稀土尾矿构树根系形态及代谢研究
2019-01-14郭圣茂吴南生肖永华彭明生
丁 菲,郭圣茂,吴南生,肖永华,彭明生,蒋 礼
(1.江西农业大学职师院,江西南昌330045;2.江西省宁都县林业局,江西宁都342800;3.中储牧草科技有限公司构树饲料研究中心,河南兰考475300)
赣南拥有全国30%以上的中重稀土储备,有着“稀土王国”之称,但落后的开采工艺剥离了山体表层植被,造成了灾难性的生态环境问题,赣南稀土废弃矿山的治理迫在眉睫。尾矿土壤营养匮乏、持水性差、含砂量高的极端环境特点使植物很难在此存活,土壤修复极其困难,人工干预才能加快植被恢复进程[1]。构树(Broussonetiapapyrifera)是桑科构树属落叶大乔木,适应性特别强,民间称其为“恶树”,近年来逐渐受到众多研究者的关注。一些学者应用构树对锑矿、采石矿、铅锌矿、菱镁矿等矿区进行植被修复,认为其是矿区废弃地生态恢复的适生先锋树种[2-3],本课题组前期研究工作也证明构树对赣南稀土尾矿具有较强的耐受性[4]。文献表明,关于构树对矿区的适应性和对旱盐、低温、重金属等胁迫的抗逆性研究主要反映在地上部生长和生理生化响应[5-7],对构树地下生态系统与抗逆关系的研究极少。根系是地下生态系统中进行物质和能量交换的主要器官,由于直接与土壤接触,更容易对土壤环境做出反应[8],受生境差异影响,植物根系会在空间分布、形态特征、生理特性上表现出不同的生态适应对策。许多学者研究了根系形态对逆境的响应,Hund等[9]和李文饶等[10]认为干旱胁迫下,根系生长受到抑制,根系生物量、根冠比、根长、根系表面积和体积等发生变化以适应干旱胁迫,王树凤等[11]研究发现Cd胁迫抑制了杞柳根的伸长,促进了根的径向生长,导致根系平均直径增加。根系适应逆境的生理响应报道也较多[12],王亚男等[13]认为鸢尾根系代谢分析有利于更好地理解植物对石油烃污染土壤的代谢响应并揭示其修复机制。代谢组学是在现代高通量检测技术平台不断发展的基础上,以生物体内小分子代谢物分析与数据统计分析为基础,通过生物体内代谢物的种类、数量及其代谢途径的变化来研究生物体的代谢过程,已经发展成为探索植物对遗传和环境的改变所做的系统响应的重要工具[14-15]。
综上,构树有望成为矿区等困难立地生态重建的先锋树种,然而构树根系在生态恢复中的作用及其地下关键生态过程对矿区干旱、贫瘠逆境的响应机制长期被忽视,从地下生态系统角度进一步探讨构树的逆境适应策略具有重要的理论和现实意义。本研究通过分析构树根系的形态特征,应用代谢组学研究根系代谢产物的变化特点,拟揭示构树适应稀土尾矿土壤环境的内在机制,发展和完善构树抗性理论,为尾矿废弃地的生态恢复提供理论支撑。
1 材料与方法
1.1 研究区概况
