桑树同源四倍体桑叶产量的遗传规律研究
2019-02-12朱方容林强肖丽萍
朱方容,林强,肖丽萍
(广西壮族自治区蚕业科学研究院,南宁市530007)
植物多倍体具有繁茂性、硕大性、适应力强、生物产量高、叶片及果实营养成分含量较高等特性,这是形成新种所必需的特性,有的多倍体还经历了二倍化的过程,让新的多倍体在基因组水平和细胞学水平上更像二倍体[1-2]。有研究表明,多倍体植物的产品质量具有遗传性的规律。刘华[3]对异源四倍体花生进行了产量和品质相关性状的遗传分析,结果表明:茎高、侧枝长和单株饱果数等3个性状遗传规律符合多基因遗传模型,总分枝数、结果枝数、百果重、百仁重、出仁率、蛋白质、脂肪、油亚比等8个性状遗传规律符合2对主基因加多基因遗传模型(E);单株果重、油酸和亚油酸等3个性状遗传规律符合3对主基因遗传模型(F)。倪红梅[4]对96份桑树地方品种农艺性状进行关联分析,其研究结果对桑树种质资源重要农艺性状基因的发掘和利用以及遗传育种都具有一定的指导意义。
在桑树数量遗传规律及杂种优势的研究方面,笔者曾报道桑树(二倍体)对花叶病的抗性遗传符合加性-显性模型,主要受加性基因控制,广义遗传力和狭义遗传力中等,一般配合力显示绝对优势,具有超显性现象,母本品种含有比父本品种更多的显性基因,其中母本8006、中桑5801离原点最近,含显性基因最多;中桑5801的Wr小于0,说明该亲本存在超显性。父本伦109、7722含有隐性基因最多。因此,在组配桑树杂交组合时,选择抗花叶病的雌性桑品种作亲本尤为重要[5]。桑叶产量和质量是影响蚕茧产量和质量的重要因素,因此桑叶产量是桑树品种选育的重要目标,但有关其遗传规律和杂交优势规律的研究仍然较少,且关于四倍体桑树桑叶产量的遗传规律的研究还未见有报道。本研究以桑树同源四倍体为亲本,按NCII设计杂交组合,对桑叶产量进行配合力分析、遗传参数估测和遗传分析,旨在为桑树四倍体的开发利用和提高桑树育种效率提供参考依据。
1 材料与方法
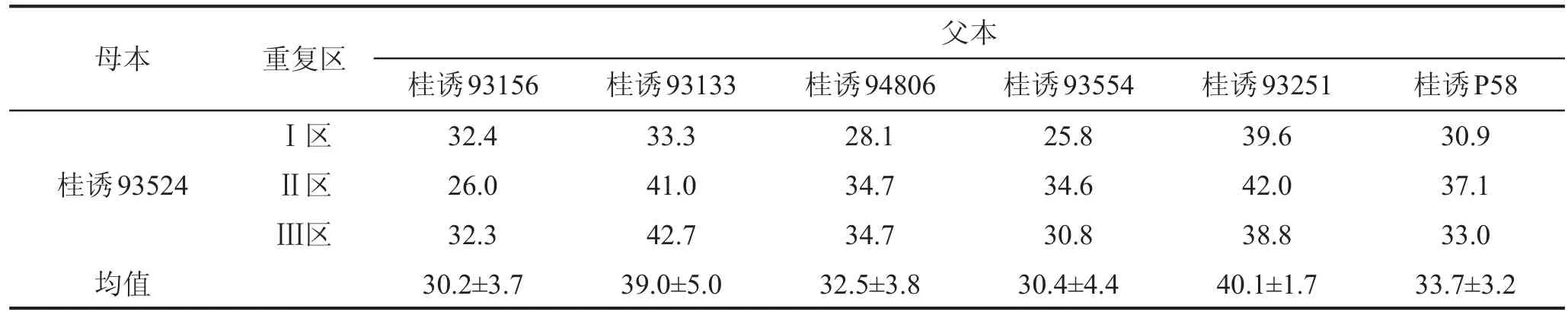
