粪源环丙沙星对潮土中抗生素抗性基因的影响
2018-12-25常旭卉贾书刚王淑平周志强
常旭卉,贾书刚,王淑平*,周志强
(1.中国科学院大学资源与环境学院,北京 100049;2.广西师范学院,北部湾环境演变与资源利用教育部重点实验室,南宁530001)
抗生素在保障人类健康、促进畜禽养殖业发展等方面起重要作用。我国畜禽养殖较为发达,经预测,到2020年每年畜禽粪便的产量将达到42.4亿t[3],畜禽粪便农用造成的抗生素和抗生素抗性基因(Antibi⁃otic resistance genes,AGRS)的污染问题已经引起了广泛的关注。世界卫生组织(WHO)早在2000年的报告中就将抗生素抗药性列为本世纪人类在健康领域面临的重大挑战之一[5]。
畜禽养殖业长期使用兽用抗生素以促进动物生长、预防和治疗动物疾病等,但约25%~75%的抗生素未被机体吸收而随粪尿排出体外进入环境[8]。抗生素残留会诱导土壤中ARGs的产生和积累[9-11]。饲料中的抗生素会显著改变肠道中的微生物群落结构,导致肠道中含有抗生素抗性基因的微生物显著增加[13]。进一步的研究表明,动物肠道内的抗性细菌随着粪便排出体外后,其携带的抗生素抗性遗传信息可以借助于整合子、转座子、质粒等可移动遗传单元(Mobile genetic elements,MGEs),通过水平基因转移(Horizon⁃tal gene transfer,HGT)机制转移到环境微生物中,最终通过直接接触或生物富集给人类健康带来巨大的威胁[5,11,14-19]。
已有研究表明,家养鸡较养殖场鸡的肠道菌群更具多样性,且抗生素抗性基因在养殖场鸡的肠道菌群中分布更广泛[13],说明动物肠道中含有丰富的耐药菌和抗性基因,肠道内的ARGs随粪尿排出体外,造成畜禽粪便中ARGs的检出。现有许多学者在畜禽粪便中检测出大量ARGs,如邹威等[22]对华北地区天津市和河北省不同类型、不同规模的养殖场粪便进行检测,研究结果发现,四环素抗性基因、磺胺类抗性基因、大环内酯抗性基因的检出频率达到100%,其他的抗生素抗性基因检出频率基本都在80%以上。ARGs随畜禽粪便进入土壤环境,同时畜禽粪便中残留的抗生素,也会进一步诱导ARGs产生,造成土壤中ARGs的检出。研究表明,在畜禽养殖场及周边土壤检出大量的抗性基因,李锦等[24]发现在辽宁地区畜禽粪便和周边土壤中,磺胺类和四环素类抗性基因均普遍存在,其中磺胺类抗性基因sulⅠ和sulⅡ在两种介质中检出率均为100%。同时,也有研究表明畜禽粪便作为有机肥农用不仅会增加农田系统中抗性基因的丰度,并通过测序发现粪肥施用也会增加土壤中抗生素抗性基因的多样性[10,14,25-28]。
目前整合子(Integron)在广泛的环境介质中被频繁检出,其中I类整合子在各类环境中最为常见,携带的抗药基因盒目前发现100种以上[19,29],也有研究表明整合子与ISCR(Insertion sequence common re⁃gions)耦合可以进行水平转移[5,30-31]。Dang等[33]研究发现,海河沉积物中Ⅰ类整合子intⅠ与磺胺类基因sulⅠ的相对丰度之间存在显著相关性,并证实sulⅠ在环境中的水平传播与intⅠ的介导有关。赵祥等[35]通过对山东4个地区长期使用粪肥的9个典型设施菜地土壤中的ARGs和MGEs进行检测和相关分析,发现可移动元件总量intⅠ与磺胺类抗性基因及大环内酯类抗性基因呈现极显著相关。ARGs可借助MGEs进行水平转移,ARGs不仅影响了土壤环境,还会对农产品甚至人类健康造成危害。
