两种真菌接菌结香对白木香木质部组织构造的影响
2018-12-27谷丽萍郑科刘玉蓉马惠芬肖支叶
谷丽萍,郑科,刘玉蓉,马惠芬,肖支叶
(1.云南省林业科学院,云南 昆明 650201;2.西南林业大学,云南 昆明 650224)
沉香十分珍贵,但是天然结香“百无一二”,导致人工结香逐渐盛行起来。由于真正高效的结香技术却几近空白,人们开始探索快速、高效的人工结香技术。人工结香主要集中在3个方面,一是物理诱导结香(半断干、砍伤及凿洞等技术),二是化学诱导(氯化钠、亚硫酸氢钠、氯化亚铁等药剂),三是真菌诱导〔黄绿墨耳菌(Melanotusflavolivens)〕等[1]。本项研究使用国内普遍采用且比较有效的结香真菌[2]可可毛色二孢(Lasiodiplodiatheobromae)与腐皮镰孢菌(Fusariumsolani)(沉香出产大国印尼采用的结香真菌)开展了白木香(Aquilariasinensis)接菌结香试验,对白木香树枝接菌前后木质部的显微构造进行观察,确定白木香树枝中形成的沉香树脂的组织分布范围以及在荧光下观察被真菌侵染与未被真菌侵染的白木香内含韧皮部的胼胝质沉积情况,以探讨真菌接种白木香的结香机理,为白木香人工结香提供参考。
1 实验材料与方法
1.1 实验材料与设备
(1)液体菌剂制备与滴注 在无菌条件下,将扩繁后的菌液分别用已灭菌的单层医用纱布过滤,弃掉培养基,留下滤液。此滤液中含有真菌的菌丝体。将可可毛色二孢和腐皮镰孢菌体分装到预先灭菌并称重的离心管中,室温下10 000rpm离心3min,倒弃上清液,保留沉淀物,并称量离心后的离心管总重,计算出菌体湿重。由于实验条件所限,在超净工作台上分别称可可毛色二孢1g、3g(记为L1、L3),腐皮镰孢菌2g、3g、4g(记为F2、F3、F4),混菌(每个菌0.5g、1g,记为H1、H2)转入新的离心管中,在组织研磨仪中研碎。之后分别用少量纯净水将不同湿重的菌溶分装到贴有标签的输液袋(含输液管、输液针头和流速调节装置)中,再加入1 000mL纯净水制成不同浓度的液体菌剂。
在景洪市勐海县勐宋乡广顺沉香基地,选取白木香树,在白木香较大树干10-50cm部位,用电钻在树枝两侧钻出深约5cm的输液孔(直径5mm),输液孔与地面成30°-45°夹角。输液袋挂在高于接菌处的树枝上,将输液针头插入输液孔,调节流速至营养液流动但不外溢,将各处理液体菌剂输入到白木香较大树枝中,其液约2d滴注完成。每个处理3株,即3次重复。
(2)实验材料 真菌侵染白木香树5月后,从接菌处用手工锯截断白木香树枝,再用砍刀取木质部结香处,将形成树脂的木质部作为样品。
(3)试剂与设备 1%番红染色剂,苯胺蓝染色液,正丁醇,二甲苯,体积分数50%、75%、95%乙醇,无水乙醇,聚乙二醇(分子量1 500),病理级载玻片与盖玻片,莱卡划走式切片机,砍刀,烘箱,尼康80i生物数码显微镜。
