ACTN3基因R577X多态性与速度及力量外其他运动表型及 机体衰老的关联研究述评
2019-01-06周文婷
周文婷
摘 要:作为运动能力遗传学研究的热点目标基因,ACTN3基因R577X多态性与优秀速度及力量素质间的关联性已被广泛证实。但除此以外,该多态位点的其他生理学效应研究多年来在我国尚未广泛开展。对国际上迄今为止该多态位点与其他运动相关表型(运动适应性、运动恢复及运动损伤风险)及机体衰老间相关性研究的主要成果进行了综述,旨在为我国开展相关研究提供参考,并为未来运动训练方案的个性化实施和老年病的个性化防治提供新的视角。
关 键 词:ACTN3基因;基因多态性;运动适应性;运动恢复;运动损伤风险;机体衰老;综述
中图分类号:G804.2 文献标志码:A 文章编号:1006-7116(2019)06-0139-06
Abstract: As the hot topic target gene of research on sports ability genetics, ACTN3 R577X polymorphisms correlation with excellent speed and strength qualities has been widely confirmed. But besides that, research on the other physiological effects of this polymorphism has not been widely developed in China for many years. The author gave an overview of the main results of international research on the correlation between R577X polymorphism and other sports phenotypes (sports adaptation, sports recovery and sports injury risk) as well as body aging so far, so as to provide reference for developing related research in China, and to provide a new perspective for the individualized implementation of sports training programs and the individualized prevention and treatment of aged diseases in the future.
Key words: ACTN3;polymorphism;sports adaptation;sports recovery;sports injury risk;body aging;overview
体质与运动能力是受多因素影响的复杂表型,多种微效基因及环境因素对其发挥重要作用,并存在基因-基因及基因-环境间的相互效应[1]。自1997年起体质与运动能力的遗传学研究在全球范围内广泛开展,但迄今发现的200多个与运动能力或训练效果相关的多态位点中,大约仅20个与优秀运动能力确切关联[1],而其中,ACTN3基因R577X多态位点被国内外大量研究证实是优秀速度及力量素质的分子标记,更被冠以“冠军基因”用于优秀运动员的基因选材[2]。虽然目前的R577X多态研究大多围绕其与优秀运动能力之间的关联展开,但各种文献表明,该位点也可能对其他的一系列生理表型发挥作用,如运动适应性、运动恢复和运动损伤风险,以及机体衰老过程中的肌肉功能、骨密度和能量代谢[3-8]。为此,本文综述了迄今为止上述研究领域中的主要结果,旨在为我国研究者开展相关研究提供参考,为运动员运动训练方案的个性化实施和老年病的个性化防治提供参考。
1 ACTN3基因R577X多态位点概述
α-辅肌动蛋白(Alpha-actinin,ACTN) 是肌动蛋白的结合蛋白,分布于骨骼肌Z线,可与细肌丝相结合来维持肌纤维的有序排列和收缩功能[9]。ACTN有4种形式,其中ACTN2和ACTN3表达于人类骨骼肌,但后者仅特异性表达于快肌纤维,并对肌动蛋白的锚定和肌纤维收缩的调节起重要作用[2]。研究发现,R577X多态位点定位于ACTN3基因的第16外显子,由于发生了C→T多态,导致其编码的577位精氨酸变为不编码蛋白的X,从而使携带纯合型X等位基因者ACTN3缺失,表现为较低的快肌纤维比率[10]。