白洋淀区域畜禽养殖业十分发达,按照原国家环境保护总局发布的[2004]43号文件提供的系数测算,鸭粪年排放量达21.33万t[37]。我国每年生产的氟喹诺酮类抗生素(Fluoroquinolones,FQs)有一半是用于畜禽养殖。Zhao等[39]检测发现畜禽粪便中环丙沙星、恩诺沙星的含量每千克达到几十微克。Li等[41]对白洋淀区域河流及底泥中的氟喹诺酮类等22种抗生素进行检测,发现底泥中氟喹诺酮类抗生素含量达到65.5~1166 μg·kg-1。
针对以上情况,本研究选择环丙沙星作为研究对象,借助室内培养试验,采用基因扩增技术检测培养81 d土壤中的ARGs和MGEs的浓度,并进一步对ARGs、MGEs与土壤理化性质之间的相关性进行分析,探究粪源环丙沙星对潮土中ARGs的影响,以期为科学评价氟喹诺酮类抗生素的环境风险和粪肥安全合理施用提供理论依据。
1 材料与方法
1.1 供试土壤
供试土壤为潮土,在土壤系统分类中为干润雏形土(The ustic cambosols)。2012年7月采自河北省某地无污染表层(0~20 cm)土壤,土壤有机质27.1 g·kg-1,碱解氮 103 mg·kg-1,有效磷 3.26 mg·kg-1,pH 值7.99,土壤中无环丙沙星检出。
1.2 土壤处理与试验设计
采集新鲜土样过2 mm(10目)筛,充分混匀后放置于人工气候箱中25℃恒温培养一周,使土壤中微生物活化。然后取土装盆(培养盆规格为顶部直径12.0 cm×底部直径8.5 cm×高10.0 cm),每盆装350 g鲜土,其中含鸭粪的处理则按干土4%比例添加;调节每个培养盆中土壤含水率至田间最大持水量70%左右,置于人工气候箱中25℃非密闭培养,并模拟实际光照周期变化(昼夜各12 h),每日通过称重法补充因蒸发损失的水分,以保持恒定的土壤含水量。
试验共设置5个处理,每个处理3次重复,分别为Ⅰ:CK(对照);Ⅱ:CIP(外源添加CIP);Ⅲ:DF(添加不含CIP的鸭粪);Ⅳ:DF+CIP(在Ⅲ的基础上外源添加CIP);Ⅴ:DF(CIP)(粪源CIP)。其中,含CIP的3个处理中每盆CIP的初始含量保持一致,均为17.15 mg·kg-1。试验所使用的鸭粪从同时饲养的两批鸭子获得,一种是不含环丙沙星的鸭粪;另一种是喂含环丙沙星的饲料所得到的鸭粪。试验选取培养81 d的土样进行检测[43]。
1.3 试验方法
1.3.1 土壤DNA提取
称取约0.500 g供试土样,用FastDNASpin Kit for Soil(MP Biomedicals,USA)试剂盒,按照生产商提供的方法提取土壤总DNA,再用1%的琼脂糖凝胶进行电泳验证。提取的土壤DNA样品用微量核酸蛋白质分析仪(Nanodrop 2000)检测浓度和纯度,所提取的DNA样品的A260/A280值在1.8~2.0之间,表明DNA纯度较高。
1.3.2 ARGs和MGEs的PCR定性检出
