獭兔CREB1基因的编码区克隆、生物信息学及表达规律分析
2019-01-09秦立志赵博昊胡帅帅翁巧琴鲍国连吴信生
陈 阳, 秦立志, 赵博昊, 胡帅帅, 穆 琳, 翁巧琴, 鲍国连, 吴信生
(1.扬州大学动物科学与技术学院,江苏扬州 225009; 2.浙江省余姚市欣农兔业有限公司,浙江余姚 315400;3.浙江省农业科学院,浙江杭州 310021)
獭兔,即力克斯兔(Rex rabbit),是皮用型兔的代表之一,其被毛颜色丰富多样,且纯天然的被毛不掉色或褪色,深受消费者青睐。目前,美国认可的獭兔毛色有14种,欧洲认可的有30种,我国以青紫蓝色、黑色、海狸色、白色为主。一般而言,动物的毛色与皮肤中黑色素的数量和分布密切相关,同时许多内在因子也影响着毛色性状的表现[1]。哺乳动物毛色主要取决于黑色素沉积,黑色素细胞产生真黑素和棕黑素,二者分布的不同致使哺乳动物形成多种毛色类型[2-3]。笔者所在课题组前期利用表达谱芯片筛选与獭兔毛色形成相关的基因,发现CREB1基因在不同毛色的獭兔皮肤中表达差异显著,表明该基因可能参与到獭兔的毛色形成过程中[4]。
环腺苷酸反应元件结合蛋白(cAMP responsive element binding protein,CREB)属于DNA蛋白结合类亮氨酸拉链结构转录因子超家族,主要与cAMP等信号发生应答反应[5]。由于mRNA具有稳定性调节和翻译后修饰的调控作用,机体细胞内存在多种CREB基因的剪接方式,可以产生不同的剪接体,CREB1即为该家族中常见成员之一[6-7]。已有文献报道,磷酸化CREB可以激活下游转录因子MITF,已知MITF是主控调节黑素细胞的发展、功能和生存的基因,可激活酪氨酸酶基因的转录[8-10]。基于此,本研究通过分子克隆等技术手段,获得了獭兔CREB1基因的编码区序列(coding sequence,CDS),并利用荧光定量手段检测其在不同獭兔皮肤中的表达规律,为进一步探究CREB1基因的功能及毛色形成机制提供基础。
1 材料与方法
1.1 材料
1.1.1 试验动物 选取成年青紫蓝色獭兔、白色獭兔和黑色獭兔各3只,耳缘静脉注射空气处死,取约1 cm2相同位置的背部皮肤组织,-70 ℃保存。
1.1.2 试验试剂 Gel Extraction Kit、DNA marker、dNTPs、TaqDNA聚合酶、反转录试剂盒、限制性内切酶Hind Ⅲ、BamHⅠ、大肠杆菌感受态细胞DH5α、pMD19-T Simple Vector购于宝生物工程(大连)有限公司;Trizol试剂、质粒提取试剂盒、SYBR®Green Master Mix试剂盒购于天根生化科技(北京)有限公司。
1.2 方法
1.2.1 总RNA的提取与检测 采用Trizol法提取总RNA,通过1%变性琼脂糖凝胶和NanoDrop 2000核酸蛋白测定仪,检测总RNA的完整性、浓度和D260 nm/D280 nm。