试验地点位于江西省赣南地区宁都县东山坝镇,东经116°31′,北纬26°44′,地处中亚热带丘陵山区季风湿润气候区,四季分明,气候温和,平均气温在摄氏19.8 ℃度左右,全年无霜期近11个月,年降雨量1 600 mm左右,多集中在4—6月份,约占总降雨量的50%。土壤为稀土矿开采4年后的废弃残矿尾砂,开采后的山头有效土层和风化层基本剥离,水土流失严重,使得土壤沙化,沙砾含量高,保水性能非常差,养分含量极低,寸草不生。由于稀土洗矿工艺和水土流失造成试验地沟蚀严重,在造林之前用挖机对废弃尾矿地进行了充分平整,2015年3月,选择长势基本一致的1 a构树实生苗对稀土尾矿进行造林,平均株高51 cm,平均地径1.1 cm,造林密度为1.0 m×0.5 m,同时在同一山头周边未开采的土壤种植构树作为对照。
1.2 研究方法
1.2.1 根系形态研究采样 2017年7月设置试验样地(稀土尾矿区,以Y表示)和对照样地(周边未开采的坡地,以CK表示)各4块,样地面积为10 m×20 m,每块样地各选4株标准木共16株,采用挖掘法,先挖开幼苗周边表层土,再沿着根系分布小心挖掘,同时用水冲洗根部,使其露出土壤,以保护侧根不脱落。选取根系完整的植株装入塑料袋,编号装入密封的塑料袋,带回实验室测量根系形态和生物量[16]。将每株主根上的全部侧根剪下,用Epson Expression 00XL扫描仪获取根系扫描图像,用Win RHIZO根系分析软件(加拿大Regent公司)自动统计不同直径总根长度、总根表面积,扫描后的根系样品置入烘箱80 ℃烘干24 h至恒质量,再分别称质量和记录。比根长(specificrootlength,SRL)为根系总根长与根系生物量之比,比表面积(specificrootarea,SRA)为根表面积与根系生物量之比。
1.2.2 代谢物分析取样和检测 在试验和对照样地中分别随机挖取30株侧根根系,5株混合为一个生物学样品,每组6个重复,立即用液氮速冻用于代谢组分析。取样品100 mg用液氮研磨,取100 μL样本,加入300 μL甲醇和20 μL内标,涡旋混匀30 s后,冰水浴中超声提取5 min,-20 ℃静置2 h,4 ℃ 13 000 rpm离心15 min,取200 μL上清液于2 mL进样瓶,进行LC-MS分析。质控样本(QC)由实验样本提取物等量混合制备而成,用于分析样本在相同的处理方法下的重复性。在仪器分析的过程中,每6~10个检测分析样本中插入一个质控样本,以监测分析过程的重复性。LC-MS分析的仪器平台由Agilent1290超高效液相色谱串联Thermo Fisher Scientific 的Q Exactive Orbitrap高分辨质谱仪组成。所用色谱柱为UPLC HSS T3色谱柱(1.7 μm 2.1×100 mm,Waters)。正模式流动相为0.1%甲酸水溶液和乙腈,负模式流动相为5 mM醋酸铵水溶液(用氨水调节pH值至9.00)和乙腈,采用Q Exactive Orbitrap高分辨质谱仪进行一级、二级质谱数据的采集。
