top Green qPCR SuperMix 10L,Passive Reference Dye (50×) 0.4L,上下游引物(表1)各0.2mol L–1,模板0.5mol L–1。采用Excel 2003和SPSS 19.0进行数据统计、差异显著性分析和相关性分析。
1.4 基因克隆
与对照相比,25℃和ABA处理的猕猴桃贮藏1周后POD活性显著提高(<0.01),此时猕猴桃硬度急剧下降,可溶性固形物含量上升,果实软化[21],可见POD参与了猕猴桃软化过程,并且POD活性可能受ABA影响(图1)。而低温贮藏过程中,4℃贮藏1周后POD活性明显提高,随后急剧下降,3周后重新上升恢复到对照水平(>0.01),说明猕猴桃POD活性能够响应低温诱导,在猕猴桃抗冷早期发挥重要作用。可见,POD可能在猕猴桃贮藏软化、ABA诱导和低温响应过程中发挥调控作用。
作为一家全球性的集团公司,Walter主要研发、生产和销售用于金属加工的精密刀具。品牌Walter是可转位硬质合金和PCD刀具系统生产商;品牌Walter Titex是高速钢和整体硬质合金钻头及铰刀生产商;品牌Walter Prototyp以高速钢、整体硬质合金螺纹加工刀具和铣刀而闻名;Walter Valenite提供可转位车刀片、钻头、铣刀和高技术含量的MODCO品牌非标刀具。

表1 基因克隆、qRT-PCR引物序列
1.5 生物信息学分析
采用在线软件进行POD生物信息学分析, 基本理化性质(ExPASy,http://web.expasy.org/ protparam/),信号肽(SignalP 4.1 Server,http://www.cbs.dtu.dk/ services/SignalP/),亚细胞定位(PSORT Prediction, https://psort.hgc.jp/form.html), 跨膜结构(TMpred, http://www.ch.embnet.org/software/TMPRED_form.html), 磷酸化位点(NetPhos 3.1 Server, http://www. cbs.dtu.dk/services/NetPhos/), 保守结构域(NCBI conserved domain searc, https://www.ncbi.nlm.nih. gov/Structure/cdd/wrpsb.cgi),结合位点(PROSITE, http://www.expasy.ch/prosite),卷曲螺旋结构(COILS, http://www.ch.embnet.org/software/COILS_form.html),蛋白质二级结构(PSIPRED, http://bioinf.cs.ucl.ac.uk/ psipred/), 蛋白质三维结构(SWISS-MODEL, https:// swissmodel.expasy.org/)。
1.6 qRT-PCR分析
采用植物多糖多酚试剂盒RNAprep pure Plant Kit提取猕猴桃总RNA,经琼脂糖凝胶电泳和紫外光分光光度计进行质量和浓度检测合格后进行等量混合,利用Thermo Scientific RevertAid First Strand cDNA Synthesis Kit逆转录cDNA用于PCR扩增,同时采用 TransScript All-in-One First-Strand cDNA Synthesis SuperMix for qPCR (One-Step gDNA Removel; TransGen Biotech, Beijing)逆转录cDNA用于qRT-PCR试验。
由于互联网发展的历史原因,以传输控制协议(TCP)/网际协议(IP)为核心的互联网模型将“尽力而为”的可达性作为网络的首要任务,这使得报文携带的目的地址在路由过程中成为了唯一的决定因素。这种仅依靠目的地址的路由方式,大大限制了对报文转发控制的灵活性。并且随着互联网规模的迅速增长和用户业务的多样化,传统路由协议越来越难以满足许多业务对服务质量的要求。
2 结果和分析
2.1 POD活性