研究发现,XX基因型的分布频率具有显著的种族差异,在亚洲人中约25%、欧洲白人中约18%、埃塞俄比亚人中约11%、牙买加人和非洲裔美国人中约3%、肯尼亚和尼日利亚人中则仅有1%表达该基因型[3]。目前已知,全世界ACTN3表达缺失的人口大约为15亿,但由于其基本功能可由 ACTN2补偿,因此其表达缺失并不致引起肌肉疾病[9]。此外,研究还发现,人类的ACTN2和ACTN3在氨基酸水平上存在80%以上的相似性,故二者间的这种强序列保守性及ACTN3的高度组织特异性表达均表明,ACTN3蛋白在快肌纤维中可发挥特殊作用[2]。
2 ACTN3基因R577X多态性与多种运动相关表型间的关联
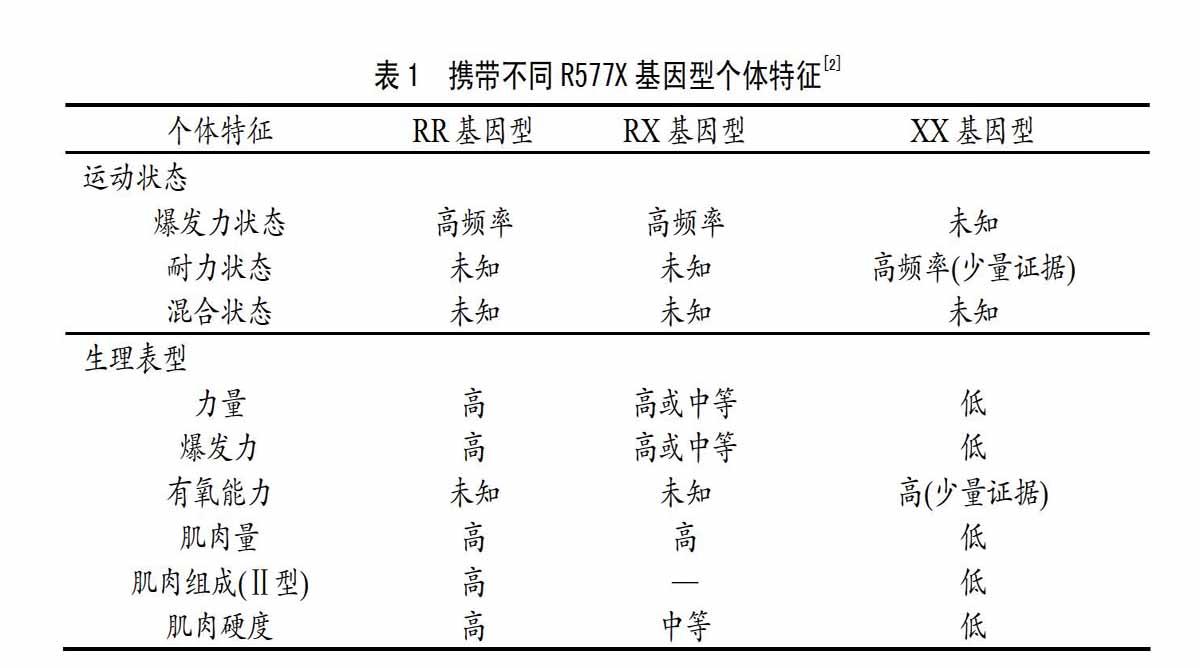
ACTN3基因R577X位点与优秀运动能力间的关联研究始于2003年。Yang等[11]在研究中发现,该多态位点的XX基因型在具有优秀爆发力素质的运动员中分布频率显著低于优秀耐力运动员和对照组,而此后研究者在世界各种族中均反复验证了R等位基因和RR基因型与优秀速度、爆发力及力量素质间的关联[2,9],并涉及了一系列的肌肉相关生理表型[2](见表1)。而根据当前文献,R577X多态性對运动能力外的多种运动相关表型也具有潜在影响,以下分别进行概述。
2.1 R577X多态性与运动适应性
业已证实,个体间的运动能力与运动适应性差异均受遗传影响。然而,相较于运动能力,个体运动适应性的遗传学研究进展较慢,截至目前,仅5项研究对R577X多态性与运动适应性间的关联进行了探索,其中4项研究为抗阻训练[12-15],1项研究为有氧训练[4]。结果发现,在抗阻训练研究中,R577X多态性对个体抗阻训练后适应性的影响程度不尽相同。其中,Erskine等[15]在对51名无训练经历年轻男性施加9周单边膝伸肌抗阻训练后发现,ACTN3基因型对抗阻训练适应性无影响;而Clarkson等[12]发现,在对非支配臂进行12周渐进式肘屈-伸抗阻训练后,女性中X等位基因组比RR基因型组具有更好的最大肌力提高量;Delmonico等[13]和Pereira等[14]则在分别对不同老人群体施加10周单边膝伸肌力量训练和12周高速爆发力训练后发现,RR基因型组的爆发力和力量增加比XX基因型组更多。究其原因,上述结果间的差异可能既与各研究所采用的运动训练方案不同有关,又与不同基因型基线水平间的异质性较大有关。但鉴于以往研究中R等位基因多见于兼备优秀速度、爆发力和力量素质的运动员中,因此有理由相信,该等位基因携带者会对速度、爆发力和力量训练产生较好的适应,这一方面可能是由于R等位基因携带者在爆发力训练中会发生更多的mTOR和p70S6k磷酸化,从而诱导骨骼肌肥大[16],另一方面则可能与R等位基因携带者具有更高的睾丸激素水平相关[17]。
再看R577X多态性对耐力训练后吸氧量提高水平的影响,可见18周耐力训练后,206名男性新招募警察中XX基因型组的基线吸氧量测试分数更高。然而,这一差异在训练后即消失,说明RR基因型携带者在训练后吸氧量提高幅度更大,也就是具有更好的运动适应性[4]。尽管此前大量研究均已证实X等位基因可用于杰出耐力运动员的选材,但由于在该研究中的实验对象是新招募的警察,故目前难以确定该结论能否反过来适用于优秀耐力运动员,因此接下来需开展更多的耐力性训练研究,以探索该多态性与训练后运动员有氧能力提高间的确切关联。
2.2 R577X多态性与运动恢复
除了运动适应性,现有研究结果还发现R577X多态性与运动引起的肌肉损伤相关[18-20]。由于在实践中,运动员肌肉损伤的增多会减慢其恢复速度,因此,研究者推测该多态性可能对运动后的恢复具有潜在影响。在迄今为止的8项有关该位点与运动后恢复的研究中,6项研究结果显示,X等位基因和(或)XX基因型携带者的肌肉损伤水平更高[6,18-22],1项研究发现该多态位点对运动恢复无影响[23],1项研究则称训练后RR携带者比XX基因型携带者力量降低更多[24]。