1.2.3 数据处理 根系形态所有数据利用Microsoft Excel和SPSS软件进行处理,同一径级不同处理之间、同一处理不同径级之间根长、根表面积、生物量、SRL、SRA的差异采用独立样本T检验。显著性水平设定为P=0.05。对质谱分析后得到的原始数据进行基线过滤、峰识别、积分、保留时间校正、峰对齐和归一化,最终得到一个保留时间、质荷比和峰强度的数据矩阵,利用归一化后的数据矩阵进行多元统计分析,结合OPLS-DA(正交偏最小二乘法判别分析)的VIP值和单变量统计分析T检验P值来筛选显著差异代谢物并画热图。
2 结果与分析
2.1 构树修复稀土尾矿的根系形态响应
2.1.1 细根根长、表面积和生物量 从图1可以看出,与周边未开采矿区构树相比,稀土尾矿区构树细根总长下降了32.39%、总表面积下降了21.69%,根系生物量下降了52.6%。除>2 mm径级外,同一径级根系长度、表面积和生物量均呈显著减少。进一步分析表明,尾矿区构树<1 mm径级的根长占总根长的84.01%,比对照区同径级根长占比增加10.05%,而>2 mm细根长度所占整个根系的百分比例呈减少的趋势。尾矿区构树根系<1 mm径级的表面积占比也出现类似情况。
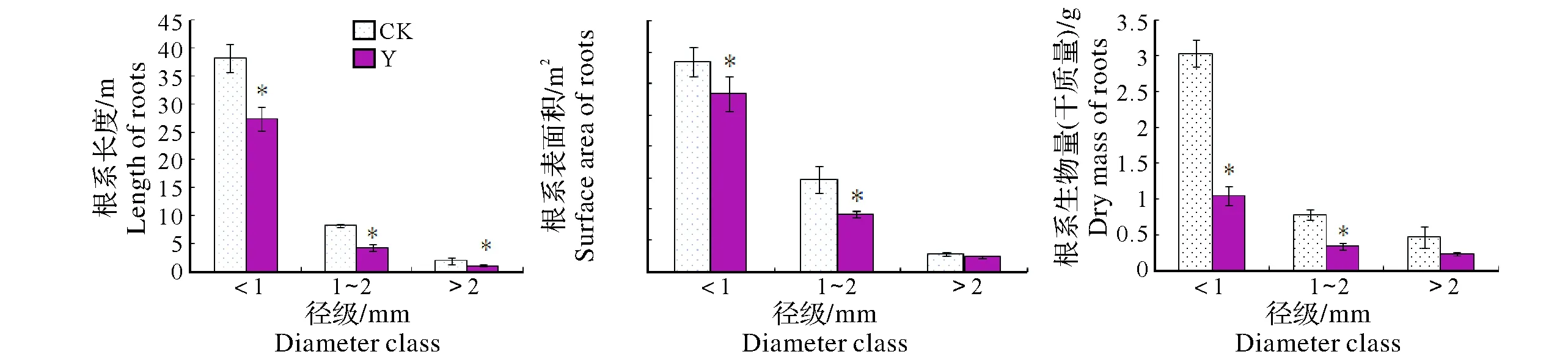
Y,尾矿区Rare earth tailing area;CK,周边未采矿区unmined area around earth tailing柱形图上方*的差异表示在5%水平上差异显著Values in each column followed by the * are significantly different at 0.05 level according to LSD图1 构树不同径级根系长度、表面积和生物量Fig.1 Root length,surface area and dry mass of different diameter class in B. papyrifera
2.1.2 比根长、比表面积 由图2可知,尾矿构树总的比根长表现出增加的变化趋势,其中Ø<1 mm的比根长比对照增加109.58%,1 mm<Ø<2 mm的比根长增加19.55%,总比表面积和各径级根比表面积分别比对照增加81.91%,152.09%,50.61%,55.26%,最小径级的比表面积增幅最大,分析发现,小径级的根系的比根长和总比表面积占总比根长和总比表面积的比例也较大。尾矿构树直径大于2 mm的根系比对照增加但未达显著水平,除此之外,尾矿土壤环境对构树各径级的比根长、比表面积的影响均达到显著水平。

图2 构树不同径级根系比根长、比表面积Fig.2 Special root length and Specific surface area of different diameter class in B. papyrifera
2.2 构树修复稀土尾矿的根系代谢分析

图3 构树处理(Y)与对照样品(CK)的PCA得分图Fig.3 PCA scores of treated(Y) and control (CK) in B. papyrifera
