烟草NtCIPK家族基因表达与其含钾量之间的关系分析
2019-01-09杨尚谕李立芹任学良鲁黎明
杨尚谕,卓 维,陈 倩,蒋 垚,童 铸,李立芹,任学良,鲁黎明
(1.四川农业大学 农学院,四川 成都 611130;2.贵州省烟草科学研究院,贵州 贵阳 550003)
钾在植物中含量很高,是植物必需的大量元素之一,在植物的光合作用、气孔运动、蛋白合成和氧化代谢等生命活动中起重要作用[1-3]。缺钾时,会导致植物生长受限和代谢紊乱,严重缺钾时,植物甚至无法正常生长[4]。
研究表明,不同植物的钾含量不同。胡笃敬等[5]研究了30种植物的含钾量,包括水生植物、陆生野生植物、绿肥植物、早稻和晚稻等,结果发现,不同植物间的钾含量差别很大。此外,同一植物的不同品种钾含量也不同,如大豆[6]、燕麦[7]、小麦[8]、香蕉[9]、甘薯[10]、甘蔗[11]、玉米[12]、西瓜[13]、油菜[14]等。烟草品种间的烟叶含钾量,也存在明显差异[15-17]。杨欢等[18]研究了93份烟草种质资源材料的烟叶钾含量,并根据钾含量的高低,将这些种质材料分为高钾基因型、普通型与低钾型。
在分子层面,植物对外界钾的吸收,主要通过高亲和与低亲和2类吸收机制进行[3]。其中,低亲和机制主要是由位于细胞质膜上的钾离子通道承担,是植物吸收钾离子的重要途径之一[19-20]。例如,拟南芥钾离子通道的AKT1,是拟南芥根从外界获取K+的主要组分之一[21]。CIPK是一类植物特有的丝氨酸/苏氨酸蛋白激酶[22],CBL-CIPK信号系统广泛参与了植物钾吸收调节过程[23]。研究表明,在AtCBL1/9的介导下,AtCIPK23通过磷酸化AKT1,激活该离子通道,从而增强了拟南芥在低K+条件下对K+的吸收[24]。在水稻中,OsCBL1-OsCIPK23同样通过调控OsAKT1,参与了水稻的K+吸收[25]。在胡杨中,PeCBL1-PeCIPK23通过磷酸化钾离子通道PeKC1、PeKC2,从而实现其对低钾胁迫的响应[26]。还有研究表明,AtCIPK23在烟草中的过表达导致了转基因烟草低K+耐受性的增加[27]。此外,甘蔗CIPK23[28]、水稻CIPK10[22]、林烟草CIPK3[29]、普通烟草CIPK2等[30]均可能参与了植物对K+的吸收调节过程。