从已公布的猕猴桃基因组数据库(Kiwifruit Genome Database, http://bioinfo.bti.cornell.edu/cgi-bin/kiwi/home.cgi)中搜索并下载猕猴桃和基因可能序列(Gene ID:Achn356831和Achn132211),设计引物(表1)进行ORF全长扩增。PCR扩增体系为:2×®HiFi PCR SuperMix 12.5L,cDNA模板50 ng,上下游引物各0.1mol L–1,补足高压ddH2O至终体积25L。PCR扩增程序为:95℃预变性5 min,95℃变性30 s,TM退火30 s,72℃延伸1 min,35次循环后再72℃延伸8 min。PCR产物采用1.0%琼脂糖凝胶电泳检测,切割预期目的条带采用®Quick Gel Extraction Kit回收,根据试剂盒说明书采用®-T5 Zero Cloning Kit连接目的片段和载体,转化到-T1感受态细胞中涂板,37℃暗下培养12 h后挑取单克隆子进行PCR检测,选取阳性克隆子送至北京六合华大基因科技有限公司进行测序。
2.2 POD基因克隆
根据POD催化H2O2氧化特定底物,在470 nm有特征光吸收的原理,采用植物过氧化物酶试剂盒(购自苏州科铭生物技术有限公司)测定猕猴桃的POD活性。
2.3 POD生物信息学分析
芬顿氧化技术的影响因素主要为pH值、Fe2+、H2O2投加量及投加方式。普通的芬顿氧化技术中双氧水加药方式为单点投加,而这种加药方式在双氧水投加初期系统会产生大量羟基自由基,过多的羟基自由基不能完全与废水中有机污染物发生反应,导致芬顿试剂产生的部分羟基自由基被无效消耗,最终导致双氧水利用率下降以及降低预处理效果。本文主要研究多点投加芬顿氧化技术最佳反应条件和多点投加方式的优化(包括投加位点及投加量)。
2.4 POD表达模式分析
从图5可见,25℃贮藏1周后猕猴桃的表达量是对照的1.5倍,而的表达量未出现显著变化,说明可能参与猕猴桃果实软化过程,而的调控作用较不明显。ABA处理后,和的表达量急剧上升,分别为对照的419和23倍,说明二者均能够响应ABA诱导,尤其是。猕猴桃果实在4℃处理1周后,的表达量明显上调,为对照的98倍,随后下降,在低温贮藏后期的调控也不明显,而的表达量几乎不发生改变。此外,在所有处理中,的表达量均高于,可见,可能在猕猴桃果实软化、低温响应和ABA诱导等过程中比发挥更全面的调控功能。
生物信息学分析表明,AdPOD27和AdPOD64的基本理化性质存在一定差异(表2),二者均为亲水性碱性蛋白,但AdPOD27属于稳定蛋白,而AdPOD64属于不稳定蛋白;AdPOD27中亮氨酸(Leu)含量最多,达到9.8%,其次是赖氨酸(Lys)和丙氨酸(Ala),分别为8.3%和8.0%,而AdPOD64中丝氨酸(Ser)含量最多,达到11.3%,其次是Leu和Ala,均为10.1%。亚细胞定位表明,AdPOD27和AdPOD64可能分别定位于线粒体和细胞外,二者分别于氨基酸序列5ʹ端第1~26位和第1~24位氨基酸含有信号肽。保守结构域和结合位点预测表明, AdPOD27和AdPOD64蛋白均属于植物亚铁红素依赖Ⅲ型过氧化物酶超家族成员,主要参与细胞间隙和液泡的过氧化氢分解、生长素代谢、木质素合成和胁迫响应等(图2, 3)。TMpred分析表明,AdPOD27含有4个由内向外和由外向内的跨膜螺旋结构以及28个磷酸化位点,AdPOD64含有4个由内向外和2个由外向内的跨膜螺旋结构,磷酸化位点个数为38。二级结构预测表明,AdPOD27中-螺旋和-折叠分别占45.9%和4.6%,而AdPOD64中二者含量分别为47.8%和2.5%。SWISS-MODEL预测表明, AdPOD27和AdPOD64与棕榈(, PDB: 4usc.1.A)的相似度分别达到45.12%和46.44%,以此为模型预测2个猕猴桃POD的三维结构(图4)。
2.5 相关性分析
采用SPSS19.0进行Pearson相关性分析, 结果表明,猕猴桃经不同处理,的表达量与POD活性呈显著正相关(=0.533,=0.023<0.05),而与的表达量呈极显著正相关(=0.947,=0<0.01)。此外,的表达量与POD活性也存在正相关关系,但不显著(=0.430,=0.075>0.05)。可见,在猕猴桃果实软化、低温响应和ABA诱导过程中,和可能相互影响,而可能通过上调和下调表达影响POD活性,从而起到更重要的调控作用。