选择广西壮族自治区蚕业技术推广总站原桑树育种桑园第6杂交组合试验圃符合4×6不完全双列杂交的组合24个。母本4个,为桑树同源四倍体品种(育种材料)桂诱92L44、桂诱93200、桂诱942120、桂诱93524;父本6个,为同源四倍体品种(育种材料)桂诱P58、桂诱93156、桂诱93133、桂诱94806、桂诱93554、桂诱93251,母本和父本桑树约5年树龄,采用人工授粉方式人工杂交制杂交组合种子,每个杂交组合(F1)种植3个重复小区,按行距80 cm、株距20 cm种植,各小区同样的管理条件,种植当年即第一年就投产,投产后调查各小区第一年与第二年的桑叶产量。
参照倪先林等[6]的方法计算杂种优势。平均优势(HRav)是指杂交一代数量性状比参试组合同一性状平均值的增长率;对照优势(HRck)是指杂交一代数量性状比对照组合的增长率。CK)/CK×100。式中的F1为杂交组合的桑叶产量,为参试24个杂交组合的平均桑叶产量;CK为对照品种(杂交组合)沙2×伦109的桑叶产量。
按文献[7-9]介绍的分析法进行杂交配合力分析、遗传参数估测;用Hayman法作遗传分析。一般配合力(GCA),按估算父本一般配合力(GCAS);按估算母本一般配合力(GCAD);按估算杂交组合的特殊配合力(SCA)。式中的xi为i父本所配组合的平均产量;xj为j母本所配组合的平均产量;xij为j母本与i父本的杂交一代的桑叶产量。用EXCEL 2010进行统计分析。
2 结果与分析
2.1 桑树四倍体NCII杂交设计的产叶量及配合力分析
4个母本分别与6个父本杂交共得24个杂交组合,其F1小区群体第一年与第二年小区群体桑叶产量之和见表1。方差分析的结果见表2。结果表明,不同基因型间存在极显著的差异,而重复间差异不显著,说明试验控制良好,各基因型间有真实的差异,可进一步进行配合力分析。
配合力方差分析表明,桑叶产量母本的一般配合力达极显著水平,父本的一般配合力达显著水平,特殊配合力达极显著水平。说明父本、母本的一般配合力和组合的特殊配合力对F1的桑叶产量有显著的影响,供试的10个桑树四倍体间存在真实的遗传差异。