本试验选取共6大类抗生素27种ARGs,包括四环素类抗性基因(tetA、tetB、tetC、tetD、tetE、tetG),磺胺类抗性基因(sulⅠ、sulⅡ、sulⅢ),氯喹诺酮类抗性基因(qnrA、qnrS),氨基糖苷类抗性基因[aacC2、aacC4、aadA1、aadA2、aadB、aadD、aac(3)-Ⅰa、aac(3)-Ⅱa、aph(3)-Ⅱa],氯霉素类抗性基因(Cat1、CmlA、Flor)及β-内酰胺类抗性基因(CTX-M、OXA、SHV、TEM);4种MGEs,包括整合子intⅠ、intⅡ,插入序列共同区域(Insertion sequence common regions,ISCRs)orf 513 和ISCR2以及细菌16S rRNA基因进行了PCR检测[15]。具体目标基因及引物序列见表1,引物合成由北京新科开源基因科技有限公司完成。
PCR(Polymerase chain reaction)反应总体系 25 μL,包括:10×PCR Buffer 2.5 μL,dNTP Mix 2 μL,10 μmol·L-1上下游引物各1 μL,r-Tap 酶0.25 μL,模板DNA 2 μL,纯水补足至25 μL。扩增条件为:94 ℃预变性6 min;95℃变性30 s,55℃退火40 s,72℃延伸1 min,共36个循环;72℃延伸7 min。取5 μL扩增后产物,利用1%琼脂糖凝胶电泳进行检测。将目的基因的PCR阳性产物送至美吉生物公司进行测序,测序结果与NCBI数据库进行BLAST同源性比对,证实与已发表相似序列的同源性在95%~100%。
1.3.3 ARGs和MGEs的q-PCR定量检测
采用实时荧光定量PCR技术(Real-time fluores⁃cence quantitative PCR,q-PCR)对土壤样品中的ARGs和MGEs的丰度进行测定。q-PCR选用Thermo Scientific公司生产的Maxima SYBR Green/ROX qPCR Master Mixture(2X),将标准品用纯水按10倍进行5~6个梯度稀释,每个样品设置3个平行,同时设置无模板阴性对照,反应在ABI-7300仪器上进行。反应体系包括:SYBR Mix 12.5 μL,10 μmol·L-1上下游引物各0.5 μL,模板 DNA 1 μL(约 1~10 ng),纯水补足至 25 μL。反应程序为:95℃预变性10 min;95℃变性30 s,55℃退火40 s,72℃延伸1 min,80℃收集荧光信号1 min,共进行40个循环;72℃延伸10 min;PCR扩增完后按照仪器自带程序进行溶解曲线绘制。定量PCR产物用1.0%的琼脂糖凝胶电泳检测特异性。
1.3.4 土壤理化性质的测定
土壤有机质采用水合热重铬酸钾氧化-比色法测定[47],有效磷采用Olsen法测定[47],碱解氮采用碱解扩散法测定(LY/T 1229—1999),pH采用1∶5土水比测定。
1.4 仪器与主要试剂
本试验主要用到的仪器包括:人工气候箱(宁波江南仪器厂:RXZ-430E)、凝胶电泳成像分析系统(BIO-RAD:Universal HoodⅡ)、电泳仪(JUNYI:JY3000+)、PCR 仪(BIO-RAD:T100TM Thermal Cy⁃cler)、生物安全柜(苏洁净化:BHC-1300ⅡA/B2)、快速核酸提取仪(MP:FastPrep-24)、超微量分光光度计(Thermo Fisher Scientific:Nanodrop 2000)、Real Time PCR System(ABI-7300)。主要的试剂包括:环丙沙星(Sigma公司)、Fast DNA Spin Kit for Soil试剂盒(MP 公司)、10×PCR Buffer(Mg+plus)、dNTP Mix⁃ture、r-Taq DNA Polyersss(TaKaRa公司)。