“……这桩惨绝人寰的人身伤害案发生在28日深夜10点18分。被害人王大明是我市飞腾集团董事长兼总经理。据王大明说,凶手一共有三人,他在毫无防备的情况下被他们入室袭击,由于凶手用丝袜套头,王没有看到凶手的真面目……多年来,王大明一直致力于飞腾集团的发展壮大,为我市的经济发展……”
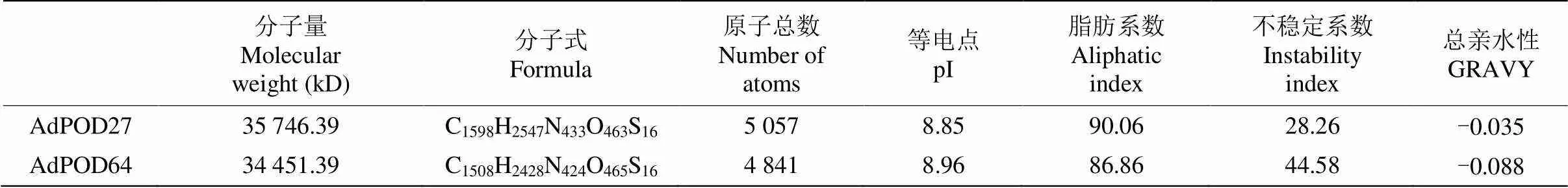
表2 AdPOD27和AdPOD64的基本理化性质
3 讨论
大量研究表明,POD在ABA介导的非生物胁迫响应过程中具有重要调控功能。外源喷施ABA能够提高水稻()幼苗的POD活性,增强其清除自由基的能力从而获得更高的抗寒能力[23]; 叶片或者根系外施ABA能够增强POD等抗氧化酶活性,维持植物膜系统和光合器官的稳定性,降低超氧阴离子和过氧化氢含量,延缓膜脂丙二醛含量增加,显著提高猕猴桃、姜()、小麦()和豌豆()等植物抗干旱能力[24–27];ABA处理还导致小麦胚胎超氧自由基、过氧化氢和氧化应激指标提高,从而诱导部分POD同工酶活性增强[28]。此外,对番茄()、甘蔗(spp.)和柽柳()等的研究也表明,ABA能够诱导上调表达[29–31]。本试验中,ABA处理的和表达量大幅度提高,说明这2个成员参与了ABA介导的非生物胁迫响应过程,然而与25℃处理相比,POD活性整体水平并没有显著变化,表明仍有其他POD同工酶参与了该过程,结果也预示着POD同工酶不同成员可能存在特异的调控功能。
低温贮藏是猕猴桃延缓果实软化,延长货架期的有效手段,然而果实低温贮藏过程中冷害常常导致果面皱缩,果肉水渍化[32]。而果实采后抗冷性与抗氧化性酶活性呈显著正相关性,通过低温锻炼或逐步降温保存可以显著增强POD等抗氧化酶活性,清除丙二醛、过氧化氢和超氧阴离子等从而明显提高果实的耐冷性[33–34]。然而,家族转录调节模式和功能丰富多样甚至在部分物种间完全相反,胡椒()的并不参与冷胁迫过程,低温敏感性的威廉蕉(sp.)低温处理后POD活性甚至下调[35]。本试验中,低温处理早期表达量和POD活性整体水平大幅度上调,而表达量并未发生显著变化,进一步验证并非所有家族成员都能响应低温诱导。此外,处理中后期表达量和POD活性均下降,与王允等[26]的研究结果相似。因此,POD同工酶参与了猕猴桃低温贮藏早期的抗冷作用,然而这种响应机制只在部分家族成员中发挥调控。
大型机械压力机大多为二到三级传动结构,第一级为电动机通过皮带带动主轴转动,第二级为主轴上的小齿轮带动曲轴上的大齿轮转动。从而通过曲柄滑块机构驱动滑块作上下往复的直线运动。因此大型机械压力机传动系统噪声主要是齿轮副的噪声。
本研究中,25℃室温贮藏软化过程中,POD活性也显著提高,但只有略微上调表达。可见,这一过程可能存在这2个成员以外的其他POD同工酶编码基因发生了转录水平改变。此外,试验中所有处理的表达量均高于,说明可能在猕猴桃果实软化、低温响应和ABA诱导等过程中发挥比更为重要的调控功能。
[1] XU H H, YANG N, ZHAO J, et al. The wondrous kiwifruit: Origin, cultivation and utilization [J]. Int J Hort, 2017, 7(1): 1–6. doi: 10.5376/ ijh.2017.07.0001.
[2] CHEN Y T, CHENG C Z, LAI R L, et al. Research progress on factors influencing kiwifruit storage [J]. Fujian J Agric Sci, 2017, 32(2): 222– 227. doi: 10.19303/j.issn.1008-0384.2017.02.023.陈义挺, 程春振, 赖瑞联, 等. 猕猴桃果实贮藏影响因子研究进展[J]. 福建农业学报, 2017, 32(2): 222–227. doi: 10.19303/j.issn.1008- 0384.2017.02.023.