烟草是我国重要的经济作物,烟叶含钾量高低,是衡量烟叶品质的重要指标之一[31]。然而,目前对于烟草钾吸收的分子机制研究尚不够深入,尤其是NtCIPK基因参与钾吸收调控的研究还较少,其调控机制尚不清楚。因此,本研究对3个烤烟品种进行了不同K+浓度的处理,研究了其干质量、钾含量及NtCIPK家族11个基因的表达模式,并分析了这3个品种钾含量和其NtCIPK基因表达之间的关系,旨在为烟草钾吸收调控的分子机制研究提供一定的参考。
1 材料和方法
1.1 材料及试剂
参试烤烟品种为K326、贵烟5号、云87。
试验所需的TRIzol试剂、cDNA合成试剂盒、SYBR Green Master mix等试剂购自宝生物工程(大连)有限公司。
1.2 试验方法
1.2.1 不同钾浓度培养溶液的配制 不同钾浓度的培养液,在霍格兰溶液配方基础上进行适当调整配制而成(表1)。用氯化钾调整培养溶液的钾浓度分别为6.0,1.0,0.2 mmol/L,其他不变。
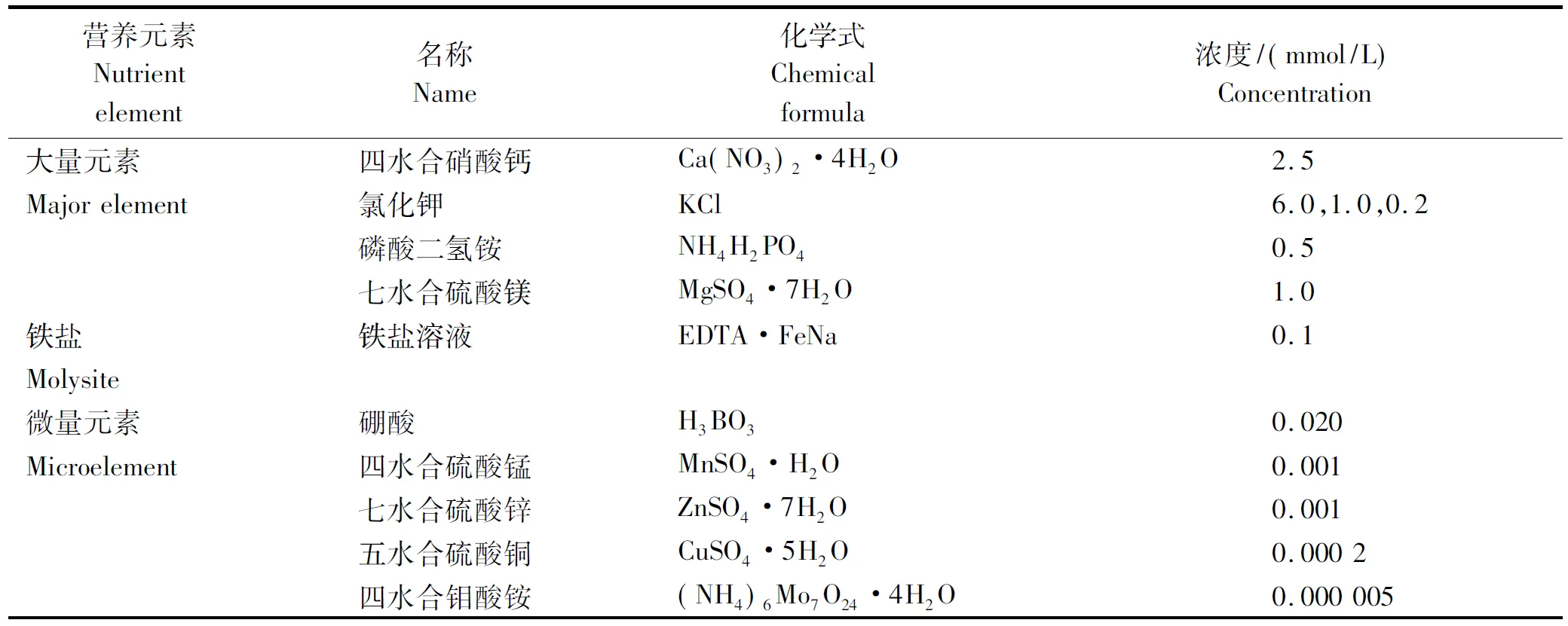
1.2.2 烟草幼苗的培养与处理 挑选大小一致且饱满的烟草种子,表面消毒后,分别播种在白色泡沫漂浮育苗盘(长65.5 cm、宽33.5 cm、高4.5 cm、162穴)中,然后,将育苗盘放置在育苗池中进行育苗。育苗池中加注K+浓度为6 mmol/L的霍格兰溶液。60 d后,每品种挑选长势一致的幼苗各90株,分为3组,分别放入K+浓度为6.0,1.0,0.2 mmol/L的霍格兰溶液中,进行处理。每个处理包括30株烟苗,10株烟苗为一个重复,共重复3次。
处理48 h后,对每个浓度每个品种的烟苗进行随机取样。取样时,取每个重复烟苗叶片混合样,在液氮中速冻后,置于-80 ℃冰箱中保存,用于RNA的提取。
处理28 d后,将参试的全部烟株取出,并将根部冲洗干净,用吸水纸吸干水分后,在烘箱中105 ℃杀青30 min,80 ℃下烘干至恒质量待用。
1.2.3 烟株干质量及钾含量的测定 用电子天平称取烟株的干质量。植株钾含量的测定采用火焰光度计法[32]进行。
1.2.4 NtCIPK基因的表达模式分析 RNA的提取:采用TRIzol法进行,cDNA的合成参照试剂盒说明,具体参照卓维等[30]的方法进行。

引物设计:根据四川农业大学农学院的分子生物技术验室前期所克隆到的11个NtCIPK基因的cDNA序列,采用Primer Premier 5.0设计qPCR引物(表2)。
