植被边界带对相邻麦田地表步甲和蜘蛛分布及蚜虫发生的影响
2019-01-18张旭珠宇振荣刘云慧
张旭珠, 张 鑫,,*, 宋 潇, 宇振荣,, 刘云慧,
1 中国农业大学资源与环境学院, 北京 100193 2 河北建筑工程学院建筑与艺术学院, 张家口 075000 3 中国农业大学生物多样性与有机农业北京市重点实验室, 北京 100193
农业景观中所残留的半自然生境管理粗放,受干扰相对较小,为农田天敌提供替代食源的同时也为其提供了栖息地及避难所[1- 3],在农业景观天敌多样性保护过程中受到越来越多的重视。在欧洲,生态学家对农田半自然生境的生态学效益进行了大量评估研究[4-5],多数结果显示半自然生境对景观中天敌多样性具有重要的保护和维持功能[6],但是由于半自然植被组成不同,其为天敌提供的生存环境不同,且害虫取食过程中,诱导不同植被产生的挥发物吸引天敌的种类不同[7],因此,不同类型半自然生境所维持的天敌类群存在较大的差异[8]。此外,许多研究表明,由于不同类型生境界面间边界效应的存在,与作物田相连接的植被边界带植被类型和结构可影响害虫及其天敌迁居的物种类型、数量和时间[9-10]。因此,调查具有不同植被边界的农田中天敌组成和害虫发生状况、研究农田与不同类型植被边界带界面之间天敌分布及迁移特征是保护和增加农田中天敌数量、提高害虫生物控制效率的重要基础。
另一方面,植被边界带中维持的天敌类群通过“溢出效应”不断向农田进行扩散[11- 13],然而,因受节肢动物自身扩散能力限制,植被边界带中的天敌向农田内部扩散存在一定的有效距离[14-15],赵紫华等[10]认为边界对麦田地表及飞行类节肢动物天敌群落分布的影响尺度在30 m左右,超过30 m的区域边缘效应及溢出效应不明显,而就步甲而言,早期的研究指出其从边界到农田的扩散距离不超过50—60 m[14-15]。充足的天敌数量是实现害虫生物控制的基础,因此,深入探讨植被边界带对相邻农田害虫发生的影响,需进一步了解不同类型半自然植被边界带所维持的不同类群天敌深入农田的扩散距离。
此外,高天敌密度有时并不意味着高害虫生物控制效率[16-17]。关于天敌多样性对害虫生物控制效率的相关研究依然不足[18],且目前为止尚未达成一致结论。尽管有学者指出在自然条件下约有99%的潜在有害生物能够被天敌持续控制[19-20],依靠天敌实现害虫控制具有极大的应用潜力,但是随后的研究则证实,天敌的多样性总是与害虫密度及作物的受害程度呈正相关关系或无直接相关关系[21- 23]。因此,景观建设过程中,不应单纯以追求增加天敌数量为目标,应当同时考虑害虫发生状况,明确不同类型天敌对目标害虫的控制能力以及群落增减动态等。
目前国内针对半自然生境对天敌多样性保护、农田虫害生物控制作用做了一些初步探索,但多集中于残存农田半自然生境对天敌多样性分布格局的影响[24- 26]。对于不同类型植被边界带、人工种植边界带对天敌分布的影响及其对周围农田中天敌-害虫群落关系的探讨依然不足;且先前的研究对于天敌-害虫相互作用关系的研究也大多是从生物化学的角度探讨[9, 27],少有从生物群落数量关系方面直接探讨。因此,要实现通过重建半自然生境促进有益昆虫多样性、降低害虫群落发生数量,尚需深入探讨不同的植被管理模式对天敌和害虫群落关系的影响。
本文通过比较北京顺义地区不同类型植被边界带(人工林地、人工种植草带以及自然演替草带)相邻麦田中天敌、蚜虫的多度分布,分析边界带植被群落结构对天敌群落分布的影响,旨在回答以下问题:(1)不同类型植被边界带对相邻麦田中步甲和蜘蛛多度的维持作用是否不同?(2)不同类型植被边界带对相邻麦田中蚜虫发生的影响以及益害比是否存在差异?(3)半自然植被边界带植被结构特征对相邻麦田步甲和蜘蛛群落分布有何影响?(4)不同类型植被边界带与其相邻麦田之间步甲和蜘蛛的“溢出”效应如何?