[3] KASRAOUI M F, DUQUESNOY I, WINTERTON P, et al. Soluble and cell wall bound peroxidase activities are markers of flower bud development stages in lemon (L.) [J]. J Appl Bot Food Qual, 2014, 87: 1–8. doi: 10.5073/JABFQ.2014.087.001.
[4] FRANCOZ E, RANOCHA P, NGUYEN-KIM H, et al. Roles of cell wall peroxidases in plant development [J]. Phytochemistry, 2015, 112: 15–21. doi: 10.1016/j.phytochem.2014.07.020.
[5] WEVAR O A L, AGOSTINI E, TALANO M A, et al. Overexpression of a basic peroxidase in transgenic tomato (Mill. cv. Pera) hairy roots increases phytoremediation of phenol [J]. Plant Sci, 2005, 169(6): 1102–1111. doi: 10.1016/j.plantsci.2005.07. 007.
[6] XIE S. Effect of boron deficiency on lignin content and the key gene expression of lignin biosynthesis in two kinds citrus rootstocks [D]. Wuhan: Huazhong Agricultural University, 2014: 11–28.谢诗. 缺硼对两种柑橘砧木木质素含量及其合成关键基因表达的影响[D]. 武汉: 华中农业大学, 2014: 11–28.
[7] HAO C M. Physiological biochemical mechanism of section dryingfruit during storage [D]. Chongqing: Southwest University, 2009: 18–34. 郝春梅. 柑桔贮藏期枯水的生理生化机理研究[D]. 重庆: 西南大学, 2009: 18–34.
[8] ZOLFAGHARI M, SAHARI M A, BARZEGAR M, et al. Physico- chemical and enzymatic properties of five kiwifruit cultivars during cold storage [J]. Food Bioprocess Technol, 2010, 3(2): 239–246. doi: 10.1007/s11947-008-0114-6.
[9] CHOU H N, NEE C C, OU A S M, et al. Characterization of the physico-chemical and antioxidant properties of Taiwanese kiwifruit () [J]. Bot Stud, 2008, 49: 215–224.
[10] ZHU S H, SUN L N, LIU M C, et al. Effect of nitric oxide on reactive oxygen species and antioxidant enzymes in kiwifruit during storage [J]. J Sci Food Agric, 2008, 88(13): 2324–2331. doi: 10.1002/jsfa.3353.
