草地贪夜蛾卵巢发育分级与繁殖潜力预测方法
2019-02-10赵胜园杨现明和伟张浩文姜玉英吴孔明
赵胜园 杨现明 和伟 张浩文 姜玉英 吴孔明

摘要 :草地贪夜蛾已成为严重威胁我国粮食生产的重大入侵害虫,生产上需要精准的测报技术。卵巢发育分级是鳞翅目害虫短期测报的主要手段,但目前尚缺乏对草地贪夜蛾卵巢发育分级技术的研究。本文对草地贪夜蛾卵巢发育分级与繁殖潜力预测方法的研究表明,草地贪夜蛾卵巢管发育始于蛹期,成虫卵巢发育可分成5个级别:乳白透明期(Ⅰ级)、卵黄沉积期(Ⅱ级)、成熟待产期(Ⅲ级)、产卵盛期(Ⅳ级)及产卵末期(Ⅴ级)。在25℃下,草地贪夜蛾卵巢发育Ⅰ~Ⅴ级成虫平均日龄分别为1.22、2.24、4.26、6.68及9.08 d,卵巢管平均长度分别为42.12、45.55、61.75、48.88和19.45 mm,卵巢管平均宽度分别为272.21、329.40、490.16、567.55和462.42 μm,卵粒平均直徑分别为126.57、339.40、490.16、477.55和489.97 μm,卵巢管抱卵量分别为1 251.68、1 747.04、1 809.92、732.64和81.12粒。卵巢发育分级与卵巢管长度及卵巢管宽度符合三次多项式方程关系,与卵粒直径及抱卵量符合二次方程关系。以上结果提出了草地贪夜蛾卵巢发育分级与繁殖潜力的预测方法,为草地贪夜蛾种群发生期和发生量的预测预报工作提供了技术支撑。
关键词 :草地贪夜蛾; 卵巢发育级别; 繁殖潜力; 测报
中图分类号:
S 435.132
文献标识码: A
DOI: 10.16688/j.zwbh.2019413
Ovarian development gradation and reproduction potential prediction in Spodoptera frugiperda
ZHAO Shengyuan1, YANG Xianming1, HE Wei1, 2, ZHANG Haowen1, JIANG Yuying3, WU Kongming1
(1. State Key Laboratory for Biology of Plant Diseases and Insect Pests, Institute of Plant Protection, Chinese Academy of
Agricultural Sciences, Beijing 100193, China; 2. College of Plant Protection Henan Agricultural University, Zhengzhou
450002, China;
3. China National Agro Tech Extension and Service Center, Beijing 100125, China)
Abstract
The fall armyworm, Spodoptera frugiperda (J. E. Smith) is threatening crop production in China as an important invasive pest, and monitoring and forecasting method of S.frugiperda is needed for its effective control. Ovary development gradation is an important technique in pest monitoring and forecasting. In this paper, we studied the methods for ovarian development gradation and reproduction potential prediction of S.frugiperda. Our results showed that the pupae of the insect has an immature ovary, and the adult ovaries can be graded into five stages, including transparent opalescent phase (Ⅰ), yolk deposition phase (Ⅱ), egg maturation phase (Ⅲ), peak phase of oviposition (Ⅳ) and terminal phase of oviposition (Ⅴ). The mean day age forⅠ Ⅴ females were 1.22,2.24,4.26,6.68 and 9.08 days at 25℃. The mean length of the ovarian tubes were 42.12, 45.55, 61.75, 48.88 and 19.45 mm, the mean width of the ovarian tube were 272.21, 329.40, 490.16, 567.55 and 462.42 μm, the mean diameters of egg were 126.57, 339.40, 490.16, 477.55 and 489.97 μm, the mean number of eggs within ovarian tubes were 1 251.68, 1 747.04, 1 809.92, 732.64 and 81.12, respectively. Regression analysis revealed that ovarian development grade fit with a cubic polynomial equation with the length and width of ovarian tubes, and fit with a quadratic equation with egg diameters and the number of eggs within ovarian tubes. The methods for ovarian development gradation and reproduction potential prediction of S.frugiperda are valuable for the accurate forecast of outbreak time and occurrence quantity of the pest in the field.