1 研究区和研究方法
1.1 研究区概况
研究区位于北京东北部约45 km处顺义区赵全营镇(40°14′21.66″—40°13′55.35″N, 116°36′0.49″—116°34′45.25″E)。气候温和,光照充足,昼夜温差大。年平均气温9—11 ℃,年日照时数为2063 h,年平均降水625 mm,降水年内分布不均,75%以上集中在夏季。镇域面积6445 hm2,人口2.8万,耕地面积415.2 hm2,林地面积120 hm2。在过去的几十年里,为了满足日益增长的人口需求,此处成为我国北方地区典型的集约化农业种植区域。主要的作物种植类型为冬小麦、夏玉米轮作,景观中只留存了少量的自然或半自然生境,主要的半自然生境类型包括道路防风林,农田经济林,废弃地以及农田间残留的农田边界。为改善半自然植被覆盖降低所造成的一系列环境问题,半自然生境重建成为环境和生态服务恢复的重要措施。资料显示,在2012年到2015年间顺义平原地区造林面积约为6.6×105hm2[28],人工林地成为当地现存最为重要的半自然生境类型。研究区灌溉措施皆采用喷灌等节水措施。取样期间,小麦蚜虫高发期,小麦田中统一施用高效氯氟氰菊酯及吡虫啉作为杀虫剂一次(具体为5月20日至27日期间施用)。
1.2 取样点设计
2014年3月份,在研究区选取了4条路旁林下沟渠种植人工草带,沟渠深约1 m,宽约2.5 m。草带植被选取北方地区较常见且被认为能够起到促进地表节肢动物多样性的高羊茅与小冠花混播[29-30],上层乔木为国槐单行林。5月至6月,在研究区典型农业景观中选取毗邻人工草带小麦田(AW)以及与景观中两类现存重要半自然生境——自然演替草带(NW)、人工林地相邻的小麦田(WW)各4块,在与植被边界带连接的小麦田内,紧邻边界带设置大小为50 m×50 m的样方进行取样。为避免样方间相互影响,取样点间距离不小于100 m。研究区取样点位置如图1所示:

图1 取样区地理位置分布图Fig.1 Geographical location of research regionWW: 毗邻人工林地的小麦田,Wheat field neighbored by woodland; AW: 毗邻人工种植草带的小麦田,Wheat field neighbored by plant grass strip; NW: 毗邻自然演替草带的小麦田,Wheat field neighbored by succession grass strip
1.3 步甲和蜘蛛调查
2014年5月10日至6月9日,采用陷阱法对小麦田及其植被边界带交界处的地表天敌进行连续取样。其中麦田与植被边界带交界0 m处设置一排4个陷阱用于采集边界处地表天敌,麦田内部距离边界15 m及30 m处平行于农田边界分别设置两排陷阱,具体方法如下图2,分别用于捕获来自农田内部到植被边界带以及植被边界带到农田内部两个方向的地表天敌,2排陷阱中间均有20 cm长、20 cm高的塑料板相隔。每排相邻陷阱间相距10 m,陷阱口径7.3 cm、深13.5 cm,陷阱中放入约占陷阱体积2/3的饱和食盐水作为防腐剂,陷阱上部距离杯口1 cm处钻有4个直径5 mm小孔,防止样本因雨水外溢。每隔10 d将陷阱内的样品回收一次,每一排陷阱合并挡板相同方向的4个杯子为一组样品,挑取两类主要天敌类群步甲和蜘蛛保存到75%的酒精中,带回实验室制作标本,分类鉴定。步甲鉴定到种,分种鉴定主要参考《河南昆虫志:鞘翅目(一)》等[31- 33],部分标本请中国科学院动物研究所相关专家鉴定。蜘蛛标本请河北大学相关专家鉴定到科。