[11] WANG C H, MENG Q K, LIU X H, et al. Effect of 1-MCP treatment on ascorbic acid content and its quality, physiological of[J]. J Chin Inst Food Sci Technol, 2014, 14(5): 134–141.王春红, 孟泉科, 刘兴华, 等. 1-MCP处理对猕猴桃中抗坏血酸含量及其品质、生理的影响[J]. 中国食品学报, 2014, 14(5): 134–141.
[12] SONG L L, GAO H Y, CHEN H J, et al. Effects of short-term anoxic treatment on antioxidant ability and membrane integrity of postharvest kiwifruit during storage [J]. Food Chem, 2009, 114(4): 1216–1221. doi: 10.1016/j.foodchem.2008.10.080.
[13] SHEN Y G, WANG W, ZHANG W, et al. Hydrogen sulfide facilitating enhancement of antioxidant ability and maintainance of fruit quality of kiwifruits during low-temperature storage [J]. Trans Chin Soc Agric Eng, 2015, 31(z1): 367–372. doi: 10.3969/j.issn.1002–6819.2015.z1. 045.
[14] SUO J T, LI H, BAN Q Y, et al. Characteristics of chilling injury- induced lignification in kiwifruit with different sensitivities to low temperatures [J]. Postharv Biol Technol, 2018, 135: 8–18. doi: 10.1016/ j.postharvbio.2017.08.020.
[15] ZHENG X L, HU B, SONG L J, et al. Changes in quality and defense resistance of kiwifruit in response to nitric oxide treatment during storage at room temperature [J]. Sci Hort, 2017, 222: 187–192. doi: 10. 1016/j.scienta.2017.05.010.
[16] DU F M, ZHANG L Y, HE L, et al. Effect of quercetin treatment on storage of kiwifruit [J]. Shaanxi J Agric Sci, 2017, 63(7): 28–32.杜芳梦, 张丽媛, 何玲, 等. 槲皮素处理对猕猴桃贮藏的影响[J]. 陕西农业科学, 2017, 63(7): 28–32.
[17] LI H, LIANG C Q, LÜ J, et al. Effects of oxalic acid treatment on lignification and related enzymes activities in ‘Huayou’ kiwifruit during cold storage [J]. Acta Hort Sin, 2017, 44(6): 1085–1093. doi: 10. 16420/j.issn.0513-353x.2016-0718.李桦, 梁春强, 吕茳, 等. 草酸对冷藏‘华优’猕猴桃果实木质化及相关酶活性的影响[J]. 园艺学报, 2017, 44(6): 1085–1093. doi: 10. 16420/j.issn.0513-353x.2016-0718.
[18] FANG L, JIANG B, ZHANG T. Effect of combined high pressure and thermal treatment on kiwifruit peroxidase [J]. Food Chem, 2008, 109 (4): 802–807. doi: 10.1016/j.foodchem.2008.01.017.
[19] BENLLOCH-TINOCO M, IGUAL M, RODRIGO D, et al. Comparison of microwaves and conventional thermal treatment on enzymes activity and antioxidant capacity of kiwifruit puree [J]. Innov Food Sci Emerg, 2013, 19: 166–172. doi: 10.1016/j.ifset.2013.05.007.
[20] CHEN K S, LI F, ZHANG S L, et al. Role of abscisic acid and indole- 3-acetic acid in kiwifruit ripening [J]. Acta Hort Sin, 1999, 26(2): 81– 86. doi: 10.3321/j.issn:0513-353X.1999.02.003.陈昆松, 李方, 张上隆, 等. ABA和IAA对猕猴桃果实成熟进程的调控[J]. 园艺学报, 1999, 26(2): 81–86. doi: 10.3321/j.issn:0513- 353X.1999.02.003.
