Cep164基因缺失对小鼠胚胎早期脑发育的影响
2019-01-11王春花

摘要:利用Foxg1、Gbx2、Otx2、Fgf8和En1探针与小鼠胚胎进行原位杂交,分析Foxg1、Gbx2、Otx2、Fgf8和En1基因在Cep164基因缺失小鼠胚胎脑中的表达情况,确定Cep164基因对脑发育的影响。结果显示,与野生型相比,Cep164基因突变小鼠胚胎头部Gbx2、前中脑Otx2和中后脑的边界Fgf8表达无明显变化,但端脑泡Foxg1表达减少,前神经脊Fgf8表达减少,Otx2在晶状体板和嗅状体板表达减少,背侧中脑和后脑前En1表达减少。这说明Cep164基因是维持胚胎头部正常发育的重要基因,Cep164基因突变会导致前脑发育异常,主要影响嗅觉和视觉功能。
关键词:Cep164;基因缺失;小鼠胚胎;脑发育;原位杂交
中图分类号:S852.2 文献标识码:A
文章编号:0439-8114(2019)23-0139-03
DOI:10.14088/j.cnki.issn0439-8114.2019.23.033 开放科学(资源服务)标识码(OSID):
Effects of Cep164 gene deletion on embryonic mouse brain development
WANG Chun-hua
(School of Life Science and Technology,Mudajiang Normal University,Mudanjiang 157000,Heilongjiang,China)
Abstract: To reveal the function of Cep164 gene in mouse brain development, the expression level of Foxg1,Otx2,Gbx2,Fgf8 and En1 in embryonic brain with Cep164 gene deletion was detected by in situ hybridization experiment. The results showed that the expression of Gbx2 and Otx2 in the brain, and Fgf8 in the midbrain-hindbrain boundary, were no significant change in the Cep164 mutation embryonic mice, compared with wild type. But the expression of Foxg1 in telencephalic vesicles, the Fgf8 in anterior neural ridge, the Otx2 in eye and olfaction plate, and En1 in ventral midbrain and anterior hindbrain decreased. The all results reveal Cep164 is an important gene to maintain the normal brain development. Cep164 gene mutation could cause forebrain dysplasia, mainly affect olfaction and vision.
Key words: Cep164; gene deletion; embryonic mouse; brain development; in situ hybridization
Cep164是中心体激活元件Cep家族的一员,位于母中心粒基体末端,参与纤毛的形成过程[1]。研究人员通过siRNA技术沉默细胞中Cep164基因,证明当Cep164基因突变时会导致细胞中纤毛缺失[2]。Moumita等[3]在对纤毛相关肾消耗病患者的基因分析时发现,患者的Cep164基因发生纯合子点突变。通过斑马鱼试验发现,当Cep164基因突变时斑马鱼发生肾消耗病(Nephronophthisis-related ciliopathies,NPHP-RC),表型包括腎小管囊肿、脑积水和视网膜发育不良[3]。但关于哺乳动物的突变表型并未见相关报道。在前期试验中发现Cep164基因突变会导致胚胎发育异常,表型包括胚胎发育迟缓、心脏外膜增大扩张、血管发育不良、背侧神经管不对称发育、胚胎躯体异常弯曲、面部前脊神经管不闭合、头部神经管闭合不全、心脏反向扭转弯曲[4]。由于Cep164基因突变导致胚胎存活不到14.5 d,脑发育的表型并未观察到。本研究利用原位杂交技术对小鼠胚胎脑部发育中特异性基因En1、Otx2、Foxg1、Fgf8和Gbx2的表达与分布进行检测,一方面利用基因在胚胎脑发育过程中特定位置来确定Cep164基因对脑发育的影响,另一方面推测Cep164基因对脑发育的影响机制。