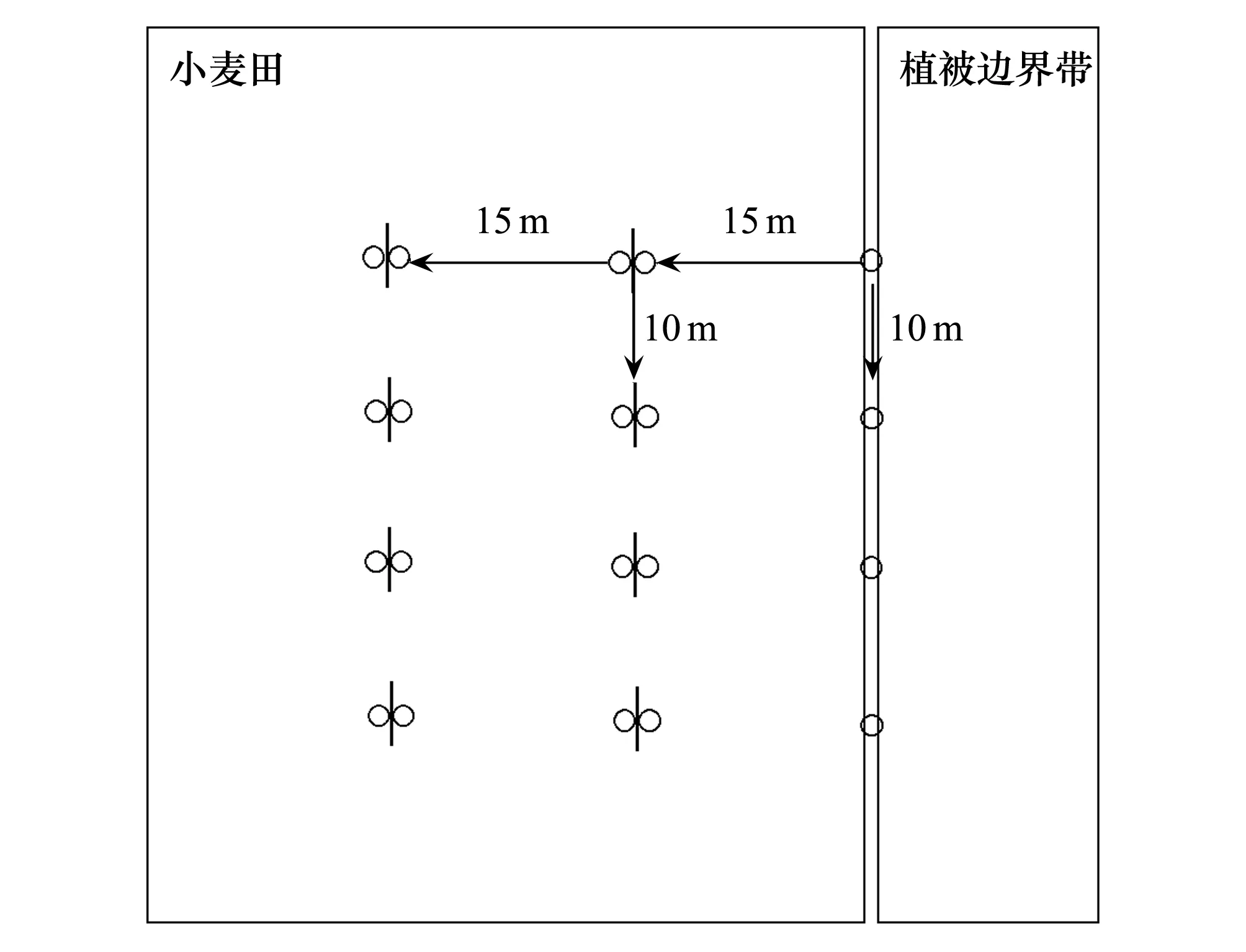
图2 样点布置图Fig.2 Layout of sampling sites
1.4 麦蚜调查
2014年5月7日至6月3日,研究区麦蚜增长期与高峰期,对样地麦田蚜虫进行调查。为保证调查充分,选取3条50 m长的平行样带,第一条距离农田边界0—1 m,第二条距离边界10—11 m,第三条距离边界20—21 m。每条样带,每隔5 m随机选取5株小麦,共调查20株,每株小麦随机选取5个茎秆,分别记录每株小麦上的蚜虫数量[11]。每隔5 d调查一次,共6次。
1.5 植被调查
植被边界带于2014年5月下旬进行植被调查。记录边界带宽度(BW)、样地植被总盖度(SC)、乔木层物种类型(TT)与盖度(TC)、草本层总盖度(HC),同时设置4个1 m×1 m的样方,记录样方内草本层的物种名称与相对盖度,合并4个样方内草本层物种信息,计算样地的物种丰富度(SR),并以以上6个指标作为植被边界带植被群落结构指标。
1.6 数据分析
为弥补取样期间陷阱破坏造成的取样不均,步甲和蜘蛛的多度数据采用其活动密度(个体数/天/杯)进行标准化。合并每个取样点农田内部20个陷阱数据用于分析不同类型植物边界带对麦田天敌发生的影响。采用单因素方差分析比较不同植被边界带麦田内部步甲和蜘蛛活动密度差异。采用基于CNESS(Chord-Normalized Expected Species Shared)相似系数的非度量多维尺度法(Non-metric Multi-Dimensional Scaling, NMDS)分析比较不同植被边界带相邻小麦田中步甲和蜘蛛群落结构的相似性。
不同类群地表天敌(步甲和蜘蛛)对小麦蚜虫的控制潜力采用益害比表示,合并小麦田不同取样带6次蚜虫调查数据作为样地蚜虫发生数据,采用取样地20个陷阱中步甲和蜘蛛多度的合并数据计算小麦田步甲蚜虫比、蜘蛛蚜虫比与天敌(步甲与蜘蛛总体多度)蚜虫比。采用单因素方差分析比较不同植被边界带对相邻小麦田中蚜虫发生以及益害比的影响。
边界带植被特征对相邻小麦田天敌群落结构的影响采用冗余分析( redundancy analysis, RDA)进行。首先对小麦田内步甲和蜘蛛的群落结构采用去趋势对应分析( detrended correspondence analysis, DCA),发现步甲和蜘蛛的群落结构呈线性分布,因而采用RDA分析[34]。取样点天敌群落通过合并整个取样季小麦田3个取样距离截面共计20个陷阱数据获得,半自然生境的特征通过植被边界带宽度、乔木种类、样地总盖度、乔木层植被盖度、草本层植被盖度以及物种丰富度6个指标来衡量。