表1 桑树四倍体NCII杂交设计的桑叶产量 kg
续表1

表2 桑树四倍体桑叶产量的配合力方差分析
2.2 配合力效应分析
对配合力效应进行分析,结果见表3。正向效应即是有利于提高产量的效应。10个四倍体亲本的一般配合力(GCA)效应中,桂诱92L44、桂诱93251、桂诱 93554、桂诱 93133、桂诱 P58、桂诱 942120、桂诱93524共7个亲本是正向效应,其中桂诱92L44正向效应最大,其次是桂诱93251。桂诱93200、桂诱93156、桂诱94806这3个亲本是负向效应。
很多杂交组合特殊配合力效应值(SCA)的绝对值比其父本、母本一般配合力效应值的绝对值要大得多。正向特殊配合力效应最大的组合为桂诱93200×桂诱P58,该组合的桑叶产量最高;其次是桂诱942120×桂诱94806,最小的组合为桂诱93200×桂诱93554。负向特殊配合力效应突出的为桂诱942120×桂诱P58,其次为桂诱93200×桂诱93133。
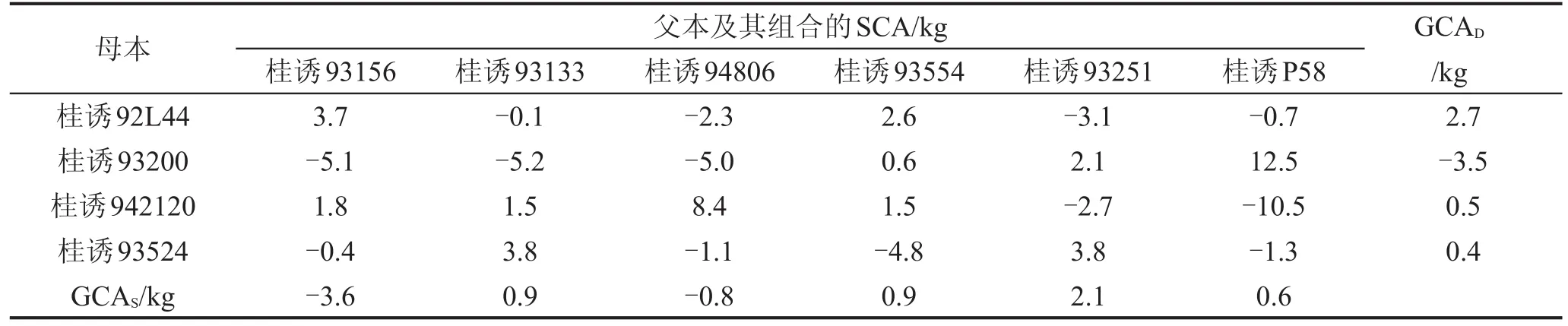
表3 桑树四倍体一般配合力效应GCA和特殊配合力效应SCA
2.3 杂种优势的表现及与特殊配合力效应的关系
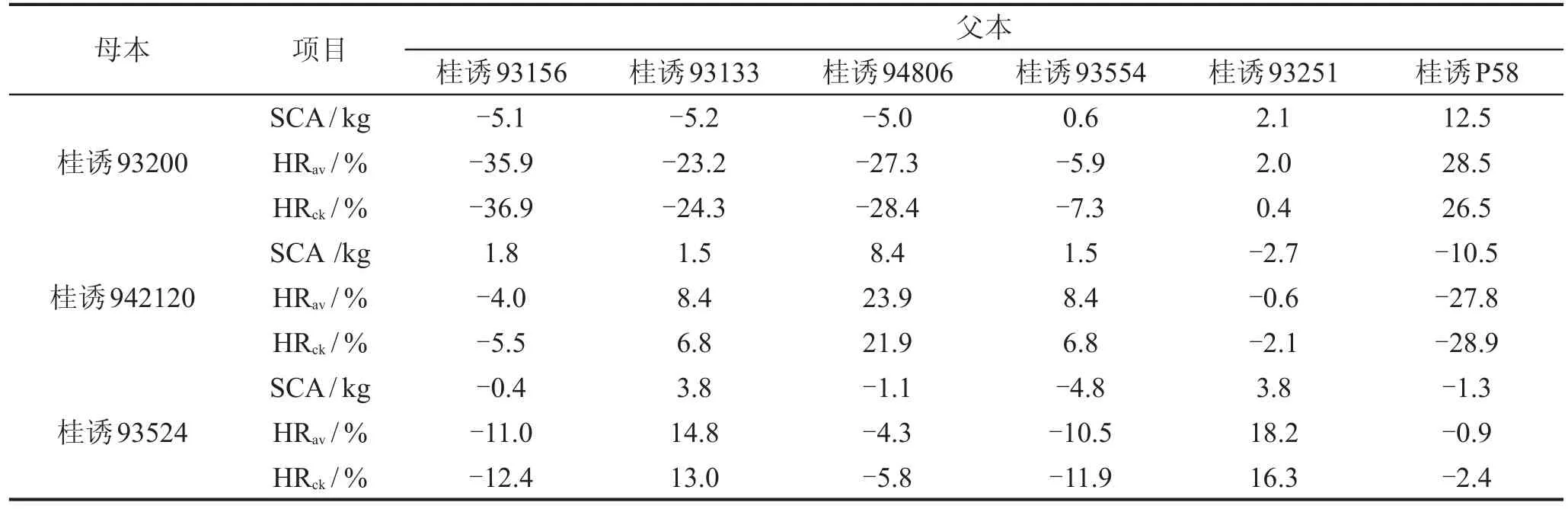
24个杂交组合的平均桑叶产量为34.0 kg,同试验的对照品种(杂交组合)沙2×伦109的桑叶产量为34.5 kg,两者差异不大,因此平均优势与对照优势差异也不大。由表4可见,平均优势与对照优势的正向优势与负向优势均为12个组合,说明桑树四倍体杂交F1,桑叶产量出现正向杂交优势与负向优势的机率是相同的。但负向优势较大的组合数较多,平均优势、对照优势的绝对值大于20%的组合数,负向优势分别有4个组合,正向优势分别只有2个组合;且负向优势最大值(绝对值)比正向优势最大值高7.4%和10.4%。

表4 桑树四倍体杂交组合的平均优势、对照优势与特殊配合力效应值
续表4