2.2.1 构树修复稀土尾矿的根系PCA和离子流分析 从图3的PCA得分图可以看出,两组样品6个重复的数据点检测结果都分别集中在一起,说明整个分析测试过程中系统误差(实验操作、仪器稳定性)较小,数据可重复性好。另外,两组样品的数据点在空间上可以明显区分,说明它们的代谢产物在数量、种类和浓度等方面存在差异,这种差异在总离子流图(图4)中也得到充分体现:如图4,检测到2.8~3.5 min,3.6~4.2 min,6.7~8.2 min几个阶段稀土尾矿构树根系中大量小分子物质显著上调,而在8.2~9.2 min,10.5~11.2 min阶段许多代谢物质与对照相比呈显著下调趋势。经过LC-MS分析,在处理组(Y)和对照组(CK)样品中共检测出1 968种代谢物存在显著差异(VIP≥1且T-testP<0.05),其中上调物质1 812种,下调物质156种。
2.2.2 稀土尾矿构树根系差异代谢产物分析 如图5所示,与周边未采矿坡地构树相比,稀土尾矿构树根系代谢物中,亚油酸、苹果酸、木二糖、葡萄糖、叶黄素、松三糖、半乳糖醇、色氨酸、酪氨酸、D-泛酸、葫芦巴碱、乳糖、麦芽酚、白藜芦醇等38种物质含量显著增加,而减少了丝氨酸、十六烷酰胺、硬脂酰胺等8种物质的含量,大部分的代谢物是在初生代谢途径中产生的,包括有机酸、氨基酸、糖类及其它组分。
为了进一步分析这些差异物质在稀土尾矿构树根系代谢中的贡献,对VIP值进行计算,如图6所示,发现葫芦巴碱、棕榈酸、谷胱甘肽醇VIP>10,其次为海藻糖、3-羟基-L-谷氨酸、亚油酸、麦芽糖、色氨酸、1,7-二甲基黄嘌呤、3-脱氢莽草酸酯等,它们在应对稀土尾矿环境中起到了实质性的作用。

图4 构树处理(Y)与对照样品(CK)的总离子流图Fig.4 Total ion chromatography of treated(Y) and control(CK) in B. papyrifera
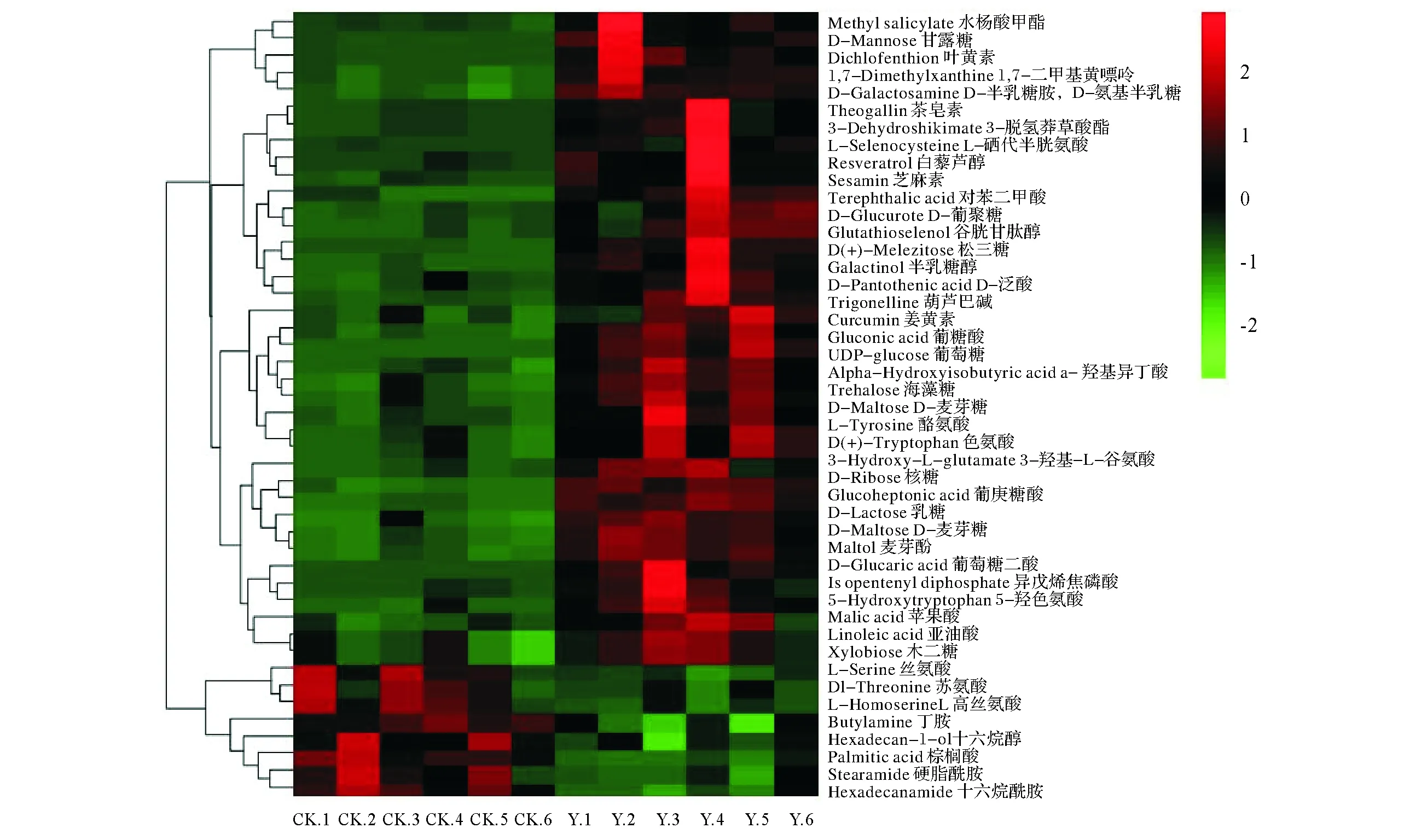
图5 构树处理(Y)与对照样品(CK)的差异代谢物热图Fig.5 Differential metabolite Heatmap of treated (Y) and control (CK) in B. papyrifera
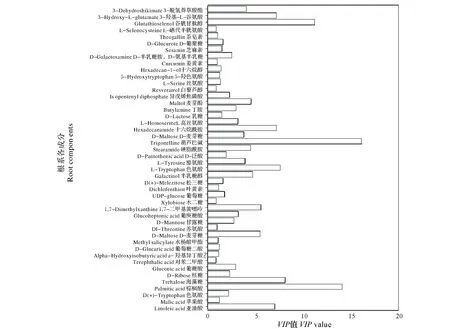
图6 构树根系差异代谢物的VIP值Fig.6 VIP of different metabolites in B. papyrifera
3 讨论与结论
3.1 稀土尾矿构树根系形态适应特征
根系生长关系到植物的水分和养分吸收,遭受胁迫时可以通过形态、生理方面的变化来适应环境[17-18],表征根系形态特征的参数主要有根系总长度、表面积、生物量、平均直径、比根长、比根面积等。本试验地土壤沙化干旱、营养匮乏,是稀土尾矿植被恢复的主要障碍因子[19]。研究发现,与周边未开采矿区构树相比,稀土尾矿土壤环境使构树根系的形态特征发生了变化,但各形态指标变化不一致。