尽管上述6项研究既包括离心收缩训练又包括长时间的耐力运动,但结果均较为一致,即X等位基因和(或)XX基因型与运动后更高的肌肉损伤表征指标水平相关。如前所述,ACTN3仅特异性表达于快肌纤维,且XX基因型会导致ACTN3的表达缺失,故快肌纤维中的 ACTN2会上调表达以补偿ACTN3的功能缺失[9]。但是,因ACTN2和ACTN3均为骨骼肌内Z线的主要结构成分,Z线本身在离心收缩中又极易受损,而基因敲除小鼠模型也证实ACTN2的浓度增加会降低Z线在肌肉收缩中的稳定性[25],由此可以推测,运动后XX基因型携带者肌肉损伤的增多和恢复速度的下降主要是蛋白结构改变的结果。但要强调的是,上述研究结果涉及的实验对象样本量均较低(总数376名,平均47名),其中样本量最高的研究又发现R577X多态性与训练后的肌肉损伤无关[23],因此上述结果能否推广尚不明确。另外,由于人群中XX基因型的分布频率较低,而上述研究中XX基因型最多仅48名,且结果未能证实该多态性与训练后肌肉损伤的关联,故未来需进一步增加实验对象样本量以明确该多态性对运动恢复的确切影响。
2.3 R577X多态性与运动损伤风险
关于R577X多态性与运动损伤风险间的关联,迄今为止共有6項相关研究,其中3项聚焦于踝关节扭伤[5,26-27],另3项则分别研究了非接触性损伤[28]、职业足球运动员的损伤情况[29]和劳累性横纹肌溶解症(exertional rhabdomyolysis,ER)[30]。在上述研究中,多数研究者发现,R等位基因和(或)RR基因型具有抗损伤保护效应[5,26-29],而唯一与上述结果相悖的研究则报道称,女性中R等位基因携带者比X等位基因携带者具有更高的肌肉损伤风险[28]。分析可知,ER的产生机制其实与运动后肌肉损伤的发生机制十分类似,均为运动中Z线的损伤增多,而踝关节扭伤的发病机制则可能与肌肉功能相关,即由于R等位基因携带者具有更高水平的肌肉量(尤其是Ⅱ型肌纤维),因此更易于获得力量素质[9]。而综合前面研究中R等位基因具有抗损伤保护效应的发现可知,R等位基因的这种抗损伤保护效应其实是该等位基因在力量素质及运动后肌肉损伤方面效应的综合体现[3],至于在女性中发现的相反结果,则需更多研究进一步确认。
3 ACTN3基因R577X多态性与机体衰老间的关联
随着人类寿命的延长和老龄化时代的来临,人们因机体衰老导致的运动受限情况越来越多,不仅降低了个人的生活品质,也造成了社会医疗开支的大幅攀升。健康受多种因素影响,因此,当前的研究热点多集中于老年病预防与老龄期身体机能维持领域,一系列与健康性衰老相关的身体机能指标被纳入研究,如何维持肌肉力量则是其中的重要内容。鉴于R577X多态性与优秀运动员的多种肌肉相关表型均显著相关(见表1),故研究者推测该多态性也可影响机体的健康性衰老。
3.1 R577X多态性与机体衰老过程中的肌肉量和肌肉功能
伴随年龄增长,骨骼肌的肌肉量和功能均会下降,即临床上的肌少症。事实上,该过程并非始于中、老年,而是从约25岁时就已开始,到40岁时可减少肌肉量峰值净重的10%,70岁时则可减少40%[31],从而导致老年人跌倒风险的上升和机体整体机能的衰退。迄今为止,有多项涉及R577X多态性与老年人肌肉量及功能间关联的研究出现,多数结果认为R577X多态性对老年人的肌少症风险及维持老年人肌肉功能、肌肉量具有潜在协调作用,R等位基因携带者的肌少症患病风险更低而肌肉力量及功能更好[32-37],少数研究则未发现该多态与老年人肌肉相关表型及功能间存在关联[38-39]。
为何老年R等位基因携带者具有更低的肌少症风险和更高的肌肉力量与肌肉功能?分析可知,其背后机制源于该等位基因在Ⅱ型肌纤维中的表达量较高[33-34]。已知Ⅱ型肌纤维的萎缩是肌少症的特征之一,而携带R等位基因的老人因更利于维持快肌纤维的大小与数量,从而不仅肌肉功能更好,跌倒与肌少症的风险也更低。此外,鉴于抗阻训练是目前预防肌少症的一项重要手段,而前述研究已证实R577X多态性会影响受试者的运动适应性[12-15],即抗阻训练后,老年人中的R等位基因、特别是RR基因型携带者不仅肌肉力量更好,爆发力提高效果也更显著[13-14],故由此可知,该多态与老年人的抗阻训练适应性同样相关,而R等位基因携带者具有更好的抗阻训练适应性。
3.2 R577X多态性与机体衰老过程中的骨密度
