基于EST-SSR标记和表型性状的皖北部分牡丹品种遗传多样性分析
2018-12-18谢实龙马同富
陈 杰,蔡 健,李 浩,谢实龙,马同富
(阜阳师范学院 生物与食品工程学院,安徽 阜阳 236037)
牡丹(Paeonia suffruticosa)是原产于我国的芍药属(PaeoniaL.)多年生落叶灌木,具有很高的观赏和利用价值,素有“花中之王”的美誉,在全国各省市区均有栽培,并已被欧美、日本和澳大利亚等多个国家广泛引种种植[1]。根据野生原种和栽培地区的差异,我国的牡丹栽培品种可分为中原牡丹、西北牡丹、西南牡丹、江南牡丹和东北牡丹等5大品种群[2],总计近千个品种,种质资源十分丰富,为牡丹育种和开发利用提供了有利条件。但由于对品种间的遗传背景和亲缘关系缺少深入研究,导致育种工作受到较大限制[3]。
近年来,牡丹品种遗传多样性研究从表型特征、染色体、同工酶等方面更多地转向以分子标记技术为基础的相关研究[4-6]。分子标记是一种较理想的遗传标记形式,基于DNA直接揭示植物物种的遗传差异,具有不受环境影响、标记数量多、多态性高等优点[3]。裴颜龙等[7]采用RAPD(random amplified polymorphic,DNA)技术检测了矮牡丹与紫斑牡丹的遗传多样性,得到的多态条带分别占22.5%和27.6%,种内个体之间分子标记的平均差异分别是7.9和8.7,两个种间分子标记的平均差异是10.3,大于各自的种内差异。侯小改等[8]应用荧光标记AFLP(amplified fragment length polymorphism)技术研究了来自不同地区的30份牡丹品种的遗传差异,结果表明多态性条带百分率达86%,同时还检测到具有一定鉴别价值的品种特异带型。石颜通等[9]利用ISSR(inter simple sequence repeat)技术对不同种源的89个牡丹品种的遗传多样性进行了研究,得到多态位点百分率为94.1%,计算了有效等位基因数、Nei’s基因多样性指数和Shannon信息指数,并通过UPGMA(unweighted pair-group method with arithmetic)聚类分析了亲缘关系,能够有效地区分不同来源的品种。
SSR(simple sequence repeat)又称微卫星,广泛存在于真核生物基因组中,因其具有数量丰富、多态性高等优点,成为植物遗传多样性研究的重要分子标记[10-11]。但早期的基因组SSR标记方法周期长、成本高,不利于推广应用。近年来EST(expressed sequence tag)的迅速发展为SSR标记开发提供了有利条件[12],使得基于EST开发SSR标记更加快速、有效,并应用于牡丹品种遗传多样性等研究[13]。皖北地区是中原牡丹的重要分布区,栽培品种多、种植面积大、来源区系复杂,本研究选择30个品种,根据已有的EST-SSR引物,在分子水平上探讨遗传多样性与亲缘关系,为进一步培育牡丹新品种和产业化开发利用提供参考。