1.2.2 引物设计 根据GenBank数据库中提供的CREB1基因CDS序列(XM_002712603)设计特异性克隆引物,并分别在上下游引物中引入Hind Ⅲ和BamHⅠ酶切位点,同时设计兔CREB1、GAPDH基因实时荧光定量PCR引物,由生工生物工程(上海)股份有限公司合成(表1)。
1.2.3CREB1基因的克隆 将总RNA进行反转录反应,利用CREB1基因特异性引物扩增编码区序列。50 μL PCR反应体系:ddH2O 33.5 μL,10×buffer 5 μL,dNTP 5 μL,正向引物2 μL,反向引物2 μL,模版2 μL,TaqDNA聚合酶0.5 μL。PCR反应条件:95 ℃预变性5 min;95 ℃变性35 s,60 ℃退火35 s,72 ℃延伸1 min,共35个循环;72 ℃延伸 10 min,4 ℃保存。构建pMD19-T-CREB1克隆质粒,用Hind Ⅲ和BamHⅠ进行双酶切鉴定,选取阳性质粒送生工生物工程(上海)股份有限公司测序。

表1 CREB1基因克隆引物序列
注:限制性内切酶位点即序列中下划线的部分。
1.2.4 生物信息学分析 进行氨基酸序列同源性比对,利用软件MEGA 5.1构建系统进化树;蛋白质组成及疏水性分析:http://cn.expasy.org/tools/protparam.html/;蛋白质信号肽预测:http://www.cbs.dtu.dk/services/SignalP/;蛋白质跨膜区预测:http://www.cbs.dtu.dk/services/TMHMM-2.0/;蛋白质亚细胞定位分析:http://psort.hgc.jp/form2.html/;用Hopfield神经网络预测CREB1蛋白的二级结构(http://npsa-pbil.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_gor4.html);用swiss-model(http://swissmodel.expasy.org/workspace/index.php)预测CREB1蛋白的三级结构。
1.2.5 实时荧光定量PCR 根据荧光定量试剂盒说明书,分别对青紫蓝色、白色和黑色獭兔总RNA通过反转录反应,进行qRT-PCR试验。20 μL反应体系:AceQTMqPCR SYBR®Green Master Mix 10 μL,正向引物0.4 μL,反向引物0.4 μL,ROX Reference Dye 2 0.4 μL,DNA模板2.0 μL,灭菌蒸馏水6.8 μL。反应程序:95 ℃预变性5 min;95 ℃变性 10 s,60 ℃退火30 s,40个循环。融解曲线:95 ℃ 15 s,60 ℃ 60 s,95 ℃ 15 s。以白色獭兔为对照组,以GAPDH基因为内参基因,采用2-ΔΔCT法计算CREB1基因的相对表达量,利用SPSS 22.0统计软件进行差异显著性分析。