低骨密度水平通常与全因死亡率、中风死亡及骨折风险的增加相关。因此,对老人而言,骨密度下降与骨质疏松的风险远比肌肉量下降及肌肉功能缺失危害性更大,后果也更严重[31]。截至目前,3项研究对老年人中R577X多态与骨密度缺失间的关系进行了探索,结果表明,R等位基因是抗老龄性骨密度缺失的保护性遗传标记[37,40-41]。对于该效应的作用机制,目前存在两种解释。第一种解释发现,握力及增加的肌肉量均与骨密度成正相关[7],因此,表达R等位基因所导致的肌肉量及力量的增加可保护其携带者不致出现骨密度缺失。而由于老年人中该等位基因携带者往往肌肉量与肌肉功能更好,故他们在生活中更乐于运动,日常骨架的承重也更多,从而更利于其骨骼结构的维持,骨密度缺失也更少[31]。另一种解释发现,缺失ACTN3的基因敲除小鼠骨密度更低,且ACTN3可表达于骨组织并参与骨质疏松的发生,而与此同时,小鼠的成骨细胞数下降,成骨细胞活动则增强[40]。鉴于上述两种机制可分别较好地解释R等位基因及XX基因型对骨密度的影响,因此可以认为,R577X位点不同基因型老人中骨密度缺失间的个体差异其实可能是上述两种机制共同作用的结果。
3.3 R577X多态性与机体衰老过程中的能量代谢水平
除了肌肉量与骨密度下降,一系列的代谢相关疾病,如胰岛素抵抗和2型糖尿病,也多发于老年人中。尽管目前全球此类疾病的致死率持续下降,但它们却可引起未来健康风险的增加与认知能力的下降[31]。鉴于高水平的肌肉量通常与较好的胰岛素敏感性相关[32],而R等位基因携带者又通常具有更大的肌肉横截面积和较好的肌纤维类型[36],因此研究者推测R577X多态性对老年人的2型糖尿病风险可发挥直接或间接的作用。当前,涉及此领域的研究数量有限,但研究者已发现,2型糖尿病人中的XX基因型分布频率显著高于对照组[42],表明X等位基因可提高老年人的2型糖尿病风险。究其原因,研究者认为这与XX基因型可潜在增加脂肪酸氧化和糖原储存,从而导致骨骼肌的代谢改变有关,并在ACTN3基因敲除小鼠中得到了验证[43],但遗憾的是,此结果目前并未在人类中进行检验,故该多态是否与老年人2型糖尿病的患病风险相关,仍需进一步明确。
除2型糖尿病外,研究者认为ACTN3基因还可通过造成其他代谢紊乱来影响健康。在2017年的一项鼠模型研究中,研究者发现XX基因型具有抗肥胖的保护效应[8],但在后续人类研究中未发现上述关联[8,44];而与此类似,有研究报道称,年轻人中的X等位基因可正向影响其他健康指示指标(如血压[45]和HDL-C[46]),但上述结果在老年人中是否具有同样适用目前仍需进一步检验。
综上所述,迄今为止已有大量研究证实了遗传对运动员优秀运动能力的影响,而其中,ACTN3基因的R577X多态是研究的热点之一。但事实上,大量研究表明该多态性不仅可影响速度、爆发力和力量素质,还可影响其他的运动相关表型及老年人的肌肉、骨骼和糖、脂代謝情况,其中RR基因型与较好的运动后适应性、更低的训练后肌肉损伤水平、较低的运动损伤风险、老年人中较好的肌肉功能、骨密度及潜在代谢健康水平相关,XX基因型则各项表型均较差。因此,尽管当前该基因的多态研究仍主要集中于优秀运动能力方面,在前述领域中开展研究较少且结论时有不同,但鉴于上述研究结果一旦确认即可用于未来运动员训练方案的个性化实施和老年病的个性化防治,从而提高运动员的训练效果和老年人的生活品质,故未来应在上述领域投入更多的资源与关注,并对当前研究中的各个未确定结论逐一加以确认,而本文则旨在抛砖引玉,为我国相关研究的开展提供参考和新的视角。
参考文献:
[1] SARZYNSKI M A,LOOS R J,LUCIA A,et al. Advances in exercise,fitness,and performance genomics in 2015[J]. Med Sci Sports Exerc,2016,48(10):1906-1916.
[2] KIKUCHI N,NAKAZATO K. Effective utilization of genetic information for athletes and coaches:focus on ACTN3 R577X polymorphism[J]. J Exerc Nutrition Biochem,2015,19(3):157-164.
[3] PICKERING C,KIELY J. ACTN3:More than Just a Gene for Speed[J]. Front Physiol,2017,8:1080.