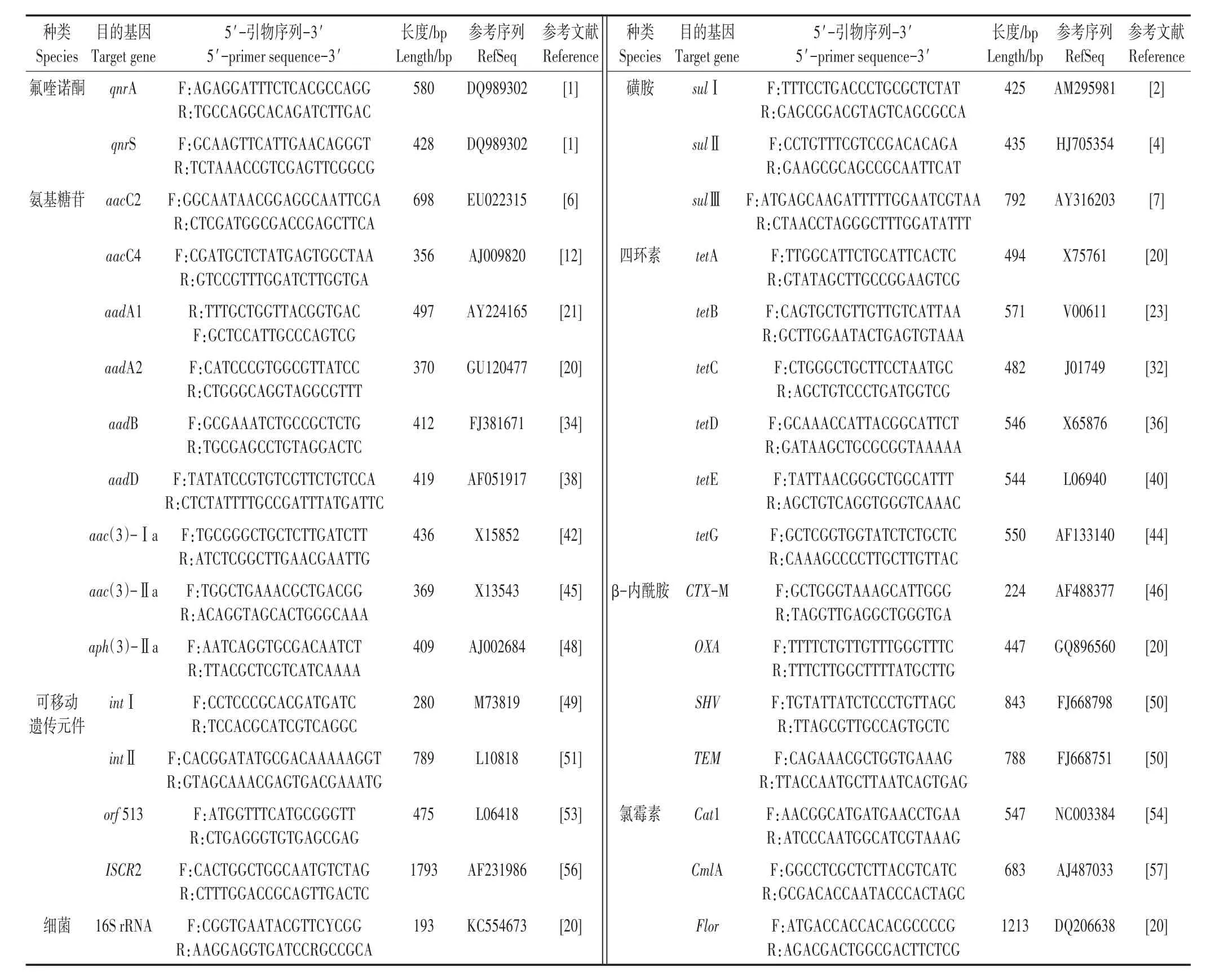
表1 基因种类及引物序列信息Table 1 Gene types and information of primer sequence
1.5 数据分析
抗生素抗性基因的拷贝数的计算:根据目的基因标准品的拷贝数和其扩增的CT值计算标准曲线,代入样品CT值得到土壤样品中各目的基因的拷贝数,并以基因拷贝数/土壤质量(g,干土)为单位进行分析。质粒浓度换算成每微升质粒溶液所携带的绝对模板拷贝数的公式为:
copies·μL-1=[x/(a+b)×660]×10-9×6.02×1023
式中:x为质粒浓度,ng·μL-1;a为载体长度,bp;b为目的基因长度,bp。生成标准曲线的R2值应大于0.98,越接近于1,结果可信度越高;扩增效率E=10-1/斜率-1,E的范围应在0.8~1.2,越接近于1越理想。
采用IBM SPSS Statistics 22对数据进行方差分析,并用软件中的偏相关分析方法进行分析,当P值小于0.05或0.01时,表明在95%或99%的置信区间内具有统计学意义上的显著差异。用Origin 9.0作图。
2 结果与讨论
2.1 不同处理土壤中ARGs和MGEs的检出情况
本研究对不同处理土壤中32种基因进行检测,包括6大类抗生素(四环素、氟喹诺酮、磺胺、氨基糖苷、氯霉素及β-内酰胺类)抗性基因,4种可移动遗传元件(整合子intⅠ、intⅡ,插入序列共同区域orf 513和ISCR2)以及细菌16S rRNA基因。对比分析以上基因在各处理中的检出差异,结果如表2所示。在调查的32种基因中共检出4大类6种抗性基因(tetG、sulⅠ、qn⁃rA、qnrS、aadA2、aadD)和1种可移动遗传元件(intⅠ)。各处理土壤中氟喹诺酮类、磺胺类、四环素类的检出率分别为2/2、1/3和1/6(检出基因数目/检测基因数目);氨基糖苷类抗性基因在CK、CIP及DF处理中检出率为1/9,在DF+CIP、DF(CIP)处理中检出率为2/9。在所检测的4种MGEs中仅检出intⅠ,在各处理中的检出率为1/4。同时各处理中均检出16S rRNA,检出率为1/1。
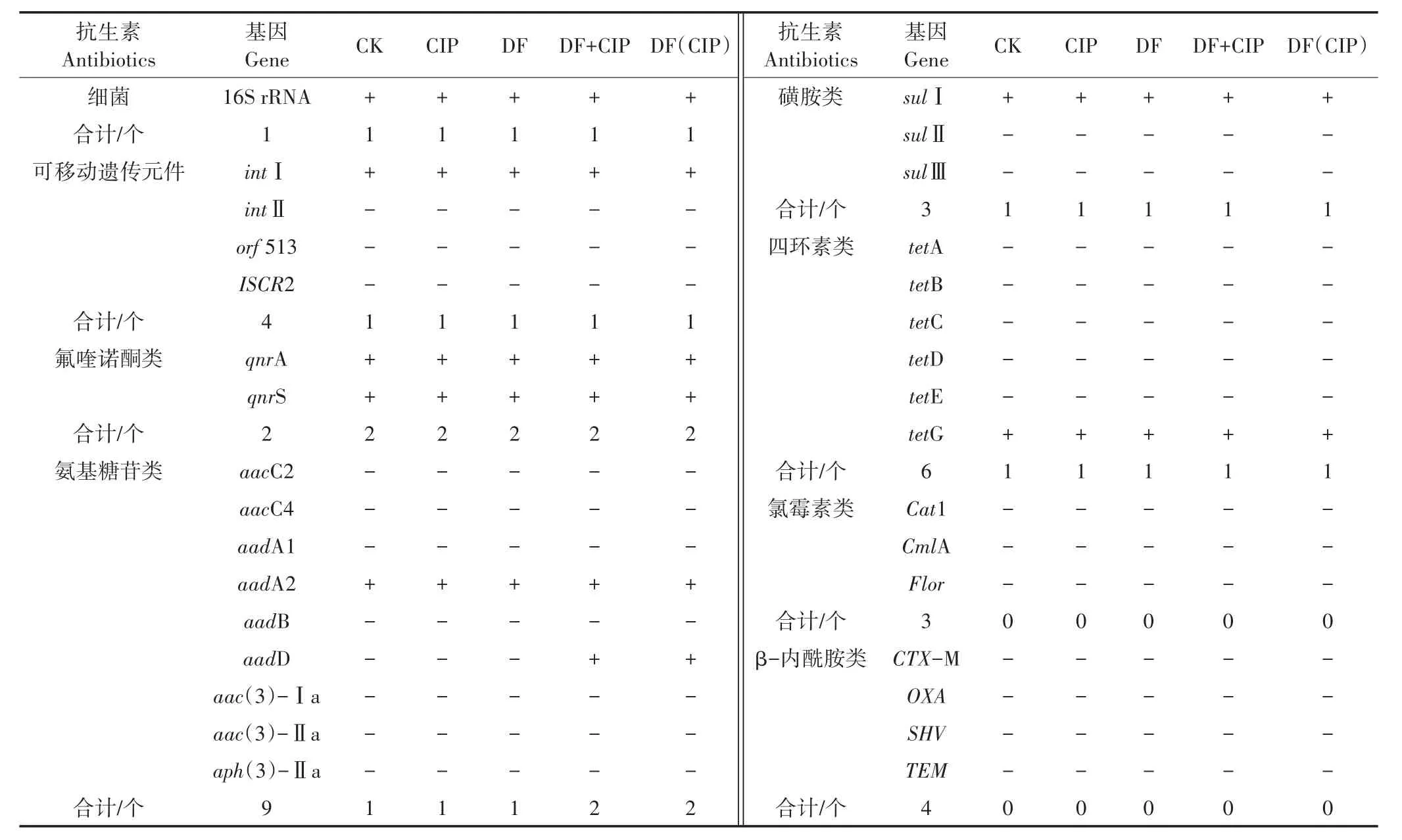
表2 不同处理土壤中ARGs、MGEs及16S rRNA的检出种类及检出率Table 2 Types and detection rates of ARGs,MGEs and 16S rRNA in soil of different treatments
2.2 不同处理土壤中ARGs、intⅠ和细菌的定量分析