步甲和蜘蛛在植被边界带与小麦田之间的溢出效应通过两个方面进行验证,一方面通过比较边界处与农田内部挡板两侧的天敌活动密度探讨步甲和蜘蛛的移动方向;另一方面通过分析距农田边界不同距离取样带步甲和蜘蛛的活动密度探讨地表天敌可能的扩散方向。分析采用一般线性回归,以小麦田不同距离取样带以及挡板两侧的步甲、蜘蛛活动密度作为响应变量,以植被边界带类型、距离农田边界取样距离以及挡板两侧方向作为固定效应变量,边界处作为第三类方向类型。由于挡板两侧的步甲和蜘蛛活动密度数据不符合正态分布,采用平方根(SQRT)转换使其符合方差正态分布后进行分析。
方差分析采用DPSv7.05[35]进行,多重比较方法采用Duncan新复极差多重比较方法。非度量多维尺度法(NMDS)分析采用PAST 3.14[36]。RDA 分析过程选用CANOCO5[34, 37]进行。一般线性回归采用SPSS Statistics 20[38]进行。
2 研究结果
2.1 天敌群落组成
2014年5月10日至6月9日,在研究区具有不同类型植被边界带的小麦田内共捕获步甲1094头(表1),分属于29个种。其中优势种为半亮虎步甲(Asaphidionsemiliucidion),占捕获步甲总个体数的的13.7%。常见种包括四斑小步甲(Tachysgradatus)、单齿蝼步甲(Scaritesterricola)、毛婪步甲(Harpalusgriseus)、棒婪步甲(Harpalusbungii)、杰氏狭步甲(Oxycentrusjelineki),分别占捕获步甲总个体数的12.0%、11.4%、9.9%、9.6%和9.3%。共捕获蜘蛛2229头,分属于14个科。其中优势科为狼蛛科,占捕获蜘蛛总个体数的58.4%,常见科为皿蛛科和平腹蛛科,分别占捕获蜘蛛总个体数的13.8%和13.5%。

表1 与不同类型植被边界带相邻的麦田中步甲和蜘蛛组成及分布
WW: 毗邻人工林地的小麦田,Wheat field neighbored by woodland; AW: 毗邻人工种植草带的小麦田,Wheat field neighbored by plant grass strip; NW: 毗邻自然演替草带的小麦田,Wheat field neighbored by succession grass strip
2.2 植被边界带类型对相邻麦田步甲、蜘蛛活动密度及群落结构的影响
具有不同植被边界带的小麦田中步甲、蜘蛛活动密度方差分析显示,人工林地边界带相邻的小麦田内步甲、蜘蛛活动密度显著高于另外两种类型(步甲:F=0.732,P=0.014; 蜘蛛:F=46.638,P<0.001),而与自然演替草带以及人工种植草带相邻的小麦田内步甲和蜘蛛的活动密度相互之间无显著性差异(图3)。

图3 不同类型生境步甲、蜘蛛活动密度方差分析Fig.3 ANOVA of carabid and spider activity density in different habitats
对具有不同植被边界带的小麦田步甲和蜘蛛群落结构分析显示,就步甲而言,除与人工种植草带相邻的小麦田中步甲群落显示出较高的相似性之外,其他两种边界类型的小麦田中步甲群落较为分散,然而总体看来,与自然演替草带相邻的小麦田主要分布在排序轴的左上侧,而与人工林地相邻的小麦田主要分布在排序轴的右下侧(图4);而蜘蛛群落则表现为,与林地相邻的小麦田高度聚集在一起,蜘蛛群落表现为高度的均质性,与其他两种类型边界相邻的小麦田分布高度分散,麦田内蜘蛛群落组成表现为高度的异质性(图4)。