1.2 实验方法
(1)样品固定 对样品去掉腐木部分,样品含过渡带与白木部分。取样后将样品迅速放入FAA固定液(50%酒精90mL+福尔马林5mL+冰醋酸5mL)中固定。
(2)切片制备 a.包埋前用自来水将FAA固定液固定的样品冲洗5min,除去残留的FAA固定液。随后用体积分数为30%、50%、70%和100%的PEG依次包埋24h;b.样品包埋完成后,用莱卡划走式切片机进行切片,切片制作2份。将1份切片平展于载玻片上,同时用盖玻片封好,转入1%番红染色剂中染色2h。染色完成后,将样品依次放入体积分数为50%、75%、95%乙醇中脱水,每次脱水5min;c.将b中在95%乙醇中脱水完成的切片放入固绿染液复染1min,依次在无水乙醇和正丁醇中冲洗5min,然后在正丁醇、1/2正丁醇+1/2二甲苯和二甲苯中依次处理5min,完成复染后封片并于50℃烘箱中烘片24h;d.采用苯胺蓝染色法,将另外1份切片置于苯胺蓝溶液中,染色30min,取出平展于载玻片上,同时用盖玻片封好。用尼康80i生物数码显微镜(荧光组件)在紫外光激发下观察胼胝质沉积情况并拍照。
2 结果与分析
2.1 白木香树枝解剖构造
白木香接菌结香后相关解剖结构见图1。

图1未滴注和滴注H2菌剂5月后白木香的微观构造
注:1为未被真菌侵染的白木香树枝的横切面构造,没有沉香树脂
形成;2为未被真菌侵染的白木香树枝的弦切面构造,没有沉香树脂形成;3为未被真菌侵染的白木香树枝的径切面构造,没有沉香树脂形成;4为H2侵染后的横切面构造,图中导管壁、木射线和内含韧皮部中均形成沉香树脂;5为H2侵染后的弦切面构造,图中导管壁和木射线细胞中有沉香树脂;6为H2侵染后的径切面构造,图中木射线细胞中和内含韧皮部产生大量沉香树脂。
Fig.1 The microstructure ofA.sinensisafter 5 months treatment without or with microbial inoculums drip(H3)
由图1可以看出,白木香树枝生长轮不明显,散孔材。导管横切面为圆形或卵圆形,单管孔较多,多数导管常2-4个排列为径列复管孔或为管孔团。木纤维细长,壁薄。木射线非叠生,单列及多列,射线组织异型III型,少数II型,正常射线细胞不含树胶。内含韧皮部甚多,四周为薄壁组织,呈多孔型(岛型),细胞壁薄,在次生木质部内呈条带状或长椭圆状均匀分布。内含韧皮部中的木射线薄壁细胞和木纤维细胞,经番红固绿双染色后呈绿色。
2.2 白木香树枝沉香树脂分布