1 材料与方法
1.1 试验材料
本研究所用的30个牡丹栽培品种,均来自安徽省阜阳市颍州西湖风景区。根据研究需要,2014-2015年在颍州西湖风景区牡丹园进行了随机区组试验,重复3次,并在每个试验小区随机定株10株考查9个表型性状:株高、株幅、株型、叶型、叶长、叶宽、花径、花色和花型。品种间的形态差异主要表现在花色、花型和株高等方面(表1)。
1.2 EST-SSR标记分析
选取试验材料的叶片,参照Murray等的方法提取DNA[14]。以侯小改等[15]和张艳丽等[16]设计的EST-SSR引物对牡丹栽培品种进行遗传多样性分析。
根据Panaud等[17]的方法进行PCR扩增。PCR 反应体系:50 ng 的 DNA 模板,1×Buffer,0.2 mmol·L-1dNTPs,1.0 U 的 DNA 聚合酶,0.2 mmol·L-1的上下游引物,1.5 mmol·L-1MgCl2,总计20 μL。反应程序:94℃预变性5 min,然后循环35次(94℃变性45 s、55℃复性45 s、72℃延伸1 min),再72℃延伸5 min。以6%聚丙烯酰胺凝胶电泳PCR扩增产物,0.1%AgNO3染色。
对扩增产物的电泳图谱,用“1”(清晰可辩)和“0”(同一位置缺失)记录带谱,建立品种间的0/1矩阵,按照Nei等[18]的方法计算栽培品种间的相似系数(GS)和遗传距离(GD)。计算公式如下:

式中,Ni、Nj分别是品种i、品种j的特有条带数,Nij是品种i和品种j共有的条带数。
1.3 聚类分析
根据田间调查获取的表型性状数据和ESTSSR分子标记数据,分别以欧氏距离和Nei等[18]的方法计算品种间的遗传距离,用DPS15.10进行系统聚类。在聚类分析前,对花型等描述性状的不同类型先分别赋值1、2、3等,然后再对所有数据进行了标准差标准化转换。通过聚类结果分析供试牡丹品种之间的亲缘关系,并比较基于表型性状和分子标记两类不同数据的聚类结果的异同性,探讨EST-SSR分子标记用于牡丹品种遗传多样性研究的可靠性。
2 结果与分析
2.1 EST-SSR标记的多态性
从30份试验材料中随机选取翠叶紫、白玉兰、雪桂和曹州红等4个栽培品种的DNA作为模板,对文献[15-16]的95对EST-SSR引物组合进行多态性检测,结果显示其中61对引物有多态性,多态率达到64.2%,多态性较高。引物P2、P3、P4、P5、P6、P7和 P9多态性扩增结果见图1所示。筛选出的61对引物在30个样品中扩增出多态性基因型311个,平均每对引物扩增5.1个。
2.2 牡丹品种EST-SSR聚类分析

图1 引物 P2、P3、P4、P5、P6、P7 和 P9 对 4 个牡丹品种的扩增结果 1翠叶紫;2白玉兰;3雪桂;4曹州红

表l 供试牡丹品种