Key words
Spodoptera frugiperda; ovary development stage; reproduction potential; monitoring and forecasting
草地贪夜蛾Spodoptera frugiperda (J.E.Smith,1797) 是联合国粮农组织全球预警的跨境迁飞性重大农业害虫[12]。自2019年1月入侵我国云南省以来,截至7月底该虫已快速扩散到20余省(区)1 000多县(市),并在我国南方地区严重发生为害[36]。该虫迁飞能力强,可借助气流一夜间飞行100~500 km[7]。经预测,草地贪夜蛾可对我国黄淮海乃至东北地区等玉米主产区粮食生产构成严重威胁[8]。因此,做好该虫科学防控对保障国家粮食生产安全具有重要意义。
草地贪夜蛾的科学防治需要有精准预测预报技术作为支撑。目前针对黏虫Mythimna separata、棉铃虫Helicoverpa armigera、二点委夜蛾Athetis lepigone、稻纵卷叶螟Cnaphalocrocis medinalis等重大鳞翅目害虫的测报主要基于成虫调查、卵/幼虫调查、作物被害状况及越冬调查等监测调查技术[911]。国内外,性诱剂及灯光诱捕是重要的成虫监测手段[911]。如灯诱成虫及幼虫调查技术在非洲被应用于非洲黏虫Spodoptera exempta幼虫的预测预报[12]。自草地贪夜蛾入侵非洲后,性诱剂及田间幼虫调查被推荐用于该虫的监测及预报[12]。在国内,针对草地贪夜蛾的威胁,刘杰等提出了草地贪夜蛾成虫、卵、幼虫、蛹等不同虫态的种群监测方法[13]。地面灯、高空测报灯诱集及性诱剂是草地贪夜蛾成虫的重要监测手段,但不能精准预测该虫的繁殖及迁飞潜力等参数。
根据鳞翅目害虫雌蛾卵巢的形状、卵粒发育状态以及卵黄沉积情况等指标,可对卵巢发育级别进行划分。卵巢发育级别在害虫测报中是一项重要指标,可以预测害虫的发生期及发生量。鳞翅目害虫二点委夜蛾A.lepigone卵巢发育进程可分为5个阶段:透明期、卵黄沉积期、成熟待产期、产卵盛期和产卵末期,根据田间成虫卵巢不同发育级别所占比例可预测下一代幼虫发生量[14]。雌蛾卵巢发育级别也可应用于预测迁飞害虫的繁殖潜力、种群性质和迁飞动向[1518]。如果迁飞害虫卵巢发生级别较低,表明有外迁的可能,卵巢发育级别较高将宿留当地为害,应注意防治下一代幼虫[1518]。张孝羲等利用卵巢解剖技术和卵巢发育级别鉴定,将稻纵卷叶螟C.medinalis虫源性质划分为4种类型:基本迁入型、部分迁入型、本地繁殖型和大部迁出型[19]。吴孔明等通过分析棉铃虫H.armigera卵巢发育特点得出,环渤海湾南部的河北省廊坊市和北部的辽宁省辽阳市第1代棉铃虫成虫种群主要由外地迁入,第2~3代种群多数年份来源于当地,部分年份亦有外地种群迁入[20]。目前针对草地贪夜蛾卵巢生理生化[2122]及成虫繁殖力、寿命和性诱剂田间监测[12]等方面已开展了较多的研究工作[2324],但尚未见卵巢发育分级的报道。因此,我们开展了草地贪夜蛾的卵巢发育级别及繁殖潜力预测方法的研究,旨在为该虫的种群预报及异地测报提供技术支持。
1 材料与方法
1.1 试验材料