1 材料与方法
1.1 材料
Cep164基因敲除杂合子小鼠(Cep164+/-)馈赠于美国康奈尔大学。Cep164基因敲除杂合子雌鼠(Cep164+/-)与Cep164+/-雄鼠进行杂交,以合笼次日雌鼠见阴栓记为怀孕0.5 d(E0.5),在E10.5采用颈椎脱臼法处死怀孕母鼠,根据基因型检验获得E 9.5的Cep164-/-与野生型(WT)胎鼠。
1.2 探针准备
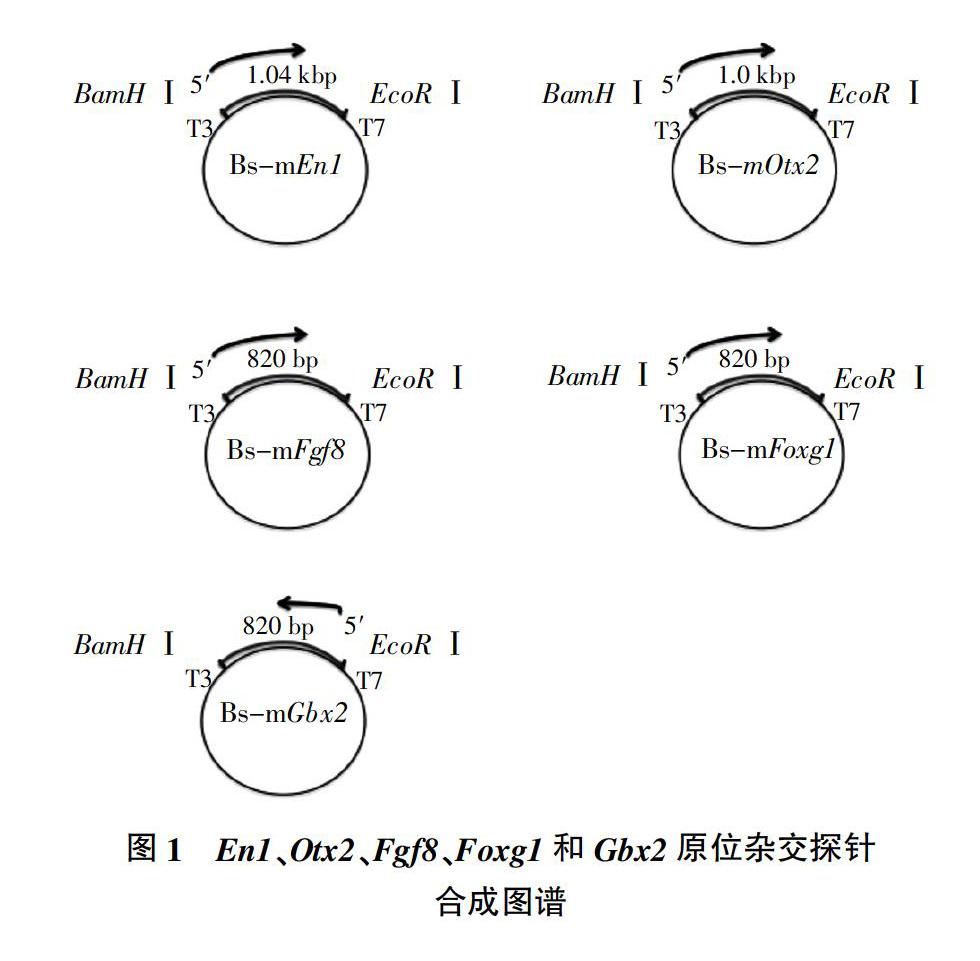
参照Graser等[1]的方法合成 En1、Otx2、Fgf8、Foxg1和Gbx2基因探针。采用5组引物(表1),选用巢式PCR分别扩增目标基因,纯化PCR产物经BamH Ⅰ和EcoR Ⅰ双酶切,连接PBS-SK质粒,转化大肠杆菌感受态细胞挑取单克隆,获得的重组质粒经测序确认序列无误及判断插入方向后,采用BamH Ⅰ将Bs-En1、Bs-Otx2、Bs-Fgf8和Bs-Foxg1重组质粒线性化,EcoR Ⅰ将Bs-Gbx2重组质粒线性化。如图1所示,以线性化片段为模版用T3或T7 RNA聚合酶体外转录合成带地高辛(DIG)标记的En1、Otx2、Fgf8、Foxg1和Gbx2基因探针。分别取1 μL反应物,用葡聚糖凝胶电泳进行探针检测。
1.3 原位杂交
将E10.5胚胎用4%多聚甲醛固定后在4 ℃条件下过夜,经甲醇梯度脱水,复水后,0.2%戊二醛再固定胚胎,参考Martinez-Barbera等[5]的方法,用En1、Otx2、Fgf8、Foxg1和Gbx2探针进行胚胎头部的原位杂交。
2 结果与分析
2.1 En1、Otx2、Foxg1、Fgf8和Gbx2原位杂交探针
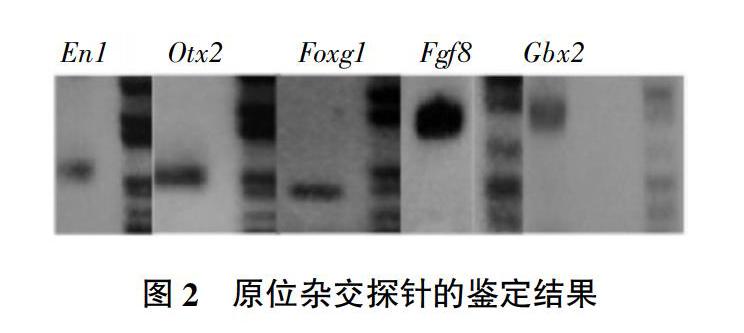
如图2所示,1 μL反应物的葡聚糖凝胶电泳结果显示,En1、Otx2、Foxg1、Fgf8和Gbx2的条带位置均与扩增目标条带位置相同,分别为1 040、1 000、820、820、820 bp,并且反应效率高,浓度足够作为探针与胚胎进行原位杂交。
2.2 Cep164-/-胚胎前脑发育异常
正常胚胎头部发育中,Foxg1表达在端脑泡(Telencephalic vesicle),如图3A所示。与其相比,图3a中,Cep164-/-小鼠胚胎头部Foxg1的表达范围减少,说明Cep164突变导致小鼠胚胎端脑泡体积减小。
正常胚胎头部发育中,Gbx2表达在中后脑边界区域(Mid-hind brian boundary,MHB),如图3B所示。与其相比,图3b中,Cep164-/-小鼠胚胎头部Gbx2的表达不变。
正常胚胎头部发育中,Otx2表达在晶状体板、嗅基板、前脑中脑区域,如图3C所示。与其相比,图3c中,Cep164-/-小鼠胚胎Otx2在晶状体板和嗅基板减少,在前脑中脑区域表达正常。
正常胚胎头部发育中,En1表达在背侧中脑和后脑前,如图3D所示。与其相比,图3d中,Cep164-/-小鼠胚胎背侧中脑和后脑前En1表达减少。
正常胚胎头部发育中,Fgf8表达在中后脑边界区域和前神经脊(Anterior neural ridge,ANR),如图3E和图3F所示。与图3E相比,图3e中,Cep164-/-小鼠胚胎头部中后脑边界区域表达范围无明显变化。与图3F相比,图3f中,Cep164-/-小鼠胚胎前神经脊表达范围明显减少。
3 讨论
在脊椎动物中,中脑和后脑边界提供了中脑和后脑喙部模式形成的基本组织信号。通过Otx2和Gbx2基因的表达可确定MHB最终位置,Otx2和Gbx2基因可激活MHB特异性基因Fgf8,Otx2或Gbx2基因的缺失不能抑制MHB的形成,但能改变MHB位置和Fgf8的表达区域,Otx2基因缺失可导致MHB向喙部移动,而Gbx2基因缺失导致MHB向尾部移动,当小鼠Otx2和Gbx2基因同时失活,Fgf8的表达完全正常,但表达区域变大,说明Fgf8的表达不依赖于这两个基因[5]。本研究发现,Cep164基因突变对胚胎头部发育中Gbx2表达在MHB和Otx2在前脑、中脑区域表达正常,Fgf8在头部中后脑边界区域表达范围也无明显变化,说明Cep164对MHB的形成位置和面积均无影响。