[4] SILVA M S,BOLANI W,ALVES C R,et al. Elimination of influences of the ACTN3 R577Xvarianton oxygen uptake by endurance training in healthy individuals[J]. Int J Sports Physiol Perform,2015,10(5):636-641.
[5] QI B,LIU J Q,LIU G L. Genetic association between ACTN3 polymorphism and risk of non-acute ankle sprain[J]. Genet Mol Res,2016,15(4).gmr15048962.
[6] BELLI T,CRISP A H,VERLENGIA R. Greater muscle damage in athletes with ACTN3 R577X (RS1815739) gene polymorphism after an ultra-endurance race:a pilot study[J]. Biol Sport,2017,34(2):105-110.
[7] IIDA T,IKEDA H,SHIOKAWA M,et al. Longitudinal study on physical fitness parameters influencing bone mineral density reduction in middle-aged and elderly women:bone mineral density in the lumbar spine,femoral neck,and femur[J]. Hiroshima J Med Sci,2012,61(2):23-28.
[8] HOUWELING P J,BERMEN Y D,TURNER N,et al. Exploring the relationship between α-actinin-3deficiency and obesity in mice and humans[J]. Int J Obes (Lond),2017,41(7):1154-1157.
[9] 楊若愚,王予彬,沈勋章,等. 基因多态性与杰出运动能力[J]. 中国组织工程研究,2014,18(7):1121-1128.
[10] VINCENT B,DE BOCK K,RAMAEKERS M,et al. ACTN3 (R577X) genotype is associated with fiber type distribution[J]. Physiol Genomics,2007,32(1):58-63.
[11] YANG N,MACARTHUR D G,GULBIN J P,et al. ACTN3 genotype is associated with human elite athletic performance[J]. Am J Hum Genet,2003,73(3):627-631.
[12] CLARKSON P M,DEVENEY J M,GORDISH-DRESSMAN H,et al. ACTN3 genotype is associated with increases in muscle strength in response to resistance training in women[J]. J Appl Physiol (1985),2005,99(1):154-163.
[13] DELMONICO M J,KOSTEK M C,DOLDO N A,et al. Alpha-actinin-3 (ACTN3) R577X polymorphism influences knee extensor peak power response to strength training in older men and women[J]. J Gerontol A Biol Sci Med Sci,2007,62(2):206-212.
