发育中小鼠大脑皮层GABA能神经元能障变化及对神经编码的影响
2018-12-28葛荣靖倪虹申林王其一
葛荣靖 倪虹 申林 王其一
摘 要:目的 探讨发育过程中能障变化规律及对小鼠大脑皮层GABA能神经元编码能力的影响。方法 取生后6-12天(未睁眼)、15-22天(睁眼)小鼠,孵育脑片并用膜片钳记录皮层GABA能神经元电的生理特性。使用Clampfit10软件记录动作电位的能障(potential barrier, PB)、动作电位间距(inter-spike intervals, ISI),并分析结果。结果 发育过程中,小鼠大脑皮层GABA能神经元能障减小(P<0.01)、ISI缩短(P<0.01)。 PB与ISI之间始终呈线性相关。结论 小鼠大脑皮层GABA能神经元在生后发育过程中,发放动作电位的能障逐渐降低,神经元编码能力上调。
关键词:发育;GABA能神经元;能障;全细胞记录
γ-氨基丁酸(GABA)是脑皮质中的主要抑制性神经递质,由GABA能神经元合成并分泌,分布于大脑皮质各个区域[1]。GABA能神经的活动对脑功能具有重要的调节作用。在一些研究中,通过对神经元动作电位参数的量化分析来研究其编码特性,如动作电位间距(inter-spike intervals,ISI),绝对不应期(absolute refractory period, ARP)、能障(potential barrier, PB)等[2,3,4,5]。本课题组之前研究了小鼠大脑皮层GABA能神经元动作电位在生后发育过程中的变化规律[6]:在发育过程中,皮层GABA能神经元动作电位的ISI缩短,ARP的缩短是重要的影响因素。这些变化使GABA能神经元的兴奋性逐渐增加。
本研究将对皮层GABA能神经元兴奋性发育做进一步探索。通过改变去极化刺激强度(阈刺激强度×0.7, ×0.85, ×1, ×1.25)诱发动作电位,通过预设刺激模式,当前一个动作电位产生并完全恢复后再施加与前次相等的去极化电流刺激,以此产生5组动作电位,分别各采样20次,获得5组连续动作电位群组,采用全细胞记录方法,探讨小鼠发育过程中内在特性的变化及对动作电位编码的影响,为GABA能神经元发育的机制研究提供新的思路。
1 材料与方法[7]
1.1 实验材料及实验用液组成
1.1.1药品:
Hepes(Sigma);Mg2ATP(Sigma);EGTA(Sigma);Tris2GTP(Sigma);Kclu(Sigma);42Na2phosphocreatine(Sigma)。
1.1.2实验动物选择:
蚌埠医学院实验动物中心提供转基因FVB-Tg(Gad GFP)45704 Swn/J小鼠,使用许可证【SYXK(皖2012-002)】。
1.1.3人工脑脊液(mmol/L)
3 KCl, 124 NaCl, 26 NaHCO3, 1.3NaH2PO4, 5 HEPES, 10 dextrose, 0.5 CaCl2和4 MgSO4 (95%O2和5%CO2充分氧合)。
1.1.4 电极标准液(mmol/L)
4 NaCl, 150 葡萄糖酸钾, 0.5 EGTA, 10 HEPES, 1 Tris-GTP, 4 Mg2+-ATP (pH调至7.35;渗透压295~305 mOsmol)。
1.2实验方法
选用生后6-12天、15-22天的FVB-Tg(Gad GFP)45704 Swn/J小鼠,剪刀断头,快速剥离大脑,转移至充分氧合的人工脑脊液,用振荡切片机制作脑片标本约3-5片(400μm)。将脑片移至装有充分氧合的人工脑脊液的培养皿中,保持环境温度约35℃,孵育1小时[5]。 随后将脑片转移至IR-DIC光学显微镜下的记录槽中,小槽内预先充灌35?C氧合的人工脑脊液,灌流速度约为40-50滴/分。通过全细胞记录[5](Axoclamp-2B 放大器,电流钳模式,3 kHz高频滤波),pClamp 10记录信号并进行分析。
通过Clampfit10软件设定刺激脉冲:5 ms去极化电流,逐步增加电流强度,至50%可能性产生动作电位时,此电流值设为阈刺激。在阈刺激基础上,改变去极化电流强度(阈刺激强度×0.7, ×0.85, ×1, ×1.25)诱发动作电位。为了使阈刺激强度×0.7, ×0.85组可以正常诱发动作电位,故将阈强度预设为阈刺激强度×0.7组的刺激强度,反推出其他组刺激强度,获得相应的动作电位波形,应用Clampfit分别对动作电位参数(PB、ISI值)进行测定(测量方法见后文述)。
1.3 实验分组
小鼠出生后6-12天由于眼睛未振凯,设为未睁眼组;15-22天眼睛睁开,设为睁眼组[6]。
1.4PB和ISI的測定
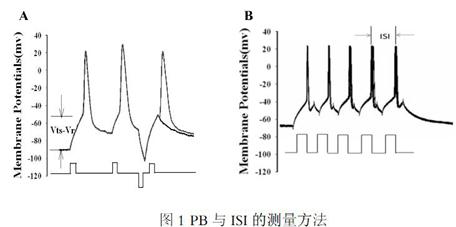
能障(potential barrier, PB):分别测得阈电位(threshold potential, Vts)及静息电位(resting membrane potential, Vr ),其差值即为PB[2]。[图1(A)]。