为了进一步研究不同处理土壤中ARGs和intⅠ的浓度特征,以及粪源环丙沙星对土壤中ARGs、intⅠ和细菌的影响,本试验利用q-PCR技术,选取各处理中检测率较高的基因(根据表2的PCR检出结果),包括四环素类抗性基因(tetG)、磺胺类抗性基因(sulⅠ)、氨基糖苷类抗性基因(aadA2)、氟喹诺酮类抗性基因(qnrA)、Ⅰ类整合子(intⅠ),对不同土壤样品中的目标基因进行定量分析。各目的基因标准曲线的R2在0.985~0.995之间,扩增效率E在0.88~1.01之间。图1即为不同处理土壤中各目标基因的绝对丰度。
2.2.1 不同处理土壤中ARGs、intⅠ和细菌的浓度特征
培养81 d后,土壤中不同目标基因在不同处理中绝对浓度范围波动较大,在 107~1016copies·g-1之间(图1)。各处理中氟喹诺酮类抗性基因qnrA的检出浓度在1010~1011copies·g-1之间,与CK相比,DF、DF+CIP、DF(CIP)处理中qnrA绝对丰度均显著增加(图1b,P<0.05)。在CK、CIP处理中四环素类抗性基因tetG和磺胺类抗性基因sulⅠ的检出浓度均在109~1010copies·g-1之间(图1c、图1e),氨基糖苷类抗性基因 aadA2的检出浓度在 107~109copies·g-1之间(图1f);在DF、DF+CIP和DF(CIP)处理中tetG检出浓度较高(检出浓度在1012~1014copies·g-1),其次是 sulⅠ(检出浓度在1012~1013copies·g-1)和aadA2(检出浓度在 1011~1012copies·g-1);施加鸭粪的 3个处理与 CK、CIP处理相比,tetG、sulⅠ、aadA2的绝对丰度均增加3~4个数量级,且差异均达到显著水平(P<0.05)。在DF、DF+CIP 和 DF(CIP)处理中intⅠ的检出浓度为1013copies·g-1左右(图1d),16S rRNA检出浓度在1016copies·g-1左右(图1a),与CK、CIP处理中intⅠ的浓度(1012copies·g-1左右)和16S rRNA的浓度(1015copies·g-1左右)相比均增加1个数量级,且差异均达到显著水平(P<0.05)。综合上述分析,添加鸭粪的处理均显著增加了土壤中tetG、sulⅠ、aadA2、intⅠ和16S rRNA的绝对丰度(P<0.05)。已有研究证明畜禽粪便作为有机肥农用会增加土壤中抗性基因的丰度。Schimitt等[27]研究发现施用猪粪后土壤中磺胺类和四环素类抗性基因数量明显增加。Zhu等[52]对饲喂抗生素的猪粪以及施用猪粪的土壤进行检测,发现有63种ARGs相比对照显著增加。黄福义等[14]利用高通量荧光测序对水稻土中295个ARGs进行了定量分析,发现未施猪粪的对照组检出66个抗性基因,施猪粪组检出107个抗性基因,并且约一半ARGs显著富集,表明猪粪施入不仅会增加土壤中ARGs的丰度,也会增加土壤中ARGs的种类,并改变土壤中抗生素抗性基因多样性。

图1 不同处理对土壤中抗生素抗性基因、intⅠ和总细菌基因的影响Figure 1 Effects of different treatments on antibiotic resistance genes,intⅠand 16S rRNA in ustic cambosols
2.2.2 粪源环丙沙星对土壤中细菌、不同类型ARGs和intⅠ的影响