平均优势与对照优势的正向优势大于15%的组合均有4个;正向优势最大的为桂诱93200×桂诱P58,平均优势(即比平均水平增产率)达28.5%,对照优势(即比对照增产率)达26.5%;其次是桂诱942120×桂诱94806,平均优势、对照优势分别达23.9%、21.9%。负向优势最大的为桂诱93200×桂诱93156,其平均优势、对照优势分别达-35.9%、-36.9%;其次是桂诱942120×桂诱P58,平均优势为-27.8%,对照优势为-28.9%;再次是桂诱93200×桂诱94806,其平均优势、对照优势分别达-27.3%、-28.4%。桑树四倍体桂诱93200、桂诱P58、桂诱942120、桂诱94806组配成杂交组合,其桑叶产量容易出现极端的正向或负向杂种优势,即这些组合的产叶量有可能是最高的,也有可能是最低的。

图1 特殊配合力效应值SCA与对照优势HRav、平均优势HRck的回归图
图1为24个杂交组合桑叶产量的特殊配合力效应与平均优势、对照优势回归图。特殊配合力效应与平均优势、对照优势呈高度正相关,相关系数同为0.852 5,达0.01%极显著水平。特殊配合力与平均优势的回归方程:y=2.944 1x,即平均优势等于特殊配合力效应值的2.944 1倍。特殊配合力与对照优势的回归方程:y=2.898 5x-1.549 9。
2.4 遗传参数估计
估测桑树四倍体桑叶产量的遗传参数,结果见表5。由母本、父本的一般配合力效应所引起的基因型方差σS2和σD2占遗传方差总和(σG2)的比重(Vg)仅为12.26%,而杂交组合的特殊配合力效应的基因型方差σSD2占遗传方差总和的比重(VSD)为87.74%;广义遗传力hB2即为遗传方差占表现型方差的方差的比率,为70.96%;狭义遗传力(hN2)即为母本、父本的一般配合力效应所引起的基因型方差σS2和σD2之和占表现型方差的比率,仅为8.70%。结果表明,育种中仅依靠四倍体亲本桑叶产量表现难以预测选择到高产杂交组合的。桑树四倍体杂交后是否出现杂种优势,是能否选育到高产多倍体杂交组合的关键。平均显性度(H/D)1/2为5.35,(H/D)1/2>1,表明有超显性作用。