动作电位间距(ISI):为相邻两动作电位峰值间的距离,其值与神经元产生动作电位的容量成反比。[图1(B)]
1.5控制指标
在获得的数据中,只选用静息电位低于-65 mV的记录纳入统计分析。静息膜电位、动作电位幅度和输入阻抗的波动范围在5%之内[8]。保持电极阻抗为5-6 M。
1.6 数据处理
实验数据以均数±标准误( ±SE)表示,SPSS19.0统计软件做方差分析,组间比较采用t检验, P <0.05为有统计学差异。
2、结果
2.1 GABA能神经元能障在发育中的变化
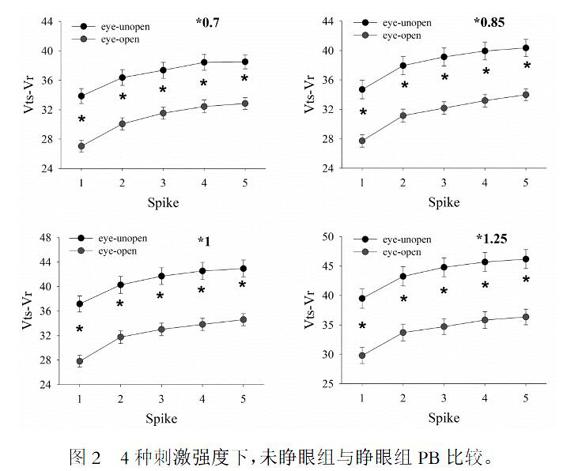
小鼠睁眼前后分别记录大脑皮层GABA能神经元的序列动作电位。结果显示,不同强度去极化脉冲所诱发的GABA能神经元的序列动作电位的PB在两组间均存在显著差异(P<0.01)。给予不同强度的刺激,睁眼组产生动作电位的PB均较未睁眼组减小[图2]。
(A)在大脑皮层GABA能神经元测量能障的方法。测量阈电位(threshold potential, Vts)和静息电位(resting potential,Vr ),其差值即为PB[2];(B)动作电位间距(ISI)的测量方法。向细胞内输入电流刺激神经元产生动作电位,测量相邻动作电位峰值间的时间差 [6]。
在STi×0.7, ×0.85, ×1, ×1.25四种电流刺激强度下,睁眼组较未睁眼组Vts-Vr1、Vts-Vr2、Vts-Vr3、Vts-Vr4、Vts-Vr5均减小。*P<0.01,睁眼组与未睁眼组比较。
2.2 发育对GABA能神经元动作电位间距(ISI) 的影响
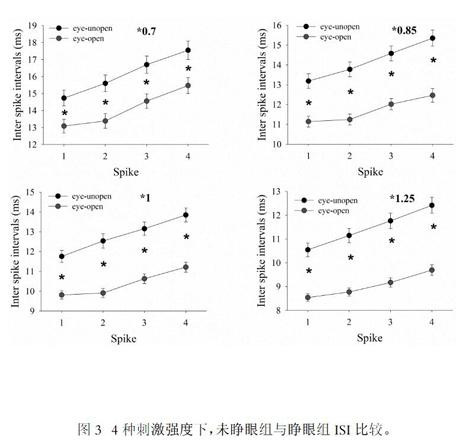
小鼠发育中,分别对睁眼前及睁眼后小鼠进行电生理记录。随着刺激强度变化,大脑皮層GABA能神经元发放的动作电位对应的ISI存在差异性,睁眼组较未睁眼组ISI缩短(P<0.01)。(图3):
2.3.发育中能障(PB)与动作电位间距(ISI)之间的关系
量化内在特性和动作电位编码间关系。未睁眼组PB和ISI之间具有线性先关。四组刺激中相关系数分别为:sti×0.7(r2=0.92),sti×0.85(r2=0.96), sti×1(r2=0.81),sti×1.25(r2=0.78)。睁眼组的PB和ISI之间也具有线性相关。四组刺激中相关系数分别为:sti×0.7(r2=0.91),sti×0.85 (r2=0.99),sti×1 (r2=0.92,P),sti×1.25 (r2=0.92)。(图4)
在STi×0.7, ×0.85, ×1, ×1.25四种电流刺激强度下,可见睁眼组较未睁眼组ISI1、ISI2、ISI3、ISI4均缩短。*P<0.01,睁眼组与未睁眼组比较。
(A)未睁眼组,不同刺激强度诱发的动作电位的PB与ISI之间线性相关。(B)睁眼组,不同刺激强度诱发的动作电位的PB与ISI之间线性相关。
讨论:
GABA是脑内重要的抑制性神经递质,在维持神经系统内环境稳态有重要意义。神经精神性疾病多与GABA能神经元活动异常有关[9,10,11]。本研究对中枢神经系统GABA能神经元动作电位的内在特性及编码模式进行研究,探讨小鼠大脑皮层GABA能神经元的发育规律。
本实验中量化研究了不同发育阶段、不同刺激强度下的小鼠GABA能神经元的能障(PB)、动作电位间距(ISI)。研究发现,在发育过程中动作电位参数均呈动态变化,主要表现在①发育过程中,GABA能神经元动作电位的能障缩短,意味着静息电位水平或最大复极电位水平与阈电位水平之间的差距减小,神经元兴奋性增加。能障主要反映了GABA能神经元神经元阈电位水平,实质是Na+通道电压依赖性的变化。能障减小,阈电位水平下移可使神经元兴奋性增加。②发育中ISI缩短意味着单位时间内发放的动作电位个数增加,是GABA能神经元兴奋性增加的结果,提示动作单位的编码能力上调。编码能力上调是情绪、学习、记忆等行为的神经信号基础,也是动作电位的编码重要形式[12-13]。③未睁眼组与睁眼组中,能障(PB)始终与ISI之间存在线性相关,提示这是一个动态的控制过程。PB通过什么机制调控ISI仍需进一步进行研究。
综上,在发育过程中GABA能神经元动作电位的能障(PB)降低,使GABA能神经元兴奋性增加;GABA能神经元动作电位间距增加,使神经元在单位时间内发放动作电位的个数增加编码信号的能力增强。本研究进一步探讨了小鼠大脑皮层GABA能神经元动作电位的发育规律,明确了能障与ISI之间的关系,为了解GABA能神经元对输入信息的编码机制及发育规律提供一些理论依据。
参考文献:
[1] 韩济生.神经科学原理[M].北京,北京医科大学出版社,1999:539-551.
[2] Guan S, Ma S, Zhu Y, et al.The intrinsic mechanism sunderlying the maturation of programming sequential spikes at cerebellar Purkinje cells[J].Biochemical and Biophysical Research Communication, 2006,345 (1):175–180.