2 结果与分析
2.1 獭兔CREB1基因编码区的克隆
利用1%琼脂糖凝胶电泳检测獭兔背部皮肤总RNA,显示28S、18S和5S 3条带,说明RNA完整性较好。用NanoDrop 2000检测发现,总RNA无DNA和蛋白质污染,符合RT-PCR要求。利用CREB1基因特异性引物进行扩增,经琼脂糖凝胶电泳检测,其产物大小与预期(984 bp)相符,且条带清晰、特异性好(图1-A)。
利用TA克隆原理,将CREB1基因连接至pMD19-T Simple载体上,获得克隆质粒pMD19-T-CREB1,并采用限制性内切酶Hind Ⅲ和BamHⅠ进行双酶切鉴定,结果显示大小约2 600、1 000 bp的2条带(图1-B),与预期结果(2 692、984 bp)相符,说明CREB1基因克隆成功。
2.2 獭兔CREB1基因的序列分析
经比对,克隆序列与NCBI(美国国立生物技术信息中心)已知序列相似度为99.6%,将克隆序列提交至GenBank(登录号为KJ139984)。CREB1基因CDS含984个核苷酸(51.63% A+T,48.37% C+G),共编码327个氨基酸,相对分子量约为35.05 ku。利用ExPASy中的工具ProtParam分析得出,CREB1蛋白等电点为5.30,不稳定性系数为61.13,为不稳定性蛋白;CREB1蛋白预测GRAVY值为-0.443,为亲水蛋白。通过SignalP 4.0和TMHMM预测发现,该蛋白无信号肽、无跨膜区,亚细胞预测显示该蛋白主要定位于细胞核(表2),推测该蛋白为核内可溶性蛋白。
用Hopfield神经网络和Swiss-model分别对CREB1蛋白的二级结构和三级结构进行预测。结果如图2所示,CREB1蛋白二级结构中含有33.33%α螺旋(h),44.95%无规卷曲(c),21.71%延伸链(e);三级结构为1条螺旋链。
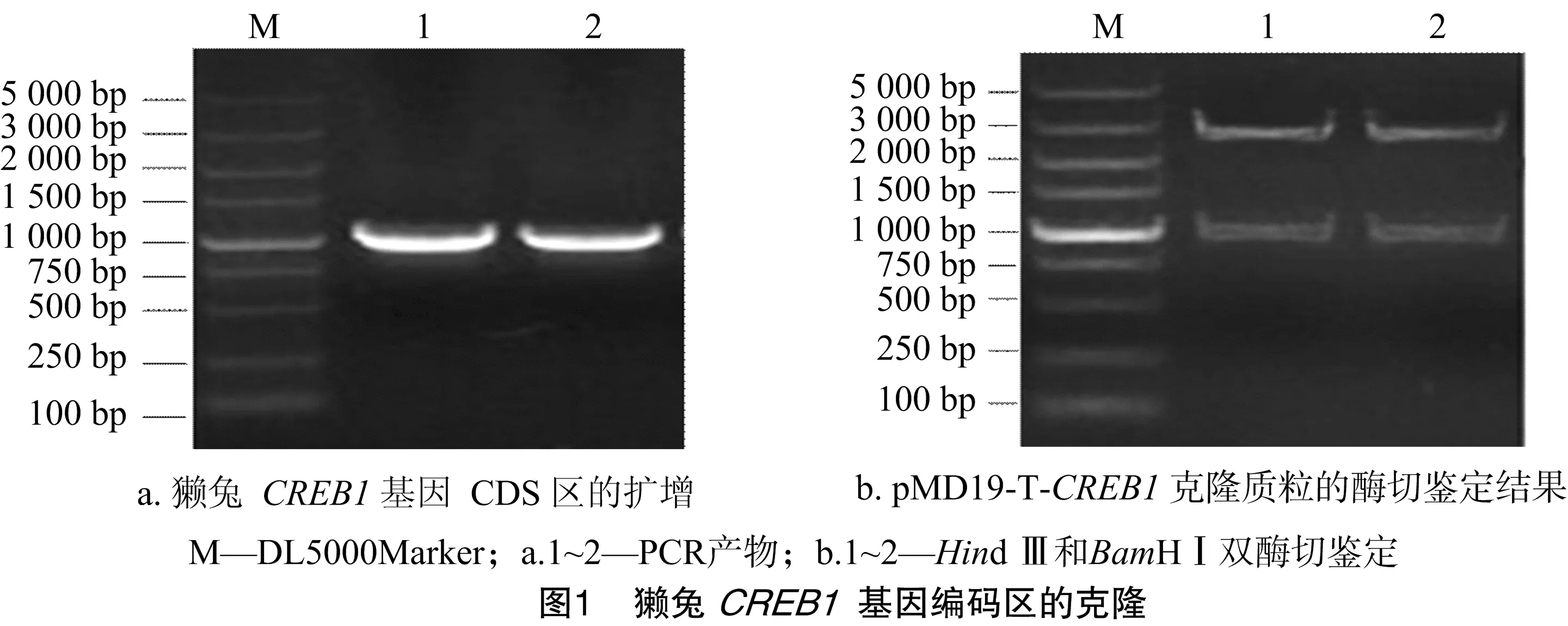

表2 CREB1蛋白质的亚细胞定位分析
2.3 獭兔CREB1蛋白与其他物种的同源性比较及系统进化分析
将克隆测序得到的獭兔CREB1基因CDS与人(Homosapiens)、牛(BosTaurus)、猪(Susscrofa)、鸡(Gallusgallus)、小鼠(Musmusculus)、猕猴(Macacamulatta)、猩猩(Pongoabelii)、大鼠(Rattusnorvegicus)、斑胸草雀(Taeniopygiaguttata)、非洲爪蟾(Xenopuslaevis)进行同源性比较发现,獭兔CREB1基因氨基酸序列与人、猪等哺乳动物的同源性较高,为92.9%~95.4%,而与其他非哺乳动物(鸡、斑胸草雀等)的同源性较低。利用MEGA 5.1软件构建邻接法(NJ)系统进化树,发现兔与人、猩猩、猕猴聚为1支,系统进化情况基本上与其亲缘关系远近一致,结果如图3所示。