一方面,侧根长度、表面积和生物量明显减少,可能主要是由于尾矿土壤贫瘠、含水量低,限制了根系的生长,这与Pregitzer等[20]研究结论一致。但是低序级侧根根长和表面积占比提高。研究表明,水和无机矿物质的吸收通过较细的根进行[21-22],直径越小的根吸收养分和水分的能力越强[23]。吴敏等[24]发现,栓皮栎幼苗的小细根可以帮助根系在土壤中占据更多的空间位置,增强根系对周围水分的吸收能力。因此,构树在尾矿干旱贫瘠的环境下根系形态的这种变化可以扩大横向吸收水分和养分的面积,是根系对资源匮乏生境的生态适应。
另一方面,尾矿构树总比根长、总比表面积比对照显著增加,小径级根系比根长和比表面积增加幅度较大且差异显著。比根长主要表征植物根系收益和投入的关系,比根长增加,单位质量的根系具有更大的吸收表面积,土壤资源的获得效率更高。武高林等[25]对4种凤毛菊属(Saussurea)植物研究后发现,植物在恶劣条件下能够通过增加比根长来更有效地吸收土壤中的养分。比表面积反映植物根系对土壤资源获取的能力[26],Bauhus等[27]认为,SRL和SRA决定了根系吸收水分和养分的能力,其值越大,表明投入相同的成本有较高的养分利用效率。本研究SRL和SRA的变化表明构树根系能够以较少的投入获得更多的收益,水分和养分利用效率较高,说明构树不是被动忍受而是主动调节其根系生长以提高吸收效率减缓逆境伤害。
综合前述,应该是由于稀土尾矿严重缺水缺肥的恶劣土壤环境条件诱导构树产生了更多数量、更小直径的细根,而构建和维持单位长度细根消耗的碳水化合物较少,有利于行使细根的吸收功能[28],这可能是构树应对稀土尾矿环境的根系调整策略。
3.2 稀土尾矿构树根系代谢响应
植物对非生物胁迫的耐受性与敏感性是一个非常复杂的生命过程,大量物质含量改变,积累渗透保护剂、调整启动抗氧化酶保护系统、激活相关保护性的代谢途径以重构代谢平衡[29]。本研究中检测到大量差异代谢物质,主要包括氨基酸、糖类、有机酸等。
氨基酸在植物应对逆境生理中扮演着重要的角色,大量研究证明,非生物胁迫下植物体内游离氨基酸呈积累趋势[30],它们应对胁迫的重要职能是参与渗透调节[31-32],其中某些特定氨基酸的积累则可能预示着植物耐胁迫能力得到增强的积极信号[33]。本研究中分析表明稀土尾矿构树根系色氨酸、酪氨酸显著上调,分别富集到多种氨基酸代谢通路,参与了应对环境压力的调节,与前人的研究结论类似。
糖为植物生长发育和代谢过程提供碳和能量,增强植物抗逆性,在进化早期即获得抵抗胁迫重要调节功能。本研究发现,构树在稀土尾矿环境压力下,松三糖、核糖、麦芽糖、海藻糖、木二糖、葡萄糖、甘露糖含量显著增加,它们可能参与渗透调节,增强了构树适应稀土尾矿环境的能力。其中海藻糖VIP值达7.8,在所有糖类差异代谢物中表现最为突出。海藻糖不同于其它糖类,它对生物体具有神奇的保护作用,是应激代谢的重要产物。它能够保护生物细胞和生物活性物质在脱水、干旱、高温、冷冻、高渗透压及有毒试剂等不良环境条件下活性免遭破坏。许多对外界恶劣环境表现出非凡抗逆耐受力的物种,都与它们体内存在大量的海藻糖有直接的关系[34]。海藻糖与构树抗逆性的关系值得进一步深入研究。