qPCR扩增:以反转录后的cDNA为模板,以烟草18S rRNA为内参进行qPCR扩增。qRT-PCR扩增体系为:2×Prime STAR Max 5.0 μL,上游引物 0.5 μL,下游引物 0.5 μL,cDNA 1 μL,ddH2O 3 μL。qRT-PCR扩增程序为:50 ℃反应2 min,95 ℃预变性3 min;然后,95 ℃变性10 s,65 ℃延伸45 s,进行39个循环。基因相对表达水平用2-ΔΔCt方法进行计算。
2 结果与分析
2.1 各处理烟草植株生物量比较
在不同K+浓度处理下,各品种生物量的测定结果如图1所示。随着培养液K+浓度的降低,各品种的植株干质量均呈下降趋势。在1.0,0.2 mmol/L K+浓度处理下,云87与贵烟5号的干质量差异不显著,但在0.2 mmol/L K+浓度处理下云87与贵烟5号的干质量均显著高于K326;在6.0 mmol/L K+浓度处理下,贵烟5号的植株干质量显著低于K326和云87,后二者之间的差异则不显著。烟草植株生物量的测定结果表明,外界供K+量的高低,显著影响到烟草植株的生长与干物质的积累。

图中每个浓度不同品种的不同小写字母表示在P<0.05水平上差异显著。图2-3同。Different lowercase letters in each concentration of the figure indicate significant difference at P<0.05.The same as Fig.2-3.
2.2 不同处理烟草植株钾积累量比较
图2为不同K+浓度处理下,各处理烟草植株钾积累量(株/g)测定结果。随培养液中K+浓度的降低,各参试材料植株的钾积累量都呈下降趋势。在6.0,1.0 mmol/L K+浓度处理下,云87的钾积累量显著高于K326及贵烟5号;而在0.2 mmol/L K+浓度处理下,云87与贵烟5号和K326的钾积累量差异显著。钾积累量的测定结果表明,不同烟草品种在不同的外界K+浓度环境中,其植株的K+积累特性明显不同。
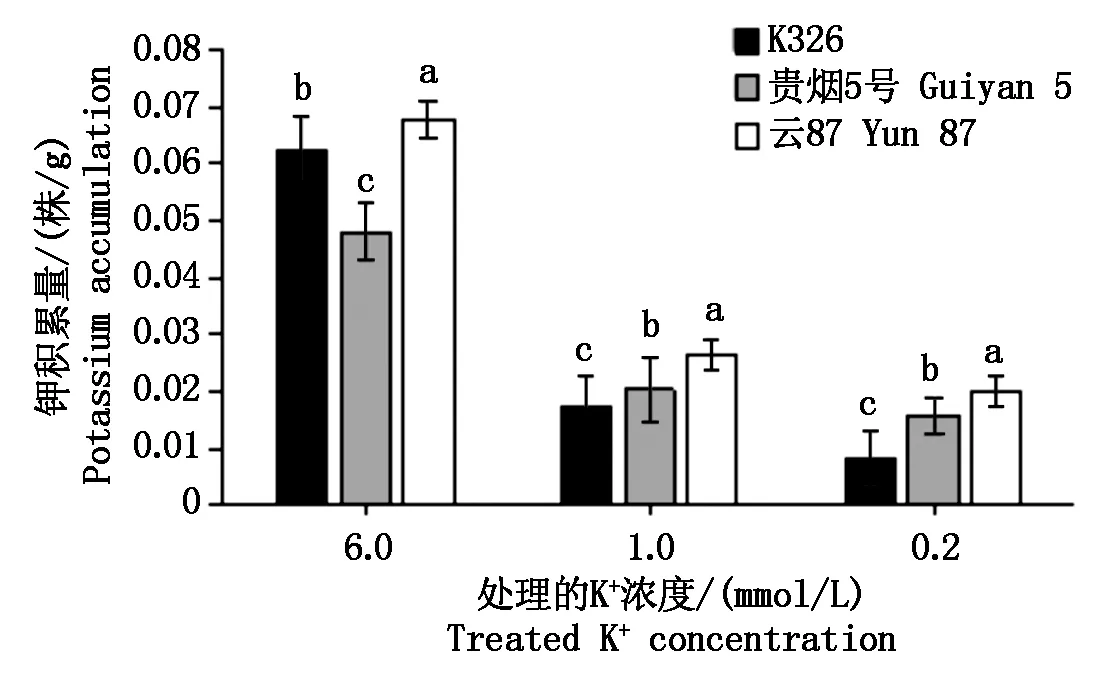
图2 各品种烤烟不同K+浓度处理的K+积累量Fig.2 K+ accumulation of different K+ concentrations of flue-cured tobacco of different varieties