根据DPS15.10中类平均法法(UPGMA)对基于EST-SSR分子标记的品种间遗传距离进行聚类分析。结果表明,若以整个遗传距离的总平均数为尺度,30个牡丹品种大致可分为5个类群(图2)。Ⅰ类包括玉兰飘香、翠叶紫、罂粟红、白玉兰、魏花、金玉交章和大红一品7个品种,花色以白色和紫色系为主,花型主要是单瓣型和皇冠型,株高差异较明显(60.2±13.1 cm);Ⅱ类包括海黄、雪桂、蓝花魁和曹州红4个品种,花色和花型都较多,株高接近(54.9±3.7 cm);Ⅲ类包括鸡爪红、八宝镶、魏紫、茄加紫、重楼点翠、丛中笑和烟笼紫7个品种,花色为红色系和紫色系,花型多皇冠型和菊花型,株高相似(42.0±2.8 cm);Ⅳ类包括胡红、小胡红、艳溢香容、凌花晓翠、墨剪绒、青山贯雪、景玉和盘中取果8个品种,花色多为红色系和紫色系,花色较多,以皇冠型为主,株高差异显著(62.2±21.9 cm);Ⅴ类包括似荷莲、凤丹、大金粉和银红巧对4个品种,花色以红色系和紫色系为主,3种花型,株高差异悬殊(95.2±44.6 cm)。在5个类群中,Ⅰ类和Ⅱ类品种间的亲缘关系最近,Ⅲ类和Ⅳ类次之,而Ⅴ类和其他4类的亲缘关系相对较远。
2.3 牡丹品种表型性状聚类分析

图2 基于EST-SSR标记的30个牡丹品种聚类图
在DPS15.10中以离差平方和法对田间调查的表型性状数据进行聚类分析(欧式距离)。结果显示,若以整个遗传距离的总平均数为尺度,30个牡丹品种大致也可以分为5个类群,见图3。Ⅰ类有10个品种,包括鸡爪红、银红巧对、重楼点翠、八宝镶、魏紫、丛中笑、茄加紫、烟笼紫、胡红和小胡红,花色全部为红色系和紫色系,花型多样化,包括除单瓣型以外的其它5类,株高在各类中最矮(平均41.2 cm);Ⅱ类有4个品种,包括白玉兰、罂粟红、玉兰飘香和翠叶紫,花色为白色和紫色系,花型以单瓣型为主,株高中等(平均70.1 cm);Ⅲ类有8个品种,包括魏花、金玉交章、艳溢香容、大红一品、蓝花魁、海黄、雪桂和曹州红花色多样,花型以皇冠型和菊花型为主,株高较矮(平均49.3 cm);Ⅳ类有7个品种,包括凤丹、大金粉、景玉、盘中取果、青山贯雪、墨剪绒和凌花晓翠花色多为白色和紫色系,花型多样,株高较高(平均82.4 cm);Ⅴ类仅有似荷莲1个品种,植株远高于其他类群(149.7 cm)。5个类群中,Ⅱ类和Ⅲ类品种之间的亲缘关系较近,Ⅴ类(似荷莲)与其他4类的亲缘关系都较远。比较表型性状的聚类结果与EST-SSR标记的聚类结果可看出,两者并不完全一致,如似荷莲、艳溢香容等品种的归类不同。
2.4 EST-SSR标记与表型性状的相关性分析
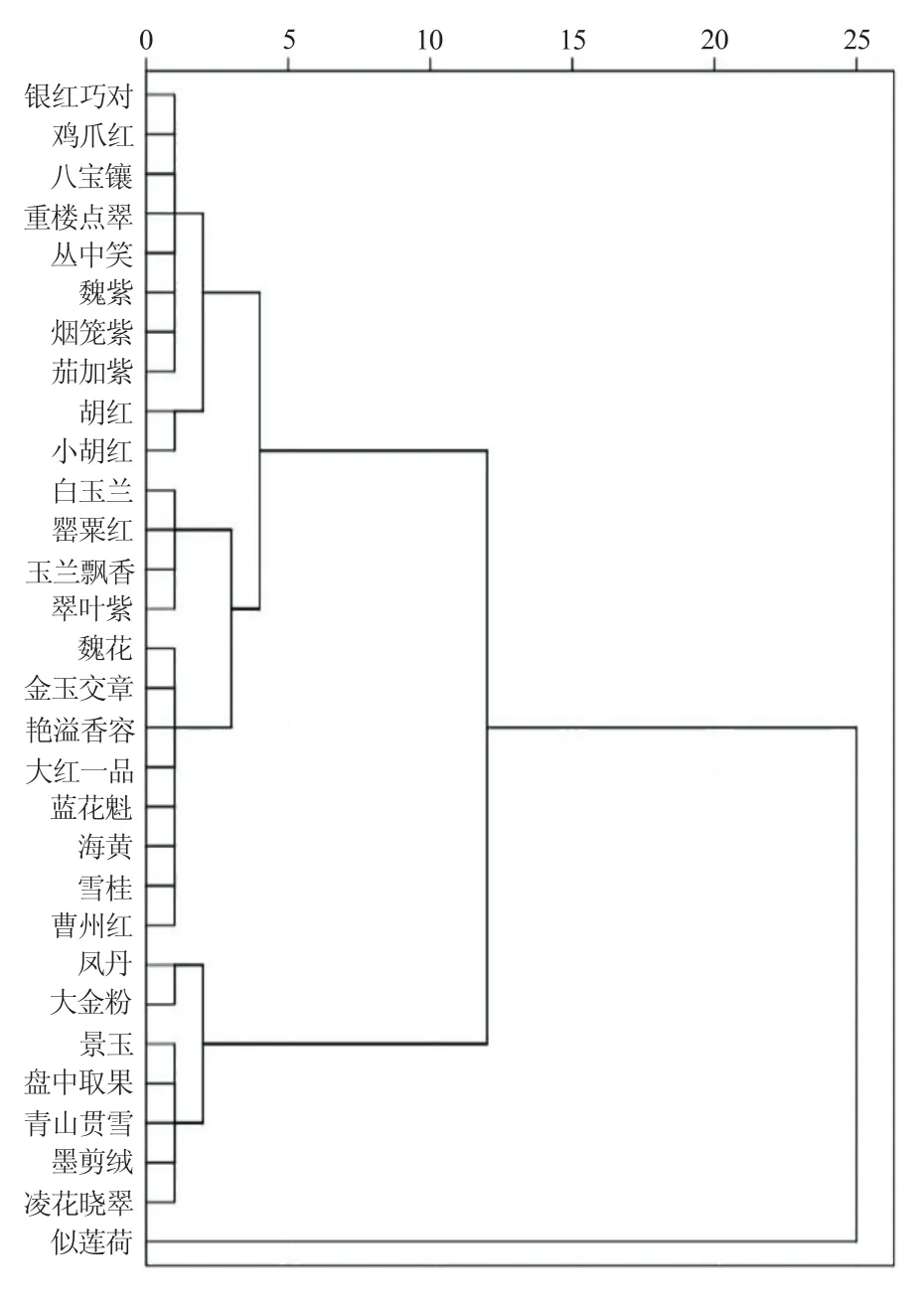
图3 30个牡丹品种基于表型性状分析的系统聚类图
通过Mantel检测方法,对EST-SSR标记与表型性状的遗传距离矩阵进行相关分析,得到相关系数为 0.795(t=10.324>t0.01),因此两个遗传距离矩阵存在极显著的相关关系,也就是说基于ESTSSR分子标记检测的多态性能够反映品种间的遗传差异。
3 讨论