试虫于2019年1月采自云南省普洱市江城县宝藏镇水城村冬玉米田(101°39′ E,22°40′ N),并在室内使用人工饲料饲养至第6代作为本研究试虫。幼虫饲养和试验环境为:温度(25±1)℃,相对湿度75%±5%,光周期L∥D=16 h∥8 h。
1.2 试验方法
老熟幼虫化蛹后标记日期,分别对1、3、5、7、9日龄雌蛹卵巢解剖,记录卵巢发育状态(卵巢管长度、宽度、卵粒有无)及脂肪体疏密程度。1、3、5、7、9日龄雌蛹解剖数量分别为15、15、15、32、30头;其余蛹羽化后雌雄分开存放,至羽化出蛾。每30头雌雄蛾以1∶1.5的比例放置养虫笼(30 cm×30 cm×30 cm,60目)饲养,每日以10%蜂蜜水饲喂,逐日解剖雌蛾卵巢,测量卵巢管长度、近总输卵管端卵巢管宽度和卵粒直径、卵粒总数,记录卵巢管颜色变化、脂肪体疏密和颜色变化,累计解剖雌蛾250头。
测量方法:在三目体视解剖镜(上海上光新光学科技有限公司,SZ380)下进行卵巢解剖后将待测卵巢、脂肪体、卵粒等放置于物镜测微尺(株洲卡特光电仪器有限公司,C1,DIV=0.01 mm)并通过转接环与佳能6D MarkⅡ相机连接、拍照;利用Image J 1.52a(Wayne Rasband National Institutes of Health, USA, http:∥imagej.nih.gov/ij)测量软件完成卵巢管、卵粒、卵粒数等测量。
1.3 數据分析
所有数据通过Excel汇总,计算平均值和标准差;基于SAS 9.4软件,使用REG过程进行非线性回归分析,并使用Gplot过程绘制生殖参数与发育级别关系的多项式回归曲线。
2 结果与分析
2.1 草地贪夜蛾卵巢发育分级
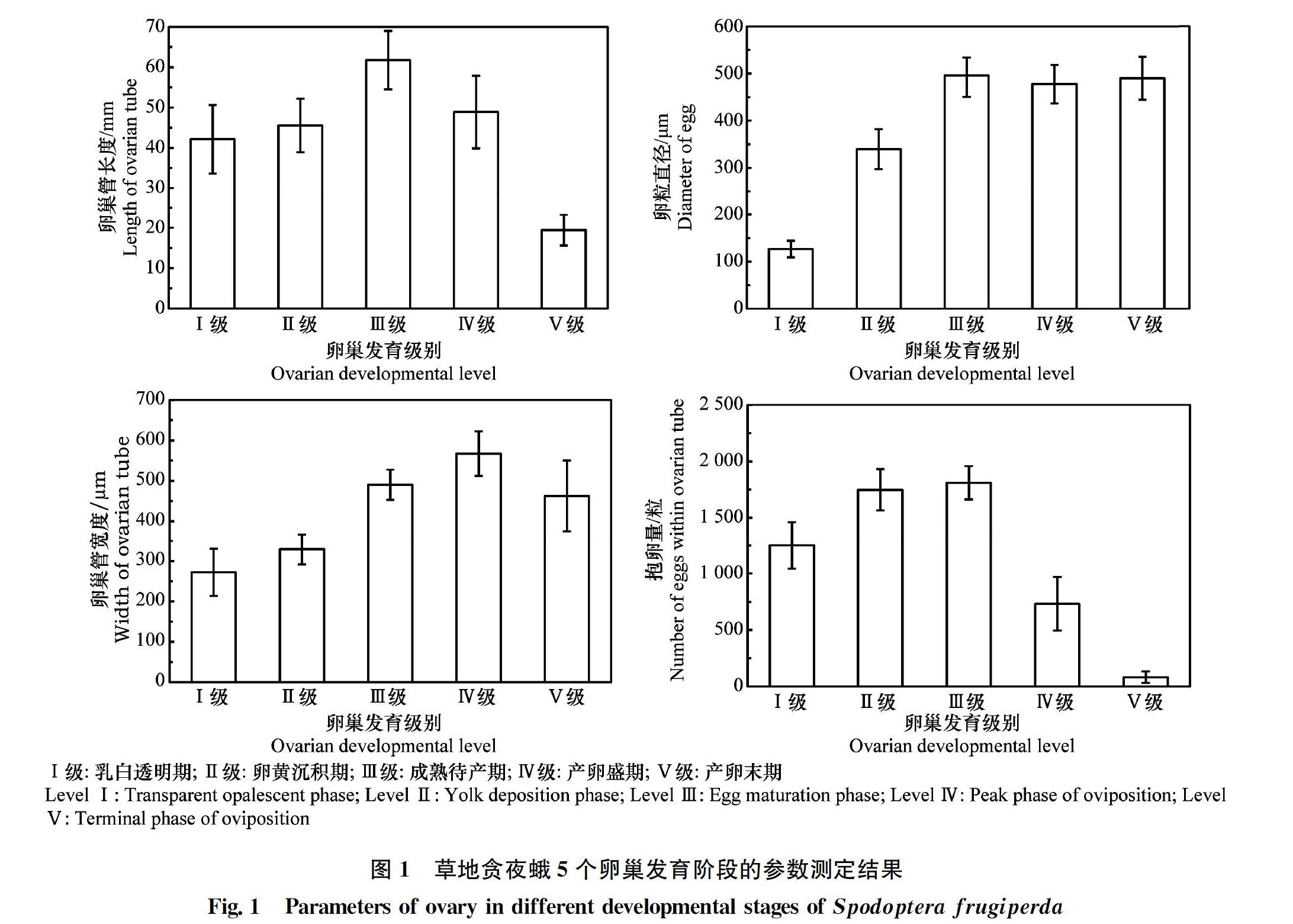
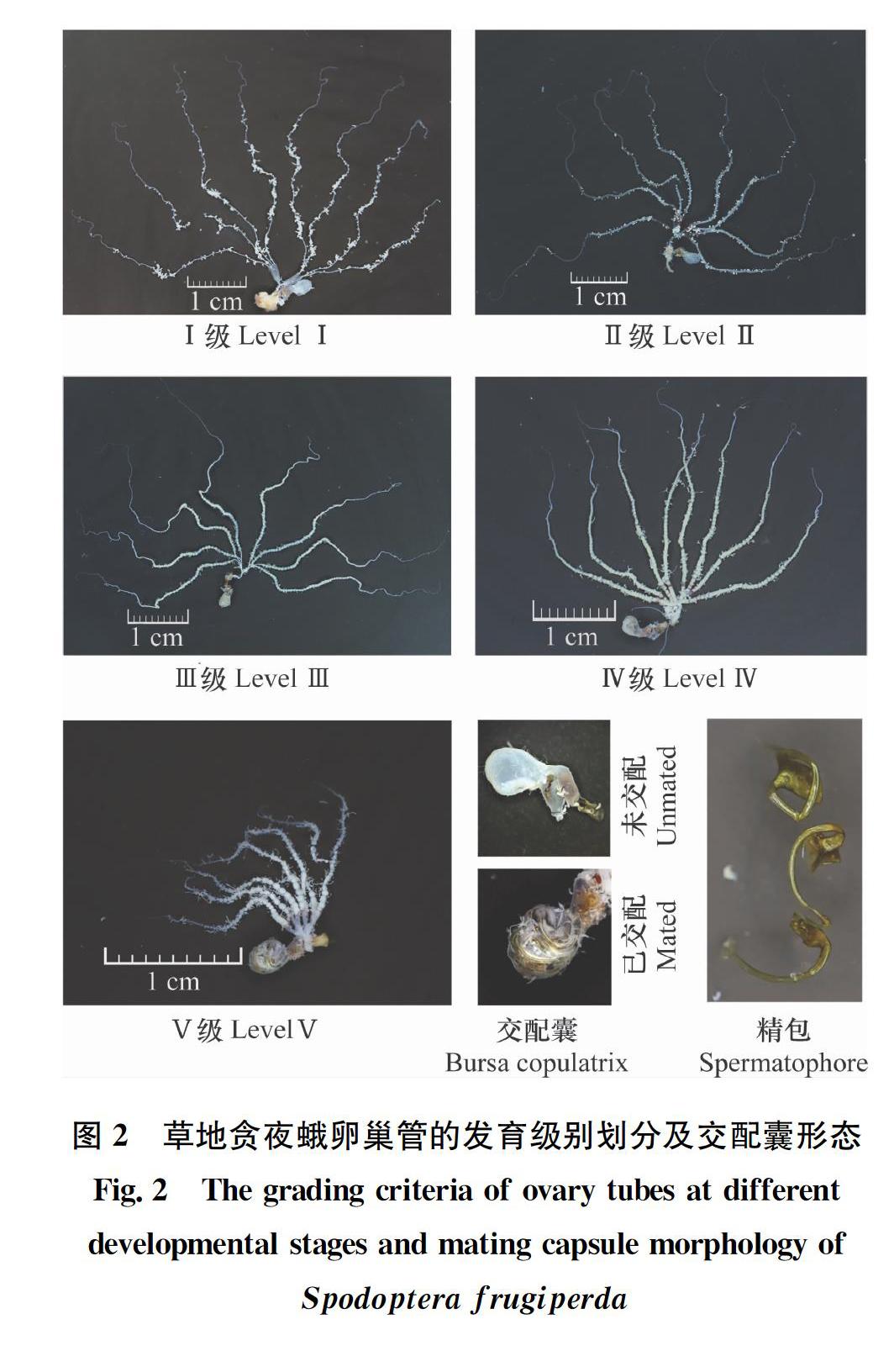
草地贪夜蛾7日龄蛹卵巢管平均长度为22.46 mm,平均宽度为256.08 μm;9日龄蛹卵巢管平均长度为26.96 mm,平均宽度为266.27 μm。根据卵巢小管内卵粒的成熟度、色泽及脂肪体多少等,可将草地贪夜蛾成虫的卵巢发育级别划分成乳白透明期(Ⅰ级)、卵黄沉积期(Ⅱ级)、成熟待产期(Ⅲ级)、产卵盛期(Ⅳ级)及产卵末期(Ⅴ级)5个发育级别(图1~2,表1)。Ⅰ级卵巢管长度平均值为42.12 mm,卵巢管宽度为272.21 μm,卵粒直径126.57 μm,抱卵量为1 251.68粒;Ⅱ级卵巢管长度平均值为45.55 mm,卵巢管宽度为329.40 μm,卵粒直径339.65 μm,抱卵量为1 747.04粒;Ⅲ级卵巢管长度平均值为61.75 mm,卵巢管宽度为49016 μm,卵粒直径496.30 μm,抱卵量为1 809.92粒;Ⅳ级卵巢管长度平均值为48.88 mm,卵巢管宽度为567.55 μm,卵粒直径477.55 μm,抱卵量为732.64粒;Ⅴ级卵巢管长度平均值为19.45 mm,卵巢管宽度为462.42 μm,卵粒直径489.97 μm,抱卵量为81.42粒(图1)。