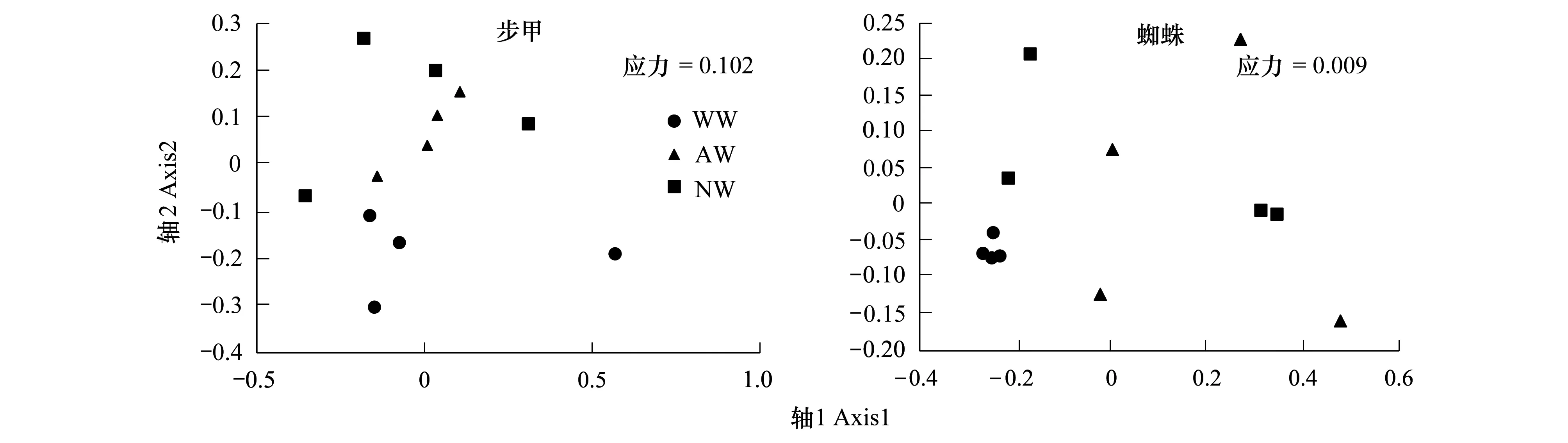
图4 不同边界类型麦田步甲、蜘蛛群落非度量多维度分析(NMDS)Fig.4 Non-linear multi-dimensional scaling (NMDS) based on Chord Measure for natural enemy assemblages at wheat fields with different field margins
2.3 植被边界带对相邻麦田蚜虫分布及益害比的影响
与不同类型植被边界带相邻的小麦田中蚜虫多度方差分析显示,人工林地相邻的小麦田中蚜虫个体多度显著高于与其他两种类型麦田(图5,F=4.903,P=0.036)。然而,具有不同类型植被边界的小麦田相互之间,益害比无显著性差异(图5, 步甲/蚜虫:F=1.210,P=0.342, 蜘蛛/蚜虫:F=2.670,P=0.123;(步甲+蜘蛛)/蚜虫:F=2.045,P=0.185)。

图5 不同边界类型小麦田蚜虫及益害比方差分析Fig.5 ANOVA of aphid and enemy-aphid ration in wheat fields with different field margins
2.4 农田边界特征对相邻麦田步甲、蜘蛛群落分布的影响
Monte Carlo 检验结果显示,不论步甲还是蜘蛛,其群落结构分布都受植被边界带宽度以及乔木层盖度的影响显著(表2)。就步甲而言,RDA排序图前两轴共计解释了步甲群落结构变异的31.3%,边界宽度和乔木盖度共计解释前两轴解释率的43.5%。而就蜘蛛而言,RDA排序图前两轴共计解释了蜘蛛群落结构变异的45.9%,边界宽度和乔木盖度则解释了前两轴解释率的71.8%。
步甲RDA排序前两轴特征根植之和为0.313。RDA排序图第一轴解释了物种环境关系的23.2%,乔木层盖度和植被边界带宽度都在第一轴有较高的正得分;第二轴显示物种环境累积贡献率为8.1%。RDA排序图显示(图6),与林地相邻的小麦田都分布在第一排序轴右侧,与乔木层盖度以及植被边界带宽度显著正相关;与人工种植草带相邻的小麦田主要分布于第二象限;与自然演替草带相邻的小麦都分布于第三象限。步甲优势种半亮虎步甲(SP5)与常见种四班小步甲(SP28)、微婪步甲(Harpalusmicrodemas)(SP19)主要分布于与人工林地相邻的小麦田中,与乔木盖度以及植被边界带宽度呈正相关关系。常见种单齿蝼步甲(SP25)则主要分布在第一轴左侧,与乔木层盖度及植被边界带宽度呈负相关关系;常见种毛婪步甲(SP18)、杰氏狭步甲(SP22)与棒婪步甲(SP14)则主要分布于无乔木层覆盖的自然演替草带相邻的小麦田中。而稀有种如Agonumsp.(SP1)、短胸暗步甲(Amarabrevicollis)(SP2)以及强婪步甲(Harpaluscrates)(SP17)则出现在与人工种植草带相邻的小麦田中。