单独施加环丙沙星对土壤中不同种类ARGs的影响不同。CIP与CK处理相比,qnrA绝对丰度显著增加(图1b,P<0.05);tetG的绝对丰度增加4.97×109copies·g-1,sulⅠ、aadA2、intⅠ和 16S rRNA的绝对丰度分别降低1.13×109、1.30×108、3.69×1011copies·g-1和9.54×1013copies·g-1,但差异均未达到显著水平(图1a、图 1c、图1d、图1e、图1f);说明培养81 d后,单独施加环丙沙星会增加土壤中qnrA的绝对丰度,但对tetG、sulⅠ、aadA2、intⅠ和细菌丰度无显著影响。
粪源环丙沙星和等量鸭粪基础上外源添加环丙沙星处理对培养81 d土壤中细菌影响不同。经检测DF(CIP)和DF+CIP处理中环丙沙星的残留量为7.68、4.09 mg·kg-1。DF(CIP)、DF+CIP与DF处理相比,16S rRNA的绝对丰度分别减少1.51×1015、0.99×1015copies·g-1,差异均达到显著水平(P<0.05),说明鸭粪中残留的环丙沙星对细菌存在抑制作用。且与DF处理相比,DF(CIP)处理对细菌的抑制作用强于DF+CIP处理。此结果与周志强等[55]用磷脂脂肪酸方法检测的不同处理对土壤中细菌的变化情况一致。DF+CIP、DF(CIP)与CK相比,土壤中细菌的绝对丰度均显著增加,主要原因为鸭粪对细菌数量的增加高于环丙沙星对细菌数量的抑制。
粪源环丙沙星和等量鸭粪基础上外源添加环丙沙星处理对培养81 d土壤中不同种类ARGs和intⅠ的影响不同。DF(CIP)、DF+CIP与CIP处理相比,qnrA绝对丰度均显著降低(P<0.05),DF(CIP)、DF+CIP处理中鸭粪降低了土壤中的qnrA的丰度;与DF处理相比,DF+CIP处理土壤中qnrA绝对丰度显著增加(P<0.05),DF(CIP)处理中 qnrA 绝对丰度降低0.78×1010copies·g-1,但差异未达到显著水平;即 DF(CIP)处理中残留的环丙沙星对土壤中qnrA无显著影响,DF+CIP处理中残留的环丙沙星会增加土壤中qnrA的丰度(图1b)。DF(CIP)、DF+CIP与CIP处理相比,tetG绝对丰度均显著增加(P<0.05),DF(CIP)、DF+CIP处理中鸭粪增加了土壤中tetG绝对丰度;DF+CIP、DF(CIP)与DF处理相比,tetG绝对丰度均显著降低(P<0.05),即DF(CIP)、DF+CIP处理中残留的环丙沙星降低了土壤中tetG的绝对丰度(图1c)。DF(CIP)、DF+CIP与CIP处理相比,sulⅠ、aadA2绝对丰度均显著增加P<0.05),DF(CIP)、DF+CIP处理中的鸭粪会增加土壤中sulⅠ、aadA2绝对丰度;DF+CIP与DF处理相比,sulⅠ、aadA2绝对丰度分别增加3.08×1012、0.54×1011copies·g-1,但差异未达到显著水平;DF(CIP)与DF处理相比,sulⅠ、aadA2绝对丰度均显著增加(P<0.05),即DF(CIP)处理中残留的环丙沙星会增加土壤中sulⅠ、aadA2绝对丰度(图1e和图1f)。DF+CIP、DF(CIP)与CIP处理相比,intⅠ绝对丰度均显著增加(P<0.05),DF+CIP、DF(CIP)处理中鸭粪会增加土壤中intⅠ绝对丰度;DF+CIP、DF(CIP)与DF处理相比,intⅠ绝对丰度均降低,但差异未达到显著水平,表明CIP+DF、DF(CIP)处理中鸭粪对土壤中intⅠ绝对丰度无显著影响(图1d)。