[14] PEREIRA A,COSTA A M,IZQUIERDO M,et al. ACE I/D and ACTN3 R/X polymorphisms as potential factors in modulating exercise-related phenotypes in older women in response to a muscle power training stimuli[J]. Age (Dordr),2013,35(5):1949-1959.
[15] ERSKINE R M,WILLIAMS A G,JONES D A,et al. The individual and combined influence of ACE and ACTN3 genotypes on muscle phenotypes before and after strength training[J]. Scand J Med Sci Sports,2014,24(4):642-628.
[16] NORMAN B,ESBJ?RNSSON M,RUNDQVIST H,et al. ACTN3 genotype and modulation of skeletalmuscle response to exercise in human subjects[J]. J Appl Physiol (1985),2014,116(9):1197-1203.
[17] AHMETOV I I,DONNIKOV A E,TROFIMOV D Y. Actn3 genotype is associated with testosterone levels of athletes[J]. Biol Sport,2014,31(2):105-108.
[18] VINCENT B,WINDELINCKX A,NIELENS H,et al. Protective role of alpha-actinin-3 in the responseto an acute eccentric exercise bout[J]. J Appl Physiol (1985),2010,109(2):564-573.
[33] KIKUCHI N,YOSHIDA S,MIN S K,et al. The ACTN3 R577X genotype is associated with muscle function in a Japanese population[J]. Appl Physiol Nutr Metab,2015,40(4):316-322.
[34] JUDSON R N,WACKERHAGE H,HUGHES A,et al. The functional ACTN3 577X variant increases the risk of falling in older females: results from two large independent cohort studies[J]. J Gerontol A Biol Sci Med Sci,2011,66(1):130-135.
[35] WALSH S,LIU D,METTER E J,et al. ACTN3 genotype is associated with muscle phenotypes in women across the adult age span[J]. J Appl Physiol (1985),2008 Nov,105(5):1486-1491.
[36] ZEMPO H,TANABE K,MURAKAMI H,et al. ACTN3 polymorphism affects thigh muscle area[J]. Int J Sports Med,2010,31(2):138-142.
[37] CHO J,LEE I,KANG H. ACTN3 Gene and Susceptibility to Sarcopenia and OsteoporoticStatusin Older Korean Adults[J]. Biomed Res Int,2017:1-8.
[38] MCCAULEY T,MASTANA S S,FOLLAND J P. ACE I/D and ACTN3 R/X polymorphisms and muscle function and muscularity of older Caucasian men[J]. Eur J Appl Physiol,2010,109(2):269-277.
[39] LIMA R M,LEITE T K,PEREIRA R W,et al. ACE and ACTN3 genotypes in older women muscular phenotypes[J]. Int J Sports Med,2011,32(1):66-72.
[40] MIN S K,LIM S T and KIM C S. Association of ACTN3 polymorphisms with BMD,and physical fitness of elderly women[J]. J Phys Ther Sci,2016,28:2731-2736.
[41] YANG N,SCHINDELER A,MCDONALD M M,et al. α-Actinin-3 deficiency is associated with reduced bone mass in human and mouse[J]. Bone,2011,49:790-798.
[42] RIEDL I,OSLER M E,BENZIANE B,et al. Association of the ACTN3 R577X polymorphism with glucose tolerance and gene expression of sarcomeric proteins in human skeletal muscle[J]. Physiol Rep,2015,3(3). pii: e12314.
[43] MACARTHUR D G,SETO J T,RAFTERY J M,et al. Loss of ACTN3 gene function alters mouse muscle metabolism and shows evidence of positiveselection in humans[J]. Nat Genet,2007,39(10):1261-1265.
[44] MORAN C N,YANG N,BAILEY M E,et al. Association analysis of the ACTN3 R577X polymorphism and complex quantitative body composition and performance phenotypes in adolescent Greeks[J]. Eur J Hum Genet,2017,15:88-93.
[45] DESCHAMPS C L,CONNORS K E,KLEIN M S,et al. The ACTN3 R577X Polymorphism isassociated with cardiometabolic fitness in healthy young adults[J]. PLoS One,2015,10(6):e0130644.
[46] NIRENGI S,FUJIBAYASHI M,TSUZAKI K,et al. ACTN3 gene R577X polymorphism associated with high-density lipoprotein cholesterol and adiponectin in rugby players[J]. Endocr Pract,2016,22(7):786-790.