[21] CHEN Y T, LAI R L, CHENG C Z, et al. Cloning and expression of polygalacturonase gene () during fruit softening of kiwifruit () [J]. J Agric Biotechnol, 2017, 25(2): 205–213. doi: 10.3969/j.issn.1674-7968.2017.02.004.陈义挺, 赖瑞联, 程春振, 等. 猕猴桃果实软化过程中基因的克隆与表达[J]. 农业生物技术学报, 2017, 25(2): 205–213. doi: 10. 3969/j.issn.1674-7968.2017.02.004.
[22] ZHAN J H, LAN Z H. Effects of external ABA on peroxidase isozyme of rice to low temperature stress [J]. Biotechnology, 2003, 13(1): 7–9. doi: 10.3969/j.issn.1004-311X.2003.01.005.詹嘉红, 蓝宗辉. 外源ABA对低温胁迫水稻过氧化物酶同工酶的影响[J]. 生物技术, 2003, 13(1): 7–9. doi: 10.3969/j.issn.1004-311X. 2003.01.005.
[23] LATIF H H H. Physiological responses ofplant to exogenous ABA application under drought conditions [J]. Pak J Bot, 2014, 46(3): 973–982.
[24] WANG Y L, MA F W, LI M J, et al. Physiological responses of kiwifruit plants to exogenous ABA under drought conditions [J]. Plant Growth Regul, 2011, 64(1): 63–74. doi: 10.1007/s10725-010-9537-y.
[25] WANG Y, ZHANG Y, LIU C Y, et al. Effects of exogenous abscisic acid on active oxygen metabolism in ginger leaves under drought stress [J]. Acta Hort Sin, 2016, 43(3): 587–594. doi: 10.16420/j.issn.0513- 353x.2016-0034.王允, 张逸, 刘灿玉, 等. 干旱胁迫下外源ABA对姜叶片活性氧代谢的影响[J]. 园艺学报, 2016, 43(3): 587–594. doi: 10.16420/j.issn. 0513-353x.2016-0034.
[26] LI X M, ZHANG L H, HE X Y, et al. Effects of abscisic acid on photosynthetic characteristics and antioxidant enzyme activities of wheat seedlings exposed to UV-C [J]. Chin J ApplEcol, 2006, 17(5): 822–826.李雪梅, 张利红, 何兴元, 等. 脱落酸对UV-C胁迫下小麦幼苗光合特性及抗氧化酶活性的影响[J]. 应用生态学报, 2006, 17(5): 822–826.
[27] SHARMA A D, RAKHRA G, MEHTA S, et al. Accumulation of class-III type of boiling stable peroxidases in response to plant growth hormone ABA incultivars [J]. Plant Sci Today, 2014, 1(1): 3–9. doi: 10.14719/pst.2014.1.1.6.
[28] GUO X H, JIANG J, WANG B C, et al., a truncated polypeptide from, conferred drought tolerance in[J]. Mol Biol Rep, 2010, 37(3): 1183–1190. doi: 10.1007/s11033-009- 9484-8.
[29] WANG C J, CHAN Y L, SHIEN C H, et al. Molecular characterization of fruit-specific class III peroxidase genes in tomato () [J]. J Plant Physiol, 2015, 177: 83–92. doi: 10.1016/j.jplph. 2015.01.011.
[30] SU Y C, WANG Z Q, LI Z, et al. Molecular cloning and functional identification of peroxidase geneJ]. Acta Agron Sin, 2017, 43(4): 510–521. doi: 10.3724/SP.J.1006.2017.00510.苏亚春, 王竹青, 李竹, 等. 甘蔗过氧化物酶基因的克隆与功能鉴定[J]. 作物学报, 2017, 43(4): 510–521. doi: 10.3724/SP.J. 1006.2017.00510.
[31] BURDON J, LALLU N, FRANCIS K, et al. The susceptibility of kiwifruit to low temperature breakdown is associated with pre-harvest temperatures and at-harvest soluble solids content [J]. Postharv Biol Technol, 2007, 43(3): 283–290. doi: 10.1016/j.postharvbio.2006.09.011.