综合上述DF(CIP)和DF+CIP处理中鸭粪和残留的环丙沙星对ARGs和intⅠ影响的分析,DF(CIP)、DF+CIP与CK处理相比,土壤中qnrA、tetG、sulⅠ、aadA2、intⅠ的绝对丰度均显著增加,说明在本试验中,培养81 d后DF(CIP)和DF+CIP处理均增加了土壤中qnrA、tetG、sulⅠ、aadA2、intⅠ的绝对浓度。与DF处理相比,DF(CIP)、DF+CIP处理均显著降低了土壤中tetG的绝对丰度(P<0.05);DF(CIP)处理显著增加了土壤中sulⅠ和aadA2的绝对丰度(P<0.05);DF+CIP处理显著增加了土壤中qnrA的绝对丰度(P<0.05)。
本试验中DF(CIP)与DF+CIP处理对土壤中同种ARGs的影响存在差异。DF(CIP)与DF+CIP处理相比,qnrA、tetG、16S rRNA的绝对丰度显著降低(图1a、图1b和图1c,P<0.05),sulⅠ、aadA2的绝对丰度显著增加(图1e和图1f,P<0.05)。这两个处理中ARGs丰度不同的原因可能是由于DF(CIP)给鸭子饲喂CIP后,CIP造成鸭子肠道中微生物群落结构和多样性的改变,排出的污染的鸭粪与外源添加CIP的鸭粪中微生物组成和结构多样性存在差异,施用后对土壤中ARGs产生不同的影响,具体还需进一步研究分析。
CK处理中均有不同种类ARGs的检出,其中qnrA、sulⅠ、tetG的浓度较高,检出浓度在109~1010copies·g-1之间。这一方面是由于土壤中微生物的内在抗性,Farias等[58]研究发现在自然环境中,即使没有抗生素存在,ARGs也存在于微生物群落中。楼晨露[10]在不施用猪粪的土壤中也检测到较高浓度的tetA、tetG、sulⅠ和 sulⅡ,相对丰度在 10-4~10-3之间,与 Ji等[16]研究的检出结果一致。另一方面可能是由于外界环境污染,李娟等[59]对北京地区7个典型传统的养猪场废水、周边土壤中氟喹诺酮类耐药基因的污染情况进行监测,从养猪场废水和土壤中均检出了qnrD、qepA、oqxB、qnrS和oqxA等耐药基因。高敏等[60]研究发现,养殖场粪便是空气中ARGs的重要来源,且养殖场舍内浓度远高于舍外。
2.3 不同处理土壤中ARGs、intⅠ丰度与土壤理化性质的偏相关分析
影响抗性基因的产生和转移的因素有很多。理论上抗生素作为抗生素抗性基因的直接选择压力,两者之间存在一定的相关性,但并不是唯一因素。越来越多的研究表明,其他环境因子,如重金属、其他非对应抗生素、pH、有机质、可移动遗传元件等也都对ARGs产生影响。本试验对5个处理中各目的基因绝对拷贝数与16S rRNA绝对拷贝数的比值进行计算,得到目标基因的相对丰度,再利用偏相关分析对土壤样品中各类型ARGs、intⅠ的相对丰度与土壤理化性质的相关关系进行分析,分析结果如表3所示。