[3] Chen N, Zhu Y, Gao X,et al. Sodium channel2 mediated intrinsic mechanisms underlying the differences of spike programming among GABAergic neurons[J]. Biochemical and Biophysical Research Communication,2006,346 (1) :281–287
[4] Chen N, Chen S, Wu Y, et al. The refractory periods and threshold potentials of sequential spikes measured by whole-cell recording[J].Biochemical and Biophysical Research Communication,2006,340 (1): 151–157.
[5] Wang J.H, Wei X.J, Chen J, et al.,Gain and ?delity of transmission patterns at cortical excitatory unitary synapses improves spike encoding[J].Journal of Cell Science, 2008,121 (17):2951–2960.
[6] 王其一,申林.发育中小鼠大脑皮层GABA能神经元电生理特性的变化[J].上海交通大学学报(医学版),2016, 36(4):501-506.
[7] Lei Jiang, Hong Ni, Qi-yi Wang,et al.Dual face of axonal inhibitory inputs in the modulation of neuronal excitability in cortical pyramidal neurons.2017,12(7):1079-1085
[8] Wang Q, Liu X, Ge R,et al.The postnatal development of intrinsic properties and spike encoding at cortical GABAergic neurons[J].Biochemical and Biophysical Research Communication, 2009,378(4): 706-710.
[9] Milosevic L,Kalia SK,Hodaie M et al. Neuronal inhibition and synaptic plasticity of basal ganglia neurons in Parkinson's disease. [J]. Brain, 2018,141(1):177-190.
[10] De Jonge JC, Vinkers CH, Hulshoff Pol HE,et al. GABAergic Mechanisms in Schizophrenia: Linking Postmortem and In Vivo Studies. [J].Front Psychiatry. 2017.118(8): eCollection.
[11] Delattre V, Keller D, Perich M,et al. Network-timing-dependent plasticity[J]. Front Cell Neurosci. 2015, 9:220.
[12] Shidi Zhao, Na Chen, Zhilai Yang,et al. Ischemia deteriorates the spike encoding of rat cerebellar Purkinje cells by raising intracellular Ca2+[J].Biochemical and Biophysical Research Communications. 2008,366:401-407
[13] Spaak E, Watanabe K, Funahashi S,et al.Stable and Dynamic Coding for Working Memory in Primate Prefrontal Cortex. J Neurosci. 2017 37(27):6503-6516.
作者简介:
葛荣靖(1982-),女,安徽省蚌埠市人,民 族:汉 职称:讲师,学历:博士研究生。研究方向:神经信息编码的细胞与分子机制。
基金项目:安徽省自然科学基金(1608085QH176);安徽高校省级自然科学研究项目(KJ2012B105);蚌埠醫学院科学研究重点项目(BYKY1623ZD).