草地贪夜蛾卵巢管各发育级别特征如下(表1,图2):乳白透明期(Ⅰ级),卵粒肉眼不可辨别,脂肪体形状不规则,并附着于卵巢管管壁,交配囊干瘪未交配;卵黄沉积期(Ⅱ级),卵粒清晰可辨,淡黄色,靠近总输卵管处有部分成熟卵粒,脂肪体乳白色,多呈葡萄串形,密布腹腔内,已有部分代谢,有一定萎缩,分枝发达,附着于卵巢管壁,交配囊乳白色,囊腔干瘪,大部分未交配;成熟待产期(Ⅲ级),卵粒饱满,大部分已成熟,呈念珠状,排列紧密,脂肪体米黄色,密度显著降低,部分脂肪粒已经干瘪,分枝发达,附着于卵巢管壁,交配囊颜色变深,至淡褐色,囊腔膨大,多已交配1~2次,交配囊中存在1~2个褐色精包,顶部干瘪球形;产卵盛期(Ⅳ级),卵粒黄绿色,饱满,已有部分成熟卵粒排出,排列稀疏,在中输卵管还存有待产卵粒,脂肪体白色,大部分脂肪粒已代谢、萎缩,仅剩丝状分枝,交配囊囊腔显著膨大,多已交配2~3次;产卵末期(Ⅴ级),卵巢管显著萎缩,绝大部分卵粒已排出,仅剩少量遗卵,几乎无脂肪粒,仅剩少量丝状分枝残存体腔或附着卵巢管壁,交配囊囊腔显著膨大,多已交配2~4次(表1,图2)。
Ⅰ乳白透明期乳白色卵巢管乳白色,长20~50 mm,平均42.12 mm,宽200~300 μm,平均272.21 μm;卵粒肉眼不可辨别脂肪体乳白色,形状不规则,多呈球形、椭球形、棍棒形,密布腹腔内,并附着于卵巢管管壁交配囊乳白色,囊腔干瘪,未交配羽化后1~2 d
Ⅱ卵黄沉积期乳白色卵巢管乳白色,长40~60 mm,平均45.55 mm, 宽200~400 μm,平均329.40 μm;卵粒清晰可辨,淡黄色,靠近总输卵管处有部分成熟卵粒脂肪体乳白色,多呈葡萄串形,密布腹腔内,因已有部分代谢而呈一定程度萎缩,分枝发达,附着于卵巢管壁交配囊乳白色,囊腔干瘪,大部分未交配羽化后2~3 d
Ⅲ成熟待产期黄白色卵巢管黄绿色,长45~80 mm,平均61.75 mm,宽400~600 μm,平均490.16 μm;卵粒饱满、黄绿色,大部分已成熟,呈念珠状,排列紧密脂肪体米黄色,密度显著降低,部分已经干瘪,分枝发达,附着于卵巢管壁交配囊颜色变深,至淡褐色,囊腔膨大,多已交配1~2次,顶部干瘪球形羽化后3~5 d
Ⅳ产卵盛期淡黄色卵巢管黄绿色,长35~75 mm,平均48.88 mm,宽400~700 μm,平均567.55 μm;卵粒饱满、黄绿色,已有部分成熟卵粒排出,排列稀疏,在中输卵管还存有待产卵粒脂肪体白色,大部分脂肪粒因代谢而萎缩,仅剩丝状分枝交配囊囊腔显著膨大,多已交配2~3次羽化后5~7 d
Ⅴ产卵末期淡白色卵巢管淡白色,长10~30 mm,平均19.45 mm,宽200~600 μm,平均462.42 μm,显著萎缩;绝大部分卵粒已排出,仅剩少量遗卵几乎无脂肪粒,仅剩少量丝状分枝残存体腔或附着卵巢管壁交配囊囊腔显著膨大,多已交配2~4次羽化后7~10 d
草地贪夜蛾卵巢5个发育级别成虫对应的平均日龄,乳白透明期(Ⅰ级)多为羽化后1~2 d成虫,平均日龄为1.22 d;卵黄沉积期(Ⅱ级)多为羽化后2~3 d成虫,平均日龄为2.24 d;成熟待产期(Ⅲ级)多为羽化后3~5 d成虫,平均日龄为4.26 d;产卵盛期(Ⅳ级)多为羽化后5~7 d成虫,平均日龄为668 d;产卵末期(Ⅴ级)多为羽化后7~10 d成虫,平均日龄为9.08 d(表1,图3)。
2.2 卵巢发育级别与生殖参数的关系