[32] YANG Q Z, RAO J P, YI S C, et al. Antioxidant enzyme activity and chilling injury during low-temperature storage of kiwifruit cv. Hong- yang exposed to gradual postharvest cooling [J]. Hort Environ Bio- technol, 2012, 53(6): 505–512. doi: 10.1007/s13580-012-0101-8.
[33] YANG Q Z, ZHANG Z K, RAO J P, et al. Low-temperature condi- tioning induces chilling tolerance in ‘Hayward’ kiwifruit by enhancing antioxidant enzyme activity and regulating endogenous hormones levels [J]. J Sci Food Agric, 2013, 93(15): 3691–3699. doi: 10.1002/ jsfa.6195.
[34] ZHANG Q, ZHANG J Z, CHOW W S, et al. The influence of low temperature on photosynthesis and antioxidant enzymes in sensitive banana and tolerant plantain (sp.) cultivars [J]. Photosynthetica, 2011, 49(2): 201–208. doi:10.1007/s11099-011-0012-4.
[35] WANG J E, LIU K K, LI D W, et al. A novel peroxidasegene of pepper is involved in defense responses toinfection as well as abiotic stress tolerance [J]. Int J Mol Sci, 2013, 14(2): 3158–3177. doi: 10.3390/ijms14023158.
Cloning and Expression Analysis ofGenes in Kiwifruit
CHEN Yi-ting1, LAI Rui-lian1, FENG Xin1, GAO Min-xia1, CHENG Chun-zhen2, CHEN Wen-guan, WU Ru-jian1*
(1. Fruit Research Institute, Fujian Academy of Agricultural Sciences, Fuzhou 350013, China;2. Institute of Horticultural Biotechnology, Fujian Agriculture and Forestry University, Fuzhou 350002, China)
To understand the regulation function of peroxidase genes in kiwifruit, two members ofgene family (and) were clonedfrom‘Miliang-1’ by using reverse transcription PCR, the bioinformations of AdPOD27 and AdPOD64 were analyzed, and the correlation between their expression and POD activity were also studied.The results showed that the length of open read frame ofandwas 984 and 957 bp, encoding 327 and 318 amino acids, respectively, which GenBank accession No. were MF774100 and MF774101. AdPOD27 and AdPOD64 were hydrophilic and alkaline proteins belonging to class III of the plant heme-dependent peroxidase superfamily. The two proteins had signal peptide, transmembrane structure and phosphorylation sites. AdPOD27 and AdPOD64 located in mitochondrion and extracellular region, respectively. qRT-PCR analysis showed that the expression ofwas up-regulated greatly treated with abscisic acid (ABA) and under 4℃, whileexpression only enhanced treated with ABA. Moreover, theexpression had significant correlations with POD activity andexpression. Therefore,andwould be involved in fruit softening, low temperature response and ABA induction of.
Kiwifruit; Peroxidase; Fruit softening; Low temperature; Abscisic acid
10.11926/jtsb.3919
2018-04-04
2018-07-02
福建省自然科学基金项目(2017J01044, 2018J05051); 福建省农业科学院博士科研启动基金项目(DC2017-2); 福建省农业科学院科技创新团队项目(STIT2017-3-6); 福建省公益类科研院所基本科研专项(2015R1014-3)资助
This work was supported by the Natural Science Foundation in Fujian (Grant No. 2017J01044, 2018J05051), the Doctoral Research Starting Fund of Fujian Academy of Agricultural Sciences (Grant No. DC2017-2), the Project for Science and Technology Innovation Group of Fujian Academy of Agricultural Sciences (Grant No. STIT2017-3-6), the Project for Basic Science and Technology of Public Welfare Scientific Research Institution in Fujian Province (Grant No. 2015R1014-3).
陈义挺(1972~ ),男,博士,副研究员,研究方向为果树生物技术与遗传资源。E-mail: chyiting@163.com
E-mail: wurujian@126.com