白木香树枝被结香真菌侵染5月后,接菌口上下木质部均发生病腐。从可可毛色二孢、腐皮镰孢菌和复合菌侵染白木香其树枝的显微构造能够看出,沉香树脂主要出现在导管、木射线薄壁细胞和内含韧皮部。从切片上观察发现,树脂刚产生时附着在导管壁和薄壁细胞壁上,随着侵染时间延长,树脂沉积在导管中,薄壁细胞则被树脂填充,最终失去生理活性。导管和木射线在木材中分别主导径向和横向输送养分的作用[3],薄壁细胞是边材储存养分的生活细胞,沉香树脂主要出现在导管、木射线薄壁细胞和内含韧皮部,可能与其输送和储存养分的功能有关联。在可可毛色二孢和腐皮镰孢菌侵染的白木香中,低浓度菌液处理条件下,树脂产生部位主要在导管壁上,木射线薄壁细胞和内含韧皮部薄壁细胞中树脂甚少。随着菌液浓度增大,到3g/L时此二者细胞中均形成了树脂。在混菌接种白木香处理条件下,菌液浓度为1g/L 和2g/L时(图1),白木香在导管、木射线细胞和内含韧皮部形成大量树脂,已看不清木射线薄壁细胞形态,薄壁细胞失去了生理活性。图1为未被真菌侵染的白木香树枝和复合菌剂H2侵染白木香5月后其树枝的解剖构造。
2.3 真菌侵染对白木香胼胝质沉积的影响
利用尼康80i生物数码显微镜在紫外光激发下,观察经苯胺蓝溶液染色的样品切片的胼胝质沉积情况并拍照。胼胝质分布情况见图2。
胼胝质(callose)是一种以β-1,3键结合的葡聚糖,围绕每个筛孔的边缘积累。胼胝质可及时合成和降解来应答外界的物理、化学和生物入侵引起的生物胁迫[4],它的合成能够增强植物的抗病性。通过对样品切片的荧光显微观察,发现白木香内含韧皮部中的胼胝质呈颗粒状。白木香产生沉香树脂的地方未见或有较少胼胝质分布,在过渡带部分,内含韧皮部四周的薄壁组织壁异常发亮,在F2、F3、F4、L1、L3、H1和H2处理方式下十分明显,内含韧皮部筛管中有较多胼胝质分布。还未产生沉香树脂的地方有较多的胼胝质沉积。在一个内含韧皮部中,可观察到两个及以上胼胝质,在荧光下呈黄绿色亮斑,处于过渡带的胼胝质色泽偏浅,可能是其参与白木香的抗病反应而被消耗。在组织感病附近,胼胝质沉积较多,说明白木香树在受到伤害的情况下,会在受害组织细胞及附近迅速沉积大量胼胝质以愈合伤口。本研究发现,无论何种处理方式,白木香内含韧皮部中的胼胝质均呈现相同的变化规律。图2为F2、F3、F4、L1、L3、H1、H2和未感菌的白木香横切面胼胝质分布图。
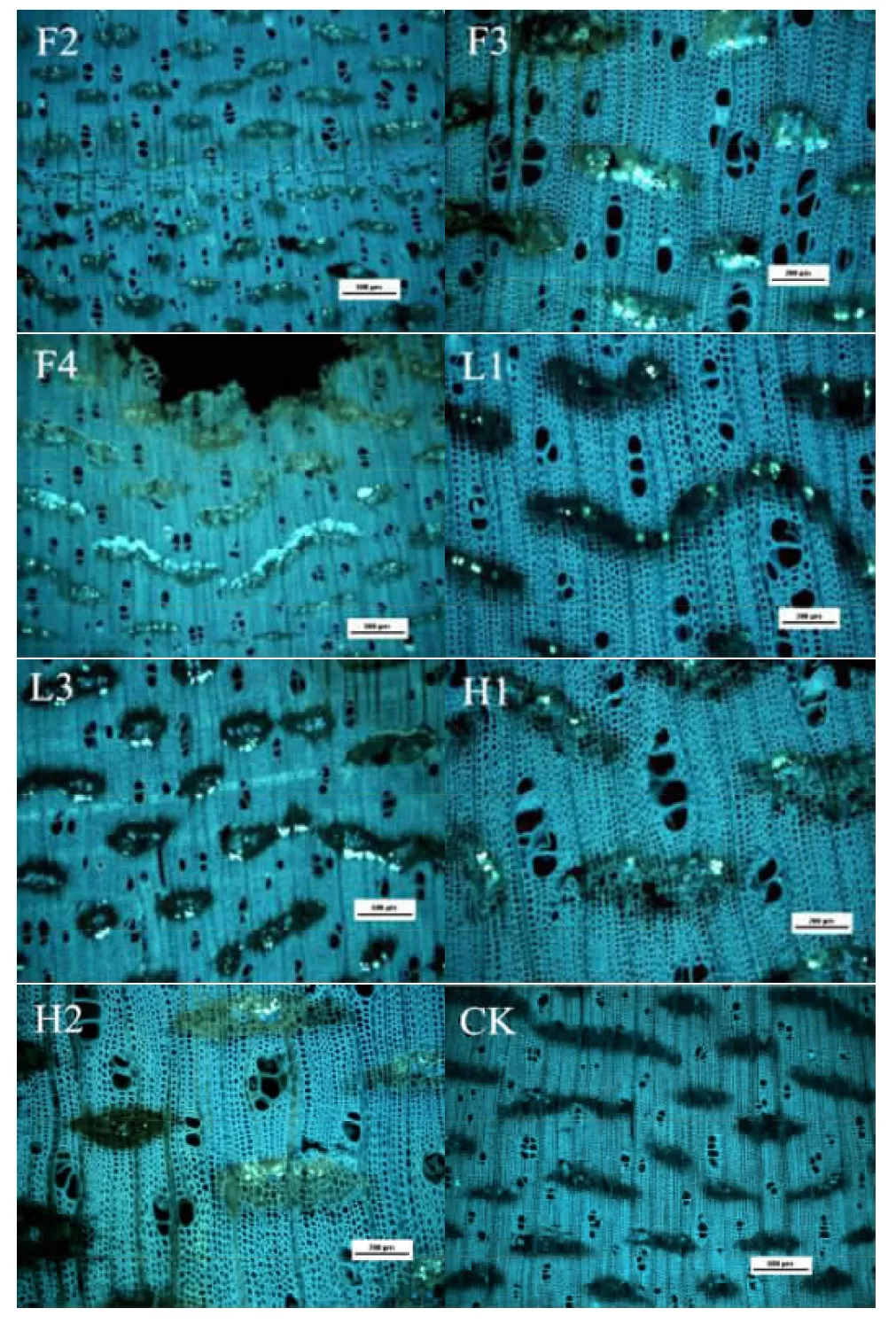
图2接菌和未接菌的白木香木材横切面的胼胝质分布
注:L1为可可毛色二孢1g;L3为可可毛色二孢3g;F2为腐皮镰
孢菌2g;F3为腐皮镰孢菌3g;F4为腐皮镰孢菌4g;H1为混菌,每个菌0.5g;H2混菌,每个菌1.0g。
Fig.2 Callose distribution on the cross section ofA.sinensisinoculated and un-inoculated fungi
2.4 真菌侵染结香机理分析