利用卵巢发育分级指标预测其繁殖潜力,对不同级别下的生殖参数,如卵巢管长度、卵巢管宽度、卵粒直径以及单支卵巢管抱卵量进行了回归分析,构建了各生殖参数与发育级别的回归方程。回归分析表明,草地贪夜蛾卵巢发育级别(x)与卵巢管长度(y1)符合三次多项式方程y1=53.93-25.49x+16.36x2-2.53x3(F3, 215=362.47, P<0.01, R2=0.837),与卵巢管宽度(y2)符合三次多项式方程y2=513.60-407.70x+211.20x2-26.25x3(F3, 202=633.10, P<0.01, R2=0.905),与卵粒直径(y3)符合二次方程y3=-161.47+336.30x-41.76x2(F2, 237=1 509.16, P<0.01, R2=0.928),与抱卵量(y4)符合二次方程y4=354.06+1 199.31x-253.82x2(F2, 232=1 141.97,P<0.01,R2=0.909)(图4)。
在5级分类指标下,卵巢管长度在Ⅰ级至Ⅲ级呈上升趋势,随着卵粒的发育成熟卵巢管逐渐伸长,Ⅲ级卵巢管最长;Ⅲ级以后,随着卵粒的排出,卵巢管逐步缩短,Ⅴ级基本无卵粒,卵巢管萎缩至最短。卵巢管宽度在Ⅰ~Ⅳ级呈上升趋势,随着卵粒成熟,卵巢管膨大变宽,Ⅳ级时达到最宽;Ⅴ级時卵巢管萎缩,宽度渐窄。随着卵粒发育,卵粒直径在Ⅰ级至Ⅲ级呈上升趋势,Ⅲ级至Ⅴ级直径降低,但不明显;同时,单条卵巢管卵粒数在Ⅰ级至Ⅱ级呈上升趋势,在Ⅱ级和Ⅲ级卵粒数最多,Ⅲ级以后卵粒数呈明显下降趋势。
3 讨论
根据卵巢小管内卵粒的成熟度、色泽及脂肪体多少,可将鳞翅目昆虫卵巢进行发育级别分级。如棉铃虫卵巢发育程度可分为6个级别,即发育初期(0级)、卵黄沉积前期(Ⅰ级)、卵黄沉积期(Ⅱ级)、成熟待产期(Ⅲ级)、产卵盛期(Ⅳ级)和产卵末期(Ⅴ级)[25]。欧洲玉米螟卵巢分为6个发育级别:无卵黄阶段、卵黄沉积阶段、卵成熟期、产卵前期、产卵高峰期、产卵末期[26]。甜菜夜蛾S. exigua、小地老虎Agrotis ipsilon、黏虫、稻纵卷叶螟等鳞翅目迁飞害虫的卵巢发育分为乳白透明期、卵黄沉积期、成熟待产期、产卵盛期和产卵末期5个级别[18,2729];而草地螟卵巢发育可分为4级,分别是初羽化期、产卵前期、产卵期、产卵末期[30]。本研究中,与大多数其他鳞翅目害虫一致,草地贪夜蛾卵巢可划分为乳白透明期(Ⅰ级)、卵黄沉积期(Ⅱ级)、成熟待产期(Ⅲ级)、产卵盛期(Ⅳ级)、产卵末期(Ⅴ级)5个发育级别。在温度(25±1)℃,相对湿度 75%±5%、光周期L∥D=16 h∥8 h条件下,卵巢处于Ⅰ级至Ⅴ级的草地贪夜蛾成虫平均日龄分别为1.22、2.24、4.26、6.68及9.08 d,卵巢管平均抱卵量分别为1 251.68、1 747.04、1 809.92、732.64和81.12粒。
昆虫迁飞与卵巢发育程度存在密切的关系。一般而言,昆虫的迁飞行为多发生在卵巢未成熟时,通常昆虫迁入种群的卵巢发育级别、交配数及交配率均较高,卵巢发育为I级的比例很低或不存在;而本地迁出虫源卵巢发育级别及交配率均较低,卵巢发育为I级的个体比例极高,而且多数未交配[1718,31]。根据研究者多年观测结果,5月到10月份,跨渤海湾迁飞的鳞翅目昆虫黄地老虎Agrotis segetum、小菜蛾Plutella xylostella、甘蓝夜蛾Mamestra brassicae、旋幽夜蛾Scotogramma trifolii及苜蓿夜蛾Heliothis viriplaca等迁飞昆虫种群雌蛾的卵巢发育级别、交配率、交配次数均呈下降趋势[3235]。