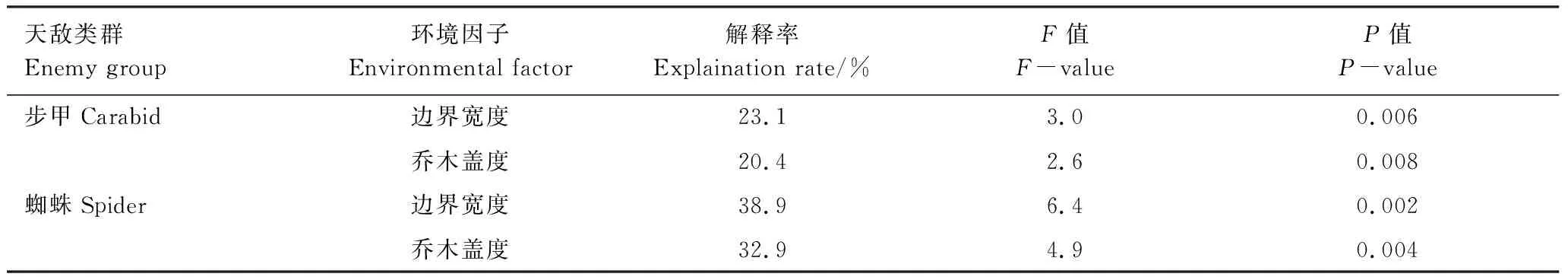
表2 RDA 模型蒙特卡洛检验具有显著性的环境因子
蜘蛛RDA排序前两轴特征根值之和为0.459。RDA排序图第一轴物种环境累积贡献率为39.0%,乔木层盖度和植被边界带宽度都在第一轴有较高的正得分;第二轴显示物种环境累积贡献率为6.9%。RDA排序图显示(图6),优势科狼蛛科(Fa8)与常见科平腹蛛科(Fa4)分布于第一轴右侧,与乔木层盖度以及植被边界带宽度呈正相关关系。而常见科皿蛛科(Fa6)则主要出现在与人工种植草带相邻的小麦田中。

图6 边界带植被群落结构与相邻麦田中步甲、蜘蛛群落RDA分析Fig.6 Canonical correlation analysis (RDA) ordination plot of carabid and spider assemblages and flora structure of field marginsTC: 乔木层盖度,Tree coverage; BW:边界宽度, Border width
2.5 步甲和蜘蛛的溢出效应
综合比较与不同类型植被边界带相邻的小麦田边界0 m处与距离边界15 m处及30 m处挡板内外两侧步甲和蜘蛛的活动密度,结果显示(表3),就步甲而言,其活动密度空间分布仅受相邻植被边界带类型影响(F=9.347,P=0.010),与农田植被边界带的取样距离以及不同取样距离挡板两侧与边界处相互之间步甲的活动密度均无显著性差异(取样距离:F=0.287,P=0.595; 挡板方向:F=0.096,P=0.785),且植被边界带类型、取样距离与挡板方向相互之间对步甲活动密度无交互作用。
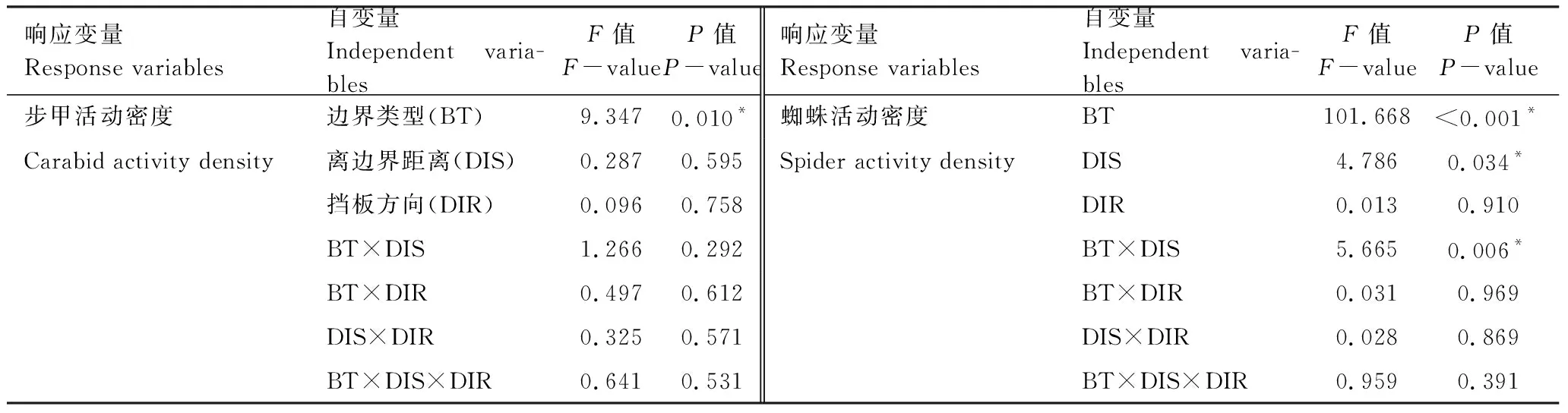
表3 植被边界带类型、取样距离以及挡板方向对步甲和蜘蛛活动密度影响的一般线性模型
BT:边界类型,Border type; DIS:离边界距离,Distance; DIR:挡板方向,Direction
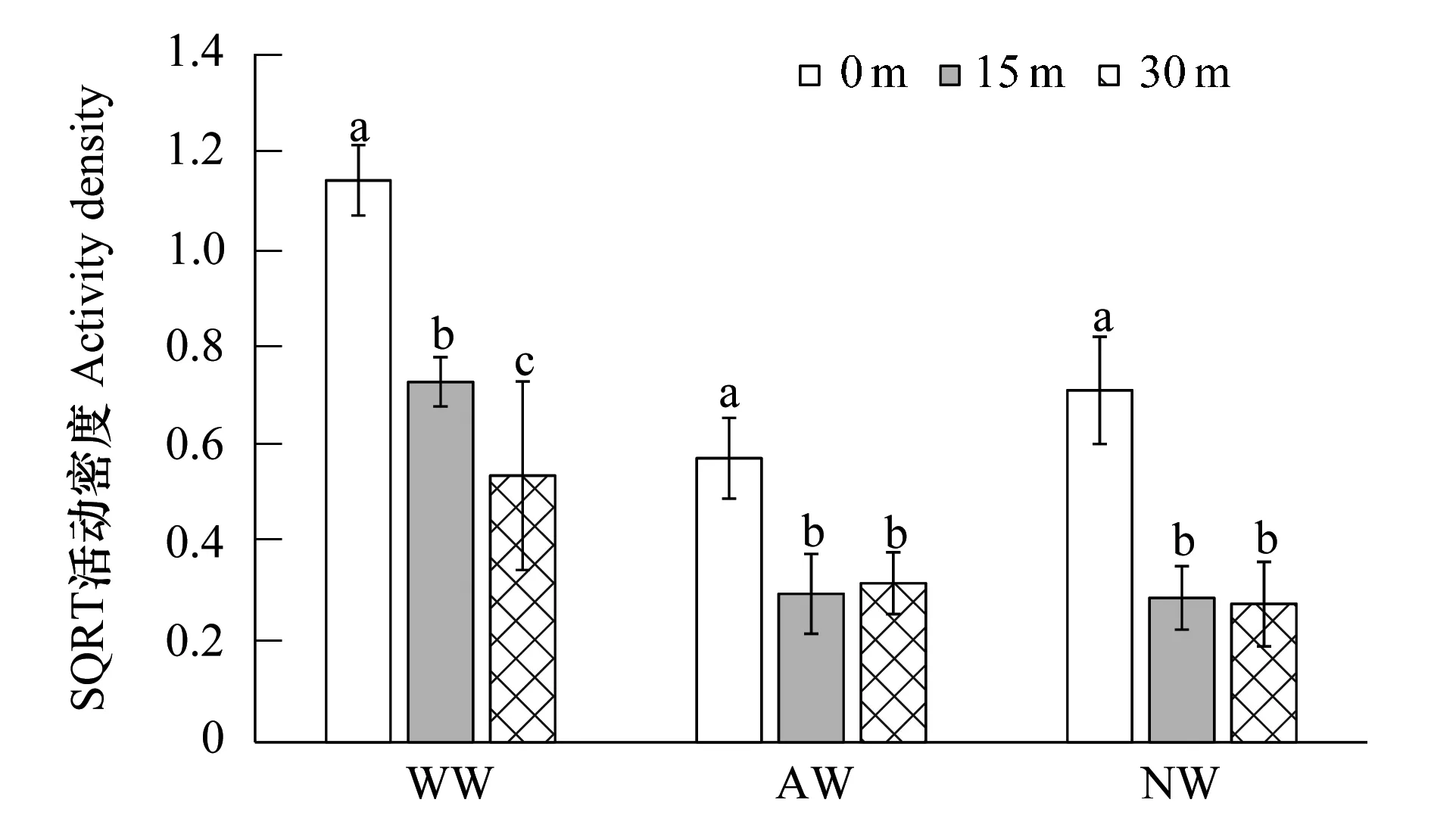
图7 不同类型植被边界带小麦中不同取样距离处蜘蛛活动密度Fig.7 Activity density of spider at different sampling distance in wheat field with different vegetation borderSQRT:平方根转换,Square root
就蜘蛛而言(表3),其活动密度不受挡板两侧方向的影响(F=0.013,P=0.910),植被边界带类型和距边界带取样距离共同决定蜘蛛的活动密度(边界类型:F=101.668,P<0.001; 离边界距离:F=4.786,P=0.034; 交互作用:F=5.665;P=0.006)。具体表现为,与人工林地相邻的小麦田表现为,从农田边界0 m处到农田内部15 m处,再到农田内部30 m处,蜘蛛的活动密度依次呈显著性降低趋势;而与自然演替草带以及人工种植草带相邻的小麦田中,农田边界0 m处蜘蛛的活动密度显著高于农田内部15 m及30 m处,而农田内部两条取样带之间,蜘蛛活动密度无显著性差异(图7)。