2.4 CREB1基因在不同毛色獭兔皮肤中的表达分析
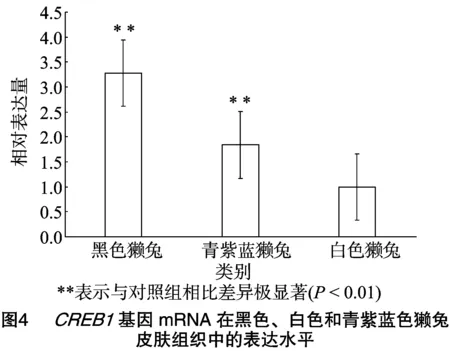
通过qRT-PCR检测黑色、白色和青紫蓝色獭兔皮肤组织中CREB1基因mRNA的表达水平,结果发现,CREB1基因在黑色獭兔和青紫蓝色獭兔中的表达量均极显著高于白色獭兔(P<0.01),且在黑色獭兔中的表达量最高(图4)。
3 讨论
cAMP反应元件(cAMP response element,CRE)是指基因启动子中的一段特定的DNA序列5′-TGACGTCA-3′[11]。CREB(环磷腺苷效应元件结合蛋白)是PKA(蛋白激酶A)、MAPK(丝裂原激活的蛋白激酶)、PKC(蛋白激酶C)、CaMKs(钙调蛋白激酶)等多种蛋白激酶的磷酸化底物,这些蛋白激酶可将CREB蛋白KID(激酶诱导区)的Ser(丝氨酸)-133位点磷酸化,从而影响CREB的活性[12-15]。已知,cAMP-PKA信号通路介导的磷酸化反应在黑色素生成过程中发挥了重要作用,CREB则是活化的PKA的主要靶标蛋白,磷酸化后CREB激活MITF的转录,促使MITF与TYR基因启动子Mbox结合,从而激活TYR的表达,促进黑色素生成。反之,降低CREB的磷酸化水平,MITF和酪氨酸酶也会受到一定的抑制[16]。在笔者所在课题组前期研究中,通过高通量技术手段发现CREB1基因与獭兔的毛色形成有关,与已有报道相符。
在本研究中,笔者发现獭兔CREB1基因的氨基酸序列与NCBI中其他哺乳动物具有很高的同源性,即CREB1基因在哺乳动物中非常保守,那么CREB1基因在獭兔毛色形成中是否具有同样的功能呢?已有报道指出,CREB1基因可调控MC1R基因与MITF基因之间色素沉着信号的传递,进而影响黑色素的沉积[17]。通过对黑色素生成信号通路的进一步研究发现,MC1R对CREB1有激活作用,增强MITF基因的表达,活化了与黑色素形成密切相关的基因,如TYR、TYRP1和KIT,所以CREB1在黑色皮肤中的表达量较高[18]。本研究中獭兔CREB1基因在黑色獭兔和青紫蓝色獭兔皮肤中表达水平高于白色獭兔,也印证了其他学者的观点,CREB1基因对黑色素的形成有明显的促进作用。
本研究利用分子克隆手段获得獭兔CREB1基因的编码区序列,通过生物信息学预测发现,该蛋白为无信号肽、无跨膜区的核内可溶性蛋白,且在哺乳动物中非常保守。同时发现,CREB1基因的表达量与毛色的黑色素沉积存在一定的关联性,本研究为探索獭兔毛色形成机制提供了一定的基础资料。