可能是由于北迁种群经过了数晚的长距离迁飞(≥60 km),卵巢已经经过了一定程度的发育和交配活动,南迁种群只经过较短距离的迁飞(≥40 km),卵巢尚未发育成熟[2835]。因此,通过对迁飞昆虫卵巢进行解剖观察其卵巢发育进度,可对迁飞昆虫的虫源性质进行判别[1718]。而草地贪夜蛾为典型的迁飞性昆虫,开展草地贪夜蛾卵巢发育级别的监测,结合各发育级别的比例及动态情况,可确定田间该虫的虫源性质。如田间大多数成虫卵巢发育级别较低(Ⅰ~Ⅱ级)且持续时间较长,表明该虫有外迁的可能;如发育级别较高(Ⅲ~Ⅴ级),表明会留宿当地为害,应及时发出预报并注意下一代幼虫的防控。多地开展监测,结合气象数据及种群动态数据等,对该虫的迁飞轨迹进行模拟预测,可指导异地测报,为该虫科学防控奠定基础。
通过卵巢发育分级也可预测害虫发生期、发生量、发生趋势及防治适期等[18,36]。甜菜夜蛾卵巢Ⅲ~Ⅳ级时是产卵高峰,基于此可以预测该虫田间发生量[37]。我国科学家利用卵巢发育法成功预测了第三代稻纵卷叶螟的发生期[38]。张孝羲等利用卵巢解剖技术和卵巢发育等级鉴定,根据虫源性质确定稻纵卷叶螟防治适期,如稻纵卷叶螟蛾高峰后4~5 d,一级雌蛾仍维持在50%以上的水平,说明成虫正在大量迁出,不会对本地造成过大的为害,可以不予全面防治[19]。根据卵巢解剖结果,河北省石家庄地区2011年7月下旬至8月上旬二点委夜蛾Ⅰ级卵巢在整个发生期所占比例较高,Ⅳ级卵巢所占比例较低,推测其下一代幼虫的发生量将较低并得到证实[14]。
本研究提出了草地贪夜蛾卵巢发育级别及繁殖潜力预测方法,可用于田间种群发生期和发生量的预测预报。根据田间草地贪夜蛾卵巢发育级别,结合本研究中平均日龄,预测产卵高峰期,结合卵孵化期及幼虫发育历期可预测下一代幼虫发生期。根据各发育级别卵巢抱卵量及田间成虫发生数量,也可对下一代幼虫的发生量进行预测预报。当然,田间环境及生物因素较复杂,如草地贪夜蛾雌虫日龄对产卵力具有显著影响,雌虫及雄虫羽化后2日龄是影响产卵力和繁殖力的关键时期,延时交配会显著延长发育历期[39]。本文研究了实验室条件下的草地贪夜蛾的各卵巢发育级别、日龄及抱卵量的关系,但田间温度、湿度等环境条件较为复杂,仍需要进一步细化,完善相关研究。
参考文献
[1] Food and Agriculture Organization of the United Nations. Integrated management of the fall armyworm on maize: A guide for farmer field schools in Africa [R]. 2018. http:∥www.fao.org/3/I8665EN/i8665en.pdf.
[2] United States Agency for International Development, The International Maize and Wheat Improvement Center, The CGIAR Research Program on Maize. Fall armyworm in Africa: a guide for integrated pest management [R]. 2018. https:∥www.agrilinks.org/sites/default/files/fall armyworm ipm guide for africa jan-30 2018.pdf.