2.3 不同供钾浓度下NtCIPK基因的表达模式分析
对在不同K+浓度处理下,11个NtCIPK基因在各烟草品种中的表达进行了测定,结果如图3所示。
从图3可以看出,NtCIPK基因的表达均随着外界钾浓度的变化而改变。在低钾条件下(0.2 mmol/L),其表达量基本是上调的,其中,云87的11个NtCIPK基因表达量均高于对照K326,且存在显著性差异,显示这些基因的表达受低钾胁迫的诱导。但NtCIPK9和NtCIPK24分别在K326及贵烟5号中的表达,随着外界钾浓度的降低,却呈下调趋势,即随着K+浓度的降低而下调,都分别显著低于云87的表达量。在所检测的11个NtCIPK基因中,NtCIPK2、NtCIPK5、NtCIPK6、NtCIPK9、NtCIPK12、NtCIPK14、NtCIPK23和NtCIPK24随着K+浓度的降低,表达量变化较大。可以推测,这8个NtCIPK基因可能以某种直接或间接的方式参与了烟草钾吸收的调控。
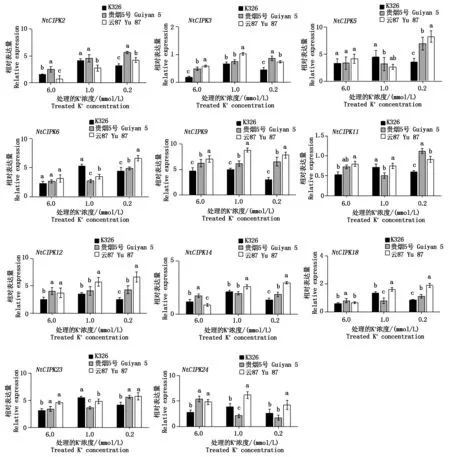
图3 不同烤烟品种在不同K+浓度处理后的NtCIPK基因的相对表达量Fig.3 Relative expression of NtCIPK gene after treatment with different K+ concentrations in different flue-cured tobacco varieties
2.4 烟草钾含量与其CIPK基因表达量之间的关系分析
2.4.1 逐步回归分析 将各处理烟草植株的K+积累量(Y)与11个NtCIPK基因的表达量进行逐步回归分析,得出了3个烟草品种在3个钾浓度处理下的逐步回归方程(表3)。
对K326来说,NtCIPK12(X7)、NtCIPK18(X9)和NtCIPK3(X2)的表达,分别在6.0,1.0,0.2 mmol/L条件下,对其K+积累量影响显著。贵烟5号的情况有所不同,其K+积累量在6.0,1.0 mmol/L条件下与NtCIPK5(X3)的表达密切相关;而在0.2 mmol/L条件下,则受NtCIPK3(X2)表达的显著影响。NtCIPK23(X10)的表达,在外界高K+浓度(6.0 mmol/L)下,与云87植株的K+积累量相关性显著;而在中(1.0 mmol/L)、低钾浓度(0.2 mmol/L)下,云87植株的K+积累量则与NtCIPK9(X5)的表达相关性显著。