随着现代分子生物学的快速发展,各种分子标记技术不断涌现,如 RAPD、AFLP、ISSR、SRAP(sequence-related amplified polymorphism)、ESTSSR、CDDP(conserved DNA-derived polymorphism)等,极大地提高了科研人员对植物遗传分析研究的能力。牡丹作为一种名贵花卉,其种质资源的开发利用一直受到重视,多种分子标记技术被用于种质亲缘关系的研究[19-20],并且由RAPD、AFLP、ISSR等传统的随机DNA分子标记技术逐步转向SRAP、CDDP、EST-SSR等新型目标分子标记技术。但是,基于不同分子标记的牡丹种质亲缘关系的研究结果并不完全一致,或者其分类结果与基于表型性状的分类结果相矛盾。
李宗艳等[21]利用优化的ISSR-PCR反应体系,筛选了27个引物,对40个不同区域来源的牡丹品种和1个野生种之间的亲缘关系进行了研究,结果显示西南地区牡丹品种栽培起源比较复杂,但本地野生种黄牡丹参与起源的可能性较小。周秀梅等[22]应用SRAP分子标记技术研究了58个中原牡丹核心种质的遗传多样性,结果表明多态性条带64.3%,遗传多样性较丰富,不过聚类结果与牡丹品种的花型、花色等性状没有明显的关系。李莹莹等[23]采用CDDP技术分析了10个花色299份菏泽牡丹品种资源的遗传多样性,检测到的多态性条带和特异条带分别达到95.58%和5.97%,揭示了在品种群水平上的遗传多样性高于花色群体水平以及牡丹花色以粉色系、红色系为中心向其他花色演化的趋势。本研究利用ESTSSR标记和田间试验分析了30份牡丹样本的遗传多样性。从95对EST-SSR引物中筛选出61对多态性引物,多态率64.2%。61对多态性引物在试验材料中扩增出多态性基因型311个,平均每对引物扩增5.1个,并把30个牡丹品种大致分为5个类群。而且基于EST-SSR分子标记的聚类结果与基于表型性状的聚类结果基本一致,两个遗传距离矩阵的相关性也达到极显著水平(r=0.795,t=10.324),说明基于分子标记的聚类结果是可靠的。因此,EST-SSR标记方法能够很好地检测品种间的差异,可应用于牡丹新品种的培育和产业化。
当然,本研究也表现出两种遗传距离的聚类结果也不完全一致,如似荷莲、银红巧对、艳溢香容等品种分别被聚为不同的类群,这种情况的出现可能有多种原因,如:表型性状易容受环境因素的影响,环境效应与环境基因型的互作效应对品种的遗传关系有干扰,这也反映了表型性状聚类的局限性;对表型性状进行统计时出现误差;ESTSSR分子标记方法还不完善,存在局限性。因此,在研究品种间的遗传差异时,需要多种分子标记方法相结合,并增加标记数量,减少外部环境因素的影响,同时要降低表型数量性状数据的统计误差,提高系统聚类的准确性。
4 小结
通过田间试验表型性状和EST-SSR分子标记技术分析了部分牡丹品种的遗传多样性,统计结果表明两个遗传距离矩阵之间存在极显著的相关关系,说明EST-SSR标记方法可以有效地区分牡丹品种间的异同,为培育牡丹新品种和进行产业化开发奠定了技术基础。