[3] 姜玉英, 刘杰, 朱晓明. 草地贪夜蛾侵入我国的发生动态和未来趋势分析[J]. 中国植保导刊, 2019, 39(2): 3335.
[4] 吴秋琳, 姜玉英, 吴孔明. 草地贪夜蛾缅甸虫源迁入中国的路径分析[J]. 植物保护, 2019, 45(2): 16.
[5] 张磊, 靳明辉, 张丹丹, 等. 入侵云南草地贪夜蛾的分子鉴定[J]. 植物保护, 2019, 45(2): 1924.
[6] 全国农业技术推广服务中心. 全国农技中心关于加强草地贪夜蛾监测预警能力建设的通知[EB/OL]. (20190623)[20190810]. https:∥www.natesc.org.cn/Html/2019-06-25/2-1878-2019-06-25-458976.html.
[7] WESTBROOK J K, NAGOSHI R N, MEAGHER R L, et al. Modeling seasonal migration of fall armyworm moths [J]. International Journal of Biometeorology, 2016, 60(2): 255267.
[8] 吳秋琳,姜玉英,胡高,等.中国热带和南亚热带地区草地贪夜蛾春夏两季迁飞轨迹的分析[J].植物保护,2019,45(3):19.
[9] 姜玉英. 我国农作物害虫测报技术规范制定与应用[J]. 应用昆虫学报, 2013, 50(3): 868873.
[10]姜玉英, 刘杰, 曾娟,等. 棉铃虫种群调查及测报技术[J]. 应用昆虫学报, 2018, 55(1): 132137.
[11]PRASAD Y G, PRABHAKAR M, ABROL D P, et al. Pest monitoring and forecasting [M]∥Integrated Pest Management,2012.
[12]ODIYO P O. Forecasting infestations of a migrant pest: the African armyworm Spodoptera exempta (Walk.)[J]. Philosophical Transactions of the Royal Society B: Biological Sciences, 1979, 287: 403413.
[13]刘杰, 姜玉英, 刘万才, 等. 草地贪夜蛾测报调查技术初探[J]. 中国植保导刊, 2019, 39(4): 4447.
[14]李立涛,王玉强,刘磊,等.二点委夜蛾卵巢发育分级及在预测预报中的应用[J].应用昆虫学报,2012,49(4):10431047.
[15]JOHNSON C G. Physiological factors in insect migration by flight [J]. Nature, 1963, 198: 423427.