3 讨论
3.1 植被边界带对相邻麦田中步甲、蜘蛛分布的影响
另一方面,尽管与人工种植草带及自然演替草带相邻的小麦田中维持了与林地相邻小麦田相对较少的步甲、蜘蛛活动密度,然而,物种群落结构排序分析显示,与人工种植草带相邻的小麦田中维持了较为稳定且区别于与毗邻人工林地小麦田中的步甲群落结构以及异质性较高的蜘蛛群落结构;同时,与自然演替草带相邻的小麦田中维持了异质性较高的步甲和蜘蛛群落结构,因此,此两种类型的植被边界带对于提高农业景观步甲和蜘蛛的Beta多样性具有十分重要的作用。早期的研究显示,边界草带对于提高相邻农田中步甲类天敌的多样性具有重要作用[43-44],欧美国家很早就将农田草带建设作为保护农田甲虫多样性的重要农业生态工程技术加以推广和利用[45- 47]。此外,作为研究区常见种毛婪步甲、杰氏狭步甲与棒婪步甲的主要生境,自然演替草带对于维持农业景观步甲群落物种库组成起到了重要作用;而人工种植草带则对维持步甲稀有种如Agonumsp.、短胸暗步甲以及强婪步甲具有重要作用;同样,蜘蛛常见科皿蛛科主要出现在与人工种植草带相邻的小麦田中,这可以应用于农业景观管理过程中,通过人工建设缓冲带有针对性的提高步甲物种多样性以及维持蜘蛛群落稳定性。
需要指出的是,以往的研究通常将不同类型的半自然景观要素笼统的合并为同一类来探讨其对农业景观步甲和蜘蛛多样性的影响[48-49],然而,本研究显示不同类型半自然植被边界带,如人工林地、半自然草带,维持了具有显著差异的步甲和蜘蛛群落结构,因此,在进行研究和景观管理过程中,应当充分考虑不同类型半自然景观要素影响和作用的差异[8, 50]。
3.2 植被边界带对相邻麦田中蚜虫的发生以及益害比的影响
蚜虫多度空间分布方差分析显示,与人工林地相邻的小麦田中具有显著高于其他两类小麦田的蚜虫个体数。这与我们最初认为的,与林地相邻的小麦田中高步甲和蜘蛛多度可能将蚜虫密度控制在较低水平相反,一方面,周围景观中半自然植被在提高天敌多样性的同时,可能也为害虫提供了相应的资源及适宜的微环境,导致害虫数量大量增加[9];另一方面,半自然生境复杂的植被群落结构可能为害虫躲避天敌的捕食提供屏障,导致天敌捕食害虫的难度增加,害虫捕获效率下降[51]。事实上,可能是小麦田中较高的蚜虫密度促进了天敌的多样性。Martin等[21]的研究也得出具有高步甲和蜘蛛多度的生境中往往具有较高植食作用,并指出景观中害虫群落的增殖较天敌增殖发生的更早,也更迅速。而农田中高密度的食物正是天敌群落迅速增殖的重要原因。此外,不同类群天敌对于不同类群害虫的控制效率存在较大差异[17, 21],且捕食者内部可能存在较为强烈的竞争或者内捕食作用[23, 52]。因此,在研究半自然生境生物控制服务功能时,不能仅仅考虑天敌和害虫的发生数量,需要综合考虑益害比和害虫的取食作用[17, 21]。
3.3 步甲和蜘蛛的溢出效应