目前,关于白木香结香机制的研究仍然集中于真菌侵染诱导结香,机理尚不明确[5]。在自然条件下,植物经常处在生物与非生物胁迫之中。真菌侵染白木香的过程是病原菌对其施加生理胁迫的过程。在生理胁迫过程中,植物细胞会通过产生次生代谢物质来抵御胁迫。马华明[3]研究了2种结香效果不同的结香菌株[层出镰刀菌(Fusariumproliferatum)和红褐肉座菌(Hypocreajecorina)]对白木香组培幼苗的生理胁迫过程,通过比较它们之间的异同点来分析胁迫与结香之间关系。分析结香真菌与糖代谢的联系发现,真菌侵染后会引发植物可溶性糖的大幅增加,可能引发植物生长素的连锁反应。由此可知,植物体内的糖作为一种信号分子存在,和其它信号分子一起组成植物的复杂信号网络体系,增强植物对逆境胁迫的抵抗性。糖的代谢是整个植物代谢的中心,调节着植物的一系列生命活动。研究发现,植物对糖信号的感知和传导可在毫摩尔水平进行[6],这与植物体内的一种超细胞结构—胞间连丝[7]的存在有关。糖在沉香形成中或许具有重要作用。
本研究发现,在各处理条件下,在白木香组织发生病腐处形成树脂,在感病附近有胼胝质沉积,但沉积数量有差异。胼胝质的沉积受到β-1,3葡聚糖酶的控制[8],β-1,3葡聚糖酶活性存在差异,使胼胝质沉积速度不同。胼胝质在真菌侵染部位富集,会持续合成与分解,具有可逆性,胼胝质的合成与分解是一种糖代谢活动。与未感菌的白木香比较可知,感菌的白木香中胼胝质的代谢更为活跃,沉积量比健康的白木香多。胼胝质的沉积可改变筛管的运输功能,也会改变细胞及溶液的渗透势,使细胞中的水分含量发生变化,进而调节植物抗协迫环境的能力。真菌侵染白木香后,白木香树应答生物胁迫,在内含韧皮部沉积的胼胝质和其他信号分子进行代谢,这个过程中,信号分子可能与病原菌产生的毒素发生化学反应,以抵抗病原菌的病腐。或在胞间连丝处大量沉积,阻止病原菌产生的次生代谢产物(小分子物质)在细胞间流通,降低细胞中的水分含量,形成不利于病原菌生长的环境,从而起到抑制真菌生长的作用。
3 结语
不同接菌处理白木香树枝经过5个月后,其导管、木射线薄壁细胞和内含韧皮部组织中均会形成沉香树脂。在木质部形成沉香树脂阻塞木射线和内含韧皮部薄壁细胞,阻止水分迁移。结香真菌侵染白木香引起其组织局部坏死反应,进而促使胼胝质积累以应答病原真菌引起的植物逆境胁迫。在已形成沉香树脂的地方未见胼胝质沉积,在过渡带有许多胼胝质沉积。胼胝质沉积堵塞筛板,阻止水分迁移,胼胝质参与了防御反应以及沉香树脂的形成过程。糖在白木香沉香形成中或许具有重要作用。
人们只有在弄清结香机理,攻克结香技术的基础上,才会提高沉香产量,沉香树种植产业也才会得到大力发展。本次研究揭示了可可毛色二孢与腐皮镰孢菌侵染白木香树体后,会促使树体产生树脂以及胼胝质以抵御真菌的侵染,相应产生一些沉香类物质。但是真菌侵染损害树体如何导致沉香活性次生代谢物的产生涉及其各个机理仍然不清楚,此外,除真菌损伤外,其他损害(物理损伤、化学损伤)如何导致白木香树沉香类物质产生仍需进一步探讨。