在构树根系差异代谢物中,葫芦巴碱( trigonel-line)的VIP值最大。葫芦巴碱是一种嘧啶甜菜碱,是细胞质中的亲和性溶质,是一种较好的渗透调节物质,对细胞膜的稳定性起到重要的保护作用[35]。Cho等[36]对17个大豆品种进行盐胁迫处理,其中抗盐品种叶片中葫芦巴碱含量增加1.5倍以上。除此之外,葫芦巴碱还被认为是一种天然的植物激素而引起生物学者的关注,因为其能使植物根尖组织细胞周期的G2期停止或减慢G2期,Mazzuca等[37]以莴苣(LactucasativaL.)的根尖组织作为试验材料,用3mmol/L葫芦巴碱处理根尖组织,减小了原生根系的伸长速率。根据前述根系形态的变化,推测葫芦巴碱可能影响了稀土尾矿构树根系的生长和形态特征。所以,类似于前人的研究成果,葫芦巴碱作为构树根系代谢的非常重要的差异代谢物对稀土尾矿土壤环境做出响应,可能一方面作为渗透调节物质保护着细胞膜,另一方面调控着根系的生长。
当植物处于逆境条件时,体内的活性氧产生和清除的动态平衡遭到破坏,近年来人们对于植物活性氧氧化伤害机制和其防御体系做了大量研究,发现抗坏血酸、类胡萝卜素等许多抗氧化物质可以通过多条途径直接或间接地猝灭活性氧[38]。本研究表明,稀土尾矿构树根系中3-脱氢莽草酸酯、谷胱甘肽、泛酸、白藜芦醇、芝麻素、胡萝卜素等差异代谢物显著上调,研究表明,3-脱氢莽草酸酯是一种十分有效的抗氧化剂,其活性优于没食子酸、丙基没食子酸[39];泛酸在体内主要以CoA形式参与糖、脂、蛋白质代谢,具有抗脂质过氧化作用,保护细胞质膜不受损害;GSH不仅可以作为GR的底物通过Halliwell-Asada途径清除H2O2,也可以作为抗氧化剂直接清除活性氧,是一类重要的抗氧化物质;白藜芦醇、胡萝卜素也是强抗氧化剂[40],可见,这些差异代谢物都是重要的活性氧清除剂和抗氧化剂。
本研究分析了稀土尾矿构树的根系形态和根系代谢响应,结果表明Ø<1 mm的细根比例和细根表面积占比显著提高,总比根长、总比表面积显著增加且小径级根系SRL和SRA增加幅度较大,这种形态上的主动调节可以减少生长消耗,提高水分和养分吸收效率;根系代谢组学分析表明渗透调节作用和抗氧化作用是稀土尾矿构树适应稀土尾矿环境的主要途径,根系色氨酸、酪氨酸、苏氨酸、丝氨酸等氨基酸物质,松三糖、核糖、麦芽糖、海藻糖、木二糖、葡萄糖、甘露糖等糖类物质及葫芦巴碱等差异代谢物是重要的渗透调节物质,3-脱氢莽草酸酯、谷胱甘肽、泛酸、白藜芦醇、芝麻素、胡萝卜素等差异代谢物显著上调,作为抗氧化物质,增强了构树应对环境压力的能力。根系形态上的调整和根系代谢的差异是构树适应稀土尾矿环境的重要机制。