与张旭珠等[53]2014年在曲周地区的研究结果相同,距农田边界不同取样界面挡板两侧步甲和蜘蛛的活动密度同样没有显著性差异,这可能是由于单个步甲、蜘蛛个体的实时移动可能无特定规律[54],并非有目的性的从边界向农田内部移动。然而,植被边界带与小麦田边界处维持了较高的蜘蛛活动密度,并且具有向农田内部溢出的倾向,这对于蚜虫爆发时期,提高蚜虫的生物控制效率具有重要作用[55-56]。早期的研究指出,多数节肢动物天敌,即使是适应农田生境的农田物种,如某些步甲和蜘蛛等[57- 59],仅仅使用农田植被边界带作为越冬场所,随后便向农田中进行扩散[15]。此外,天敌的扩散距离与不同类群天敌的扩散能力有关。本研究中,与林地相邻的小麦田中蜘蛛群落主要为狼蛛科和平腹蛛科,此两个科蜘蛛都属于游猎型蜘蛛,主要通过地表爬行进行扩散[12],因而,可通过陷阱法捕获的样本中发现其从农田边界向农田内部扩散的趋势。而与人工种植草带以及与自然演替草带相邻的小麦田中,除狼蛛科外,结网型捕食的皿蛛科同样占有较大比例。皿蛛科通过气流在空中进行传播[12],一方面通过陷阱法捕获的蜘蛛类群不能够完全代表取样截面全部的皿蛛科类群,另一方面,其扩散距离与扩散方向具有明显的不确定性,可能远远大于30 m,因此,在与人工种植草带相邻以及与自然演替草带相邻的小麦田中,仅观测到边界处具有显著较高的蜘蛛活动密度,并无发现其沿取样距离梯度依次递减的趋势。
而就步甲而言,距离农田边界不同取样截面处,步甲的活动密度无显著性差异,这可能与扩散源中天敌库密度有关,由于研究区农业生产的悠久历史及深远影响,能够存活至今的往往是那些适应性强或适应于农田生境的物种[60]。研究显示,农业景观中存在着大量的步甲农田特有种,这些物种主要以农田作为其生境,仅在农田环境条件恶劣或越冬时向半自然生境转移[15,57]。张旭珠等[53]2014年在曲周地区的研究同样得到与防护林相邻的玉米田中,农田边界处与距离农田边界10 m处,步甲的活动密度无显著性差异。此外,早期研究证明作物生境与植被边界带之间存在着明显的边界效应,生境界面特征以及农田中作物密度、空间开放度也是影响天敌扩散难易程度的重要因素[61-62]。赵紫华等[10]认为,麦田与不同类型植被边界带相邻的生境界面上,边界效应所导致的物种流动情况具有显著不同。因此,尽管农田草带被认为可以维持大量的步甲多样性[29-30],但也可能由于边界效应,导致草带中的步甲不能向农田流动。
4 结论
作为研究区残留的主要半自然生境类型,人工林地对于相邻小麦田步甲和蜘蛛多样性的维持具有重要作用。然而,并非高天敌密度就意味着低害虫发生率,未来的研究需进一步探讨天敌多样性与害虫发生关系及其影响因子。在步甲和蜘蛛群落结构方面,尽管与人工种植草地以及与自然演替草带相邻的小麦田中维持了相对与人工林地相邻小麦田较少的步甲和蜘蛛活动密度,但却各自维持了相对独特的物种群落结构,对于增加景观中步甲和蜘蛛群落的Beta多样性具有重要作用。因此,不同类型半自然生境对天敌多样性的维持和保护、害虫控制作用不同,在通过景观管理措施促进天敌多样性保护和害虫控制过程中,需要考虑不同半自然生境的作用,并充分维持半自然生境的多样性。植被边界带的宽度以及乔木层盖度是影响相邻小麦田步甲和蜘蛛活动密度的主要因素,因此建立具有复杂垂直群落结构的植被边界带是农业景观建设的一项重要措施。此外,由于不同类群天敌的生物学特性有所差异,目前对于步甲和蜘蛛的在不同生境界面间的溢出效应机制仍未达成统一结论,未来的研究应进一步探讨不同类型生境界面地表天敌的活动规律。