表5 桑树四倍体桑叶产量的方差分量和遗传参数估值
2.5 Wr-Vr的回归分析
用Hayman法作遗传分析,计算不完全双列杂交中各阵列的Vr和Wr值,其中Vr为亲本阵列i的方差,Wr为Ki里各家系平均数在与非轮回亲本表型的协方差,以Vr为自变量、以Wr为因变量进行回归分析。回归图见图2-a。4×6不完全双列杂交的回归系数为0.014 4,数值很小,不达显著水平,回归关系不明显。
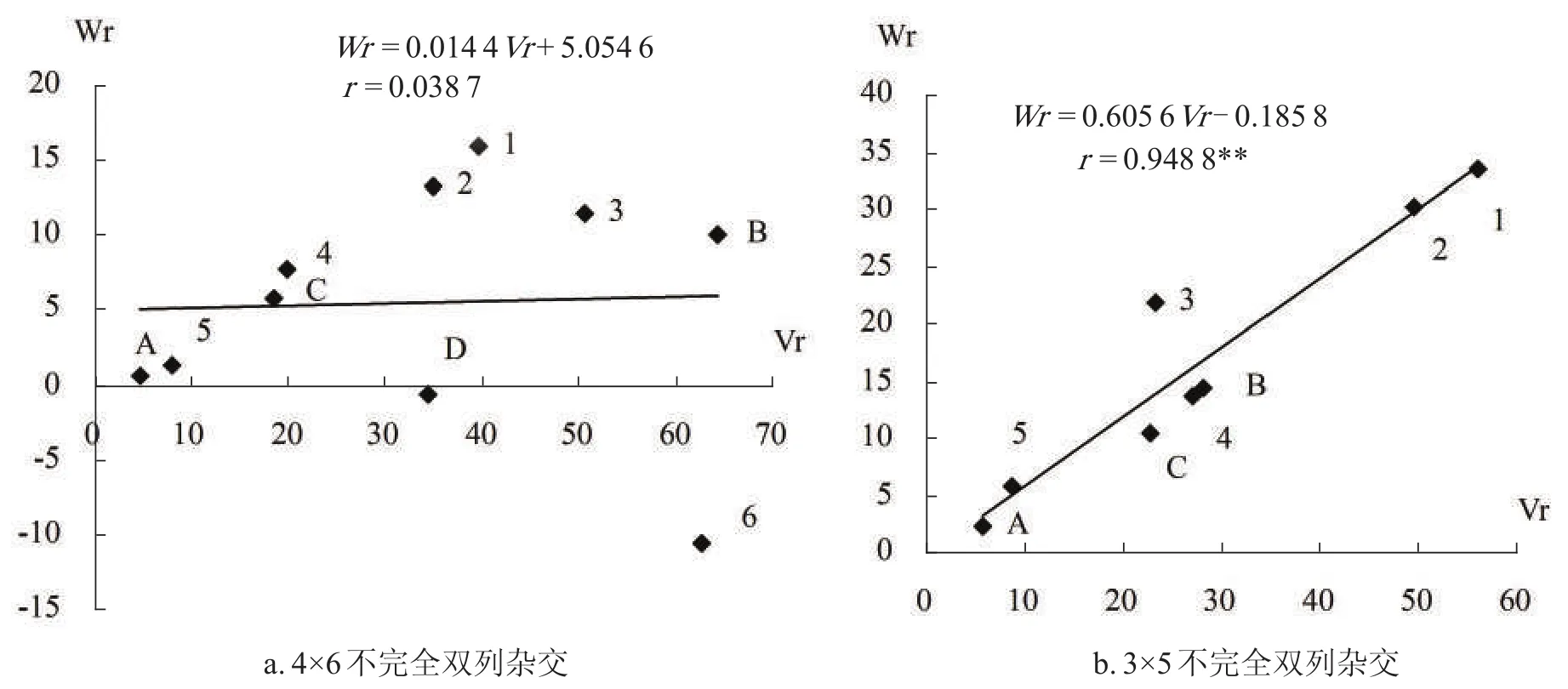
图2 桑树四倍体桑叶产量的(Wr-Vr)回归图
把具有极端(Wr-Vr)值的桂诱942120、桂诱P58两个阵列除去,余下3个母本、5个父本构成3×5不完全双列杂交,再作回归分析,结果如图2-b。得到回归方程Wr=0.605 6Vr-0.185 8,相关系数r=0.948 8,极显著;回归系数b为0.605 6,Sb为0.082 3,回归系数b与0的差异达极显著水平(tb0=7.358 3),b与1差异极显著(tb0=4.791 4),b与0.5差异不显著,因此,Hayman假设成立,四倍体的桑叶产量的遗传方式符合加性—显性模型,不存在上位性作用。截距a为-0.185 8,数值极小,Sa为2.644 1,a与0差异不显著(ta0=-0.070 2),但a<0,表明桑叶产量显性作用方式为超显性。
由各亲本在Wr-Vr回归图上的位置,可以推断各亲本显隐性基因的分布。桂诱92L44、桂诱93251离原点较近,暗示这两个亲本含显性基因较多,桂诱93156与桂诱93133离原点较远,暗示含有较少的显性基因。图2-a中,桂诱P58的Wr小于0,且与0距离甚远,表明桂诱P58的显性方式为超显性。
3 讨论
本研究参试的杂交组合的亲本是桑树同源四倍体品种,是通过诱变二倍体种子刚萌芽小苗的生长点细胞的染色体加倍、经过定向培育而育成的。由于细胞染色体加倍,基因组数也加大一倍,因此四倍体杂交后代的异基因杂合性提高,增加优良基因组合和互作的概率。李亚娟等[8]的研究显示,同源四倍体水稻杂交一代主要农艺性状具有明显的杂交优势。本研究结果表明,桑树同源四倍体杂交一代的桑叶产量,具有明显的杂交优势,正向杂交优势与负向杂种优势出现的机率相同,正向优势达15%的共有4个杂交组合,占24个参试组合的16.7%,说明利用桑树同源四倍体作亲本组配杂交组合,能够获得桑叶产量杂交优势较高的组合。