[16]JOHNSON C G. Migration and dispersal of insects by flight[M]. London, UK.: Methuen & Co, 1969.
[17]齐国君,芦芳,胡高,等.卵巢解剖在我国迁飞昆虫研究中的应用[J].中国植保导刊,2011,31(7):1822.
[18]张孝羲.昆虫生态与预测预报[M].北京:农业出版社,1985:214215.
[19]张孝羲,陆自强,耿济国.稻纵卷叶螟雌蛾解剖在测报上的应用[J].昆虫知识,1979,16(3):9799.
[20]吴孔明,郭予元,吴燕.环渤海湾地区棉铃虫成虫的卵巢发育特点及与迁飞行为的关系[J].生态学报,2001,22(7):10751078.
[21]SORGE D, NAUEN R, RANGE S, et al. Regulation of vitellogenesis in the fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae)[J]. Journal of Insect Physiology, 2000, 46(6): 969976.
[22]ALVES T, CRUZ G S, WANDERLEY TEIXEIRA V, et al. Effects of Piper hispidinervum on spermatogenesis and histochemistry of ovarioles of Spodoptera frugiperda[J]. Biotechnic & Histochemistry, 2014, 89(4): 245255.
[23]BARROS E M, TORRES J B, BUENO A F. Oviposition, development, and reproduction of Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae) fed on different hosts of economic importance[J]. Neotropical Entomology, 2010, 39(6): 9961001.
[24]SIMMONS A M, LYNCH R E. Egg production and adult longevity of Spodoptera frugiperda, Helicoverpa zea (Lepidoptera: Noctuidae), and Elasmopalpus lignosellus (Lepidoptera: Pyralidae) on selected adult diets [J]. Florida Entomologist, 1990, 73(4): 665671.
[25]张万娜, 肖海军, 梁革梅, 等. 棉铃虫卵巢形态与卵子发生过程观察[J]. 昆虫学报, 2013, 56(4): 358364.
[26]KUANG X, CALVIN D D, KNAPP M C, et al. Female European corn borer (Lepidoptera: Crambidae) ovarian developmental stages: Their association with oviposition and use in a classification system[J]. Journal of Economic Entomology, 2004, 97(3): 828835.
[27]陳若篪,丁锦华,谈涵秋,等.迁飞昆虫学[M].北京:农业出版社,1989.
[28]刘永强.小地老虎跨海迁飞规律与虫源地分析[D].北京:中国农业科学院,2015.
[29]王宪辉, 徐洪富, 许永玉, 等. 甜菜夜蛾雌性生殖系统结构、发育分级及在测报上的应用[J]. 植物保护学报, 2003,30(3):261266.
[30]孙雅杰,陈鹿瑞,王素云,等.草地螟雌蛾生殖系统发育的形态变化[J].昆虫学报,1991,34(2):248249.
[31]齐国君,谢茂昌,梁载林,等.2008年桂北地区稻飞風特大迁入事件分析[J].应用昆虫学报,2011(5):12601267.
[32]GUO Jianglong, FU Xiaowei, WU Xiao. et al. Annual migration of Agrotis segetum (Lepidoptera: Noctuidae): observed on a small isolated island in northern China[J/OL]. PLoS ONE, 2015, 10(6): e0131639.
[33]HE Limei, FU Xiaowei, HUANG Yunxin. et al. Seasonal patterns of Scotogramma trifolii Rottemberg (Lepidoptera: Noctuidae) migration across the Bohai Strait in northern China[J]. Crop Protection, 2018, 106: 3441.
[34]WU Xiao, FU Xiaowei, GUO Jianglong. et al. Annual migration of cabbage moth, Mamestra brassicae L. (Lepidoptera: Noctuidae), over the sea in northern China[J/OL]. PLoS ONE, 2015, 10(7): e132904.
[35]ZHAO Xiaoyang, FU Xiaowei, ALI A. et al. Is Heliothis viriplaca (Lepidoptera: Noctuidae) a long distance migrant?[J].Bulletin of Entomological Research, 2016, 106(6): 740748.
[36]李汝鐸, 王金其, 苏德明. 昆虫卵巢发育与害虫预测预报[M]. 上海:复旦大学出版社,1987: 3184.
[37]HAN Lanzhi, GU Hainan, ZHAI Baoping, et al. Reproduction flight relationship in the beet armyworm, Spodoptera exigua (Lepidoptera: Noctuidae)[J]. Environmental Entomology, 2008, 37(2): 374381.
[38]上海市川沙县张江农科站. 应用卵巢发育法预测第3代稻纵卷叶螟发生期[J]. 昆虫知识, 1980(3): 104105.
[39]ROGERS C E, MARTI O G J. Reproductive potential of once mated moths of the fall armyworm (Lepidoptera: Noctuidae)[J]. Florida Entomologist, 1994, 77(4): 402410.
(责任编辑:杨明丽)