本试验中不同种类的ARGs与intⅠ的相关性不同(表3)。在固定其他变量影响的情况下,intⅠ与sulⅠ、aadA2均呈极显著正相关(P<0.01),与qnrA呈显著负相关(P<0.05),与tetG无相关性。Selvaraj等[61]发现在链霉素压力下,有氧生物反应器中intⅠ与氨基糖苷类抗性基因存在显著相关性。赵祥等[35]发现长期使用粪肥的设施菜地土壤中可移动元件总量intⅠ与磺胺类及大环内酯类抗性基因呈现极显著相关。Shi等[62]研究表明在intⅠ的可变盒中,未检测到任何的四环素类抗性基因,进一步探究发现,在土霉素压力下,某些四环素类抗性基因可能是通过转座子进入到质粒,从而进行细菌之间的水平转移。上述研究结果均与本研究结论一致。赵祥等[35]研究表明在长期使用粪肥的9个设施菜地土壤中检测到intⅠ和多种氟喹诺酮类抗性基因(qepA、qnrB、qnrS),分析表明intⅠ与氟喹诺酮类抗性基因无相关性,与本试验中qnrA与intⅠ呈负相关性的结果有区别,原因有待进一步分析。
本试验中不同种类的ARGs与土壤理化性质的相关性不同(表3)。在固定其他变量影响的情况下,qnrA与CIP残留量呈极显著正相关(P<0.01),sulⅠ、aadA2、tetG与CIP残留量均无相关性;TetG与有机质呈极显著正相关(P<0.01),qnrA、sulⅠ、aadA2与有机质均无相关性;土壤中各ARGs与有效磷、pH之间均无相关性。已有研究表明土壤中ARGs与土壤理化性质存在相关关系。王凤花等[63]发现四环素类抗生素不仅与四环素类抗性基因具有相关性,与氨糖苷类、磺胺类抗生素性基因也存在相关性,此结果也与本试验中添加鸭粪和CIP,增加了土壤中sulⅠ、aadA2相对丰度结果类似。同时,王凤花等[63]在含有高浓度四环素抗生素残留的再生水浇灌后的土壤样品中,检测出高浓度的四环素类和磺胺类抗性基因,也检测出intⅠ和转座酶基因。赵祥等[35]发现长期使用粪肥的设施菜地土壤中重金属含量与氟喹诺酮类抗性基因呈极显著相关。楼晨露等[10]研究表明在对长期定量施用猪粪的稻田土壤中四环素类与磺胺类抗生素抗性基因的相对丰度与土壤pH、有机质、重金属As、Cd、Cu、Zn之间存在相关性。
3 结论
(1)本试验中共检出6种抗生素抗性基因(tetG、sulⅠ、qnrA、qnrS、aadA2、aadD)和1种可移动遗传元件(intⅠ),不同处理土壤中检测到的目的基因基本一致。DF(CIP)和DF+CIP处理对土壤中细菌和不同种类ARGs的影响不同。DF(CIP)、DF+CIP处理均显著降低了土壤中16S rRNA、tet G的绝对丰度;DF(CIP)处理显著增加了土壤中sulⅠ、aadA2的绝对丰度;DF+CIP处理显著增加了土壤中qnrA的绝对丰度。DF(CIP)与DF+CIP处理相比,土壤中qnrA、tetG和细菌的绝对丰度显著降低,sulⅠ和aadA2的绝对丰度显著增加,原因有待后续研究。
(2)不同种类的ARGs与intⅠ、土壤理化性质的偏相关性不同。土壤中intⅠ与sulⅠ、aadA2呈正相关,与qnrA呈负相关,与tetG无相关性。QnrA与CIP残留量呈极显著正相关,sulⅠ、aadA2、tetG与CIP残留量均无相关性。TetG与有机质呈极显著正相关,qnrA、sulⅠ、aadA2与有机质均无相关性。土壤中各种ARGs与有效磷、pH之间均无相关性。
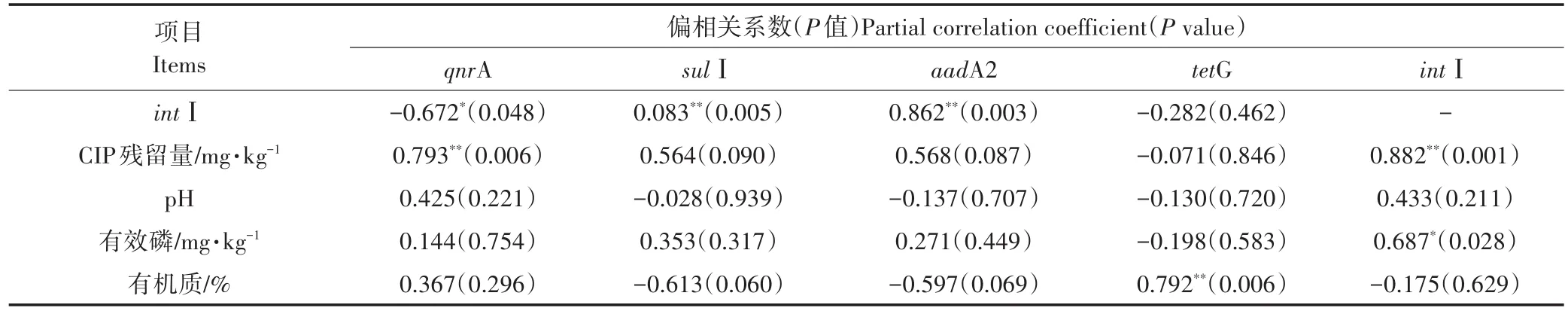
表3 土壤中ARGs、intⅠ及土壤理化性质的偏相关分析Table 3 The partial correlation analysis of ARGs,intⅠand soil physicochemical properties