父本、母本的一般配合力和组合的特殊配合力对F1的桑叶产量有显著的影响,供试的10个桑树四倍体间存在真实的遗传差异。其中7个亲本的一般配合力效应为正向的,只有3个亲本的一般配合力效应是负向的。正向一般配合力效应值大多较小,多数杂交组合特殊配合力效应值(绝对值)比其父本、母本一般配合力效应值(绝对值)要大得多。
桑树四倍体桑叶产量的广义遗传力hB2为70.96%,表明桑叶产量的表现主要是由遗传因素的作用所决定,环境因素对产量表现有一定影响,但所起作用相对较小。狭义遗传力(hN2)较小,仅为8.70%,原因是父母本基因型方差σS2和σD2占遗传方差总和(σG2)的比重(Vg)仅为12.26%,说明加性效应在桑树四倍体桑叶产量的遗传变异中没有担负重要角色,子代的桑叶产量并没有因基因累加而显著提高。亲本桑叶产量高的,其子代产量不一定高。桑树四倍体桑叶产量的平均显性度(H/D)1/2>1,表明桑树四倍体桑叶产量的遗传有超显性作用。Wr—Vr的回归分析表明,四倍体的桑叶产量的遗传方式符合加性—显性模型,不存在上位性作用,显性作用方式为超显性。
桂诱92L44、桂诱93251含有较多的显性基因,其一般配合力正向效应也较大,但是两者杂交的特殊配合力效应却是负向的,平均优势仅4.6%、对照优势仅3.0%,杂种优势不够强。桂诱P58的一般配合力效应较小,桂诱93200一般配合力效应为负向、显性基因也不多,但是两者杂交的特殊配合力效应却较大,平均优势高达28.5%、对照优势高达26.5%,这可能与桂诱P58桑叶产量超显性遗传有关。以桂诱P58为父本,与二倍体为母本组配育成的超高产杂交组合桑特优1号和桑特优2号,其桑叶产量同样表现很强的杂种优势。桂诱P58桑叶产量的超显性遗传得以发挥可能是产生较强杂种优势的重要原因。