2.4.2 通径分析 在回归分析的基础上,对NtCIPK3(X2)、NtCIPK5(X3)、NtCIPK9(X5)、NtCIPK12(X7)、NtCIPK18(X9)及NtCIPK23(X10)做了进一步的通径分析(表4)。
从逐步回归方程和通径分析结果来看,在外界0.2 mmol/L K+浓度下,NtCIPK3(X2)的表达与K326和贵烟5号的植株钾积累量分别呈显著正相关和显著负相关。同时,NtCIPK5(X3)在6.0 mmol/L浓度下的表达,与贵烟5号的植株钾积累量呈显著正相关。此外,在6.0 mmol/L K+浓度处理下,云87的植株钾积累量与NtCIPK23(X10)的表达显著负相关。
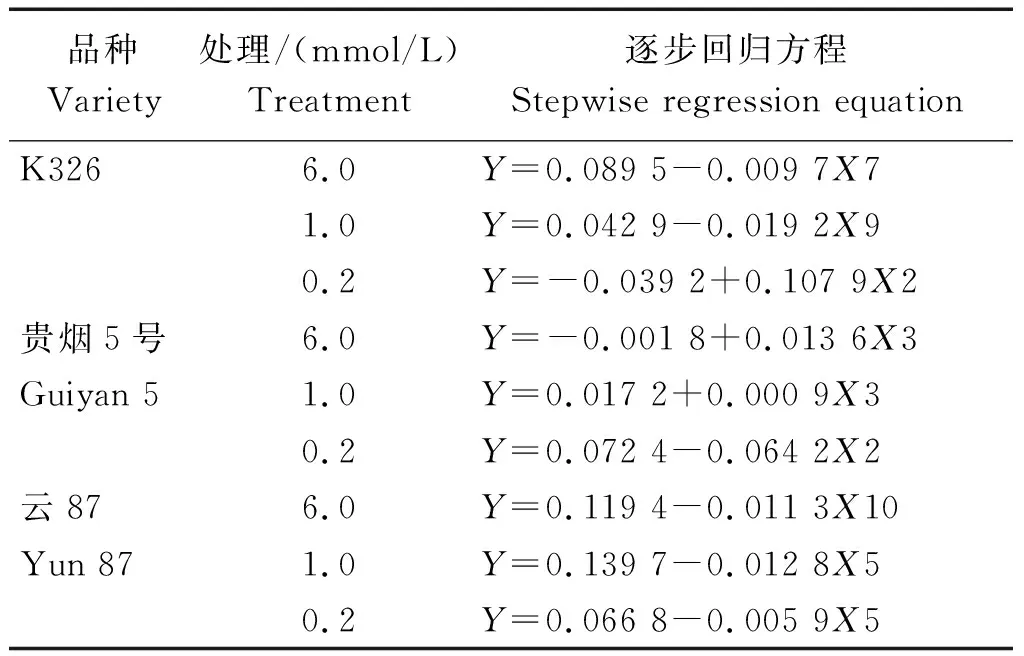
表3 各烤烟品种的K+含量和NtCIPK基因表达模式的逐步回归方程Tab.3 Stepwise regression equations of K+ content and NtCIPK gene expression pattern of various flue-cured tobacco varieties
注: Y.对应各浓度烤烟品种的K+含量;X1、X2、X3、X4、X5、X6、X7、X8、X9、X10、X11分别代表NtCIPK2、NtCIPK3、NtCIPK5、NtCIPK6、NtCIPK9、NtCIPK11、NtCIPK12、NtCIPK14、NtCIPK18、NtCIPK23、NtCIPK24基因的相对表达量。表4同。
Note: Y. The K+content of the corresponding flue-cured tobacco varieties;X1, X2, X3, X4, X5, X6, X7, X8, X9, X10, and X11 represent relative expression levels ofNtCIPK2,NtCIPK3,NtCIPK5,NtCIPK6,NtCIPK9,NtCIPK11,NtCIPK12,NtCIPK14,NtCIPK18,NtCIPK23, andNtCIPK24 genes, respectively. The same as Tab.4.