Oxt2在鼠胚胎发育过程中,除了在前脑、间脑的神经外胚层表达外,还在晶状体板表达[6],与Pax6.0共同调节控制视网膜上皮色素细胞的转分化及再生视网膜细胞的分化[7]。试验结果显示,Cep164基因突变导致晶状体板和嗅基板Otx2表达减少,说明Cep164基因的缺失影响了视觉和嗅觉正常功能的发育,Cep164基因突变可能会导致小鼠视觉和嗅觉功能障碍,这也是纤毛病变的表型之一。Fgf8是前脑发育过程中至关重要的信号分子[8,9],除了表达在中后脑边界区域,还在前神经脊表达,试验结果显示Cep164突变小鼠前神经脊Fgf8表达范围明显减少,Cep164基因突变小鼠的前脑小于野生型胚胎,这可能是Cep164基因突变导致小鼠胚胎前脑发育异常的原因之一。特异性表达在端脑泡的Foxg1基因对前脑正常发育也至关重要,Foxg1在Cep164基因突变小鼠胚胎中表达减少,端脑泡体积减小,此结果与Fgf8结论一致,进一步说明Cep164基因与胚胎前脑正常发育相关。
参考文献:
[1] GRASER S,STIERHOF Y D,LAVOIE S B,et al. Cep164, a novel centriole appendage protein required for primary cilium formation[J].J Cell Biol,2007,179(2):321-330.
[2] SCHMIDT K N,KUHNS S,NEUNER A,et al. Cep164 mediates vesicular docking to the mother centriole during early steps of ciliogenesis[J].J Cell Biol,2012,199(7):1083-1101.
[3] MOUMITA C,RANNAR A,AMIYA K,et al. Exome capture reveals ZNF423 and CEP164 mutations,linking renal ciliopathies to DNA and damage response signaling[J].Cell,2012,150(3):533-548.
[4] 王春花.Cep164基因在小鼠胚胎生長发育过程中的功能及作用[D].哈尔滨:东北农业大学,2014.
[5] MARTINEZ-BARBERA J P,SIGNORE M,BOYL P P,et al. Regionalisation of anterior neuroectoderm and its competence in responding to forebrain and midbrain inducing activities depend on mutual antagonism between OTX2 and GBX2[J].Development,2001,128(23):4789-4800.
[6] CVEKL A,PIATIGORSKY J. Lens development and crystalline gene expression:Many roles for Pax6[J].BioEssays,1996,18(8):621-630.
[7] AVDONIN P P,MARKITANTOVA Y V,ZINOVIEVA R D,et al. Expression of regulatory genes Pax6,Otx2,Six3,and FGF2 during newt retina regeneration[J].Biology bulletin,2008,35(4):355-361.
[8] MEYERS E N,LEWANDOSKI M,MARTIN G R. An Fgf8 mutant allelic series generated by Cre-and Flp-mediated recombination[J].Nat Genet,1998,18(2):136-141.
[9] SHIMAMURA K,RUBENSTEIN J L. Inductive interactions direct early regionalization of the mouse forebrain[J].Development,1997, 124:2709-2718.