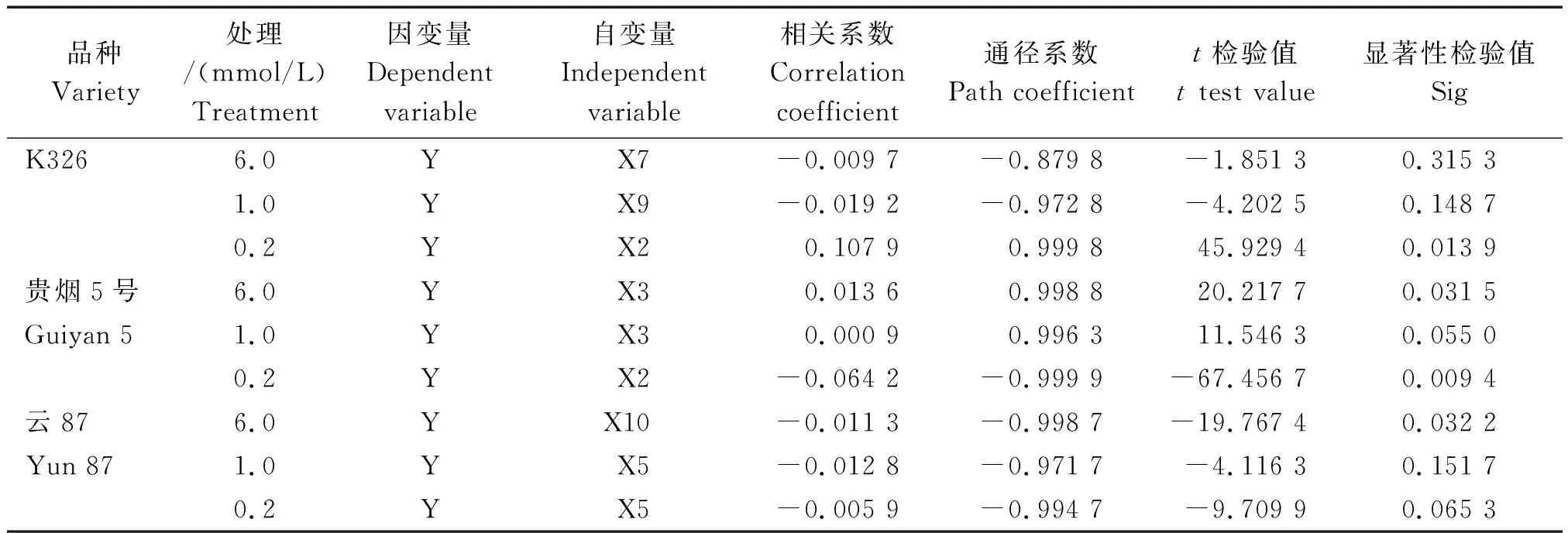
表4 各烤烟品种的K+含量和NtCIPK基因表达模式的通径分析Tab.4 Path analysis of K+ content and NtCIPK gene expression patterns of flue-cured tobacco varieties
3 讨论
本研究通过对3个烟草品种进行不同K+浓度处理,测定了其干质量、钾积累量,并分析了11个NtCIPK基因在各个处理下的表达量,结果表明,NtCIPK基因的表达量与烟草的钾积累量密切相关。
前人研究结果表明,植物的CIPK基因在植物对K+的吸收调控过程中起着重要的作用[23-26]。在本研究中,烟草11个NtCIPK基因在不同品种中的表达量并不相同,并且均能够响应外界钾浓度的变化,这与前人的研究结果相一致[25,28-30]。此结果暗示,烟草的NtCIPK基因广泛参与了烟草对钾的吸收调控过程。
植物的CIPK基因参与钾吸收调控的机制较为复杂。一般情况下,其过表达能够增强植物对钾的吸收[24-27,33 ]。然而,拟南芥AtCIPK9[34]的过表达则导致对低K+敏感;玉米ZmCIPK9[33]的过量表达却提高了钾的吸收能力。由此说明,不同的NtCIPK基因在植物钾吸收过程中的调控机制不尽相同。本研究也发现,从NtCIPK基因的表达模式来看,随着外界钾浓度的降低,其表达可以分为上调及下调2种情况。有9个NtCIPK基因是上调表达,只有K326的NtCIPK9与贵烟5号的NtCIPK24的表达为下调。其中,上调表达的基因,就包括了与前人的研究结果相一致[29-30, 35]的CIPK2、CIPK3、CIPK23基因。烟草NtCIPK基因的这种不同表达模式,在一定程度上说明了其参与烟草钾吸收调控时,存在正调控、负调控或者其他的复杂方式。
逐步回归和通径分析的结果同样说明,在11个NtCIPK基因中,其表达量与烟草钾积累量的相关性存在显著与不显著之别;即使对同一个基因来说,其在不同品种中的作用也不尽相同。例如,同样在0.2 mmol/L K+浓度处理下,NtCIPK3(X2)与K326的K+含量呈显著正相关;然而,与贵烟5号的K+含量则呈显著负相关。该结果同样说明了烟草NtCIPK基因行使钾吸收调控功能的复杂性。
虽然烟草NtCIPK基因广泛参与了烟草对钾的吸收与调控,然而,其参与的方式、调控的模式与功能行使的途径却十分复杂。而每一个NtCIPK基因在其中发挥作用的详细机制,则需更进一步地深入研究。
