流苏独蒜兰与大花独蒜兰的分类关系
2018-12-14张艺祎吴小倩曹孟霞沈立明翟俊文吴沙沙
张艺祎, 吴小倩, 曹孟霞, 沈立明, 翟俊文, 吴沙沙
(1.福建农林大学园林学院;2.福建农林大学兰科植物保护与利用国家林业与草原局重点实验室,福建 福州 350002)
独蒜兰属PleioneD. Don植物隶属于兰科(Orchidaceae),主要分布在喜马拉雅亚热带山区,中国云南是独蒜兰属植物的多样性分布中心[1-2].独蒜兰属植物共有27种[3-5],已确定的天然杂交种有9个[3,6-7],由于一些种在地理分布、开花时间以及传粉者方面存在重叠,导致物种间的生殖隔离弱,杂交相对容易.目前,英国皇家园艺学会已注册了517个独蒜兰杂交组合[8].尽管独蒜兰属植物被广泛栽培和应用,但属下进化关系一直不清,一些种的分类地位尚存争议,如流苏独蒜兰P.barbaraeBraem.流苏独蒜兰最早是由Barbara Pinkepank于1999年在云南发现,认为是独蒜兰属的新种并命名为P.barbaraeBraem[9].流苏独蒜兰的部分形态特征与大花独蒜兰P.grandiflora(Rolfe) Rolfe相似,花瓣大小相近,唇瓣边缘及褶片均呈撕裂状,顶端微凹;而流苏独蒜兰与大花独蒜兰的背萼片、花瓣、唇瓣形状存在差异,花色也有明显不同.Cribb et al[1]推测流苏独蒜兰可能是大花独蒜兰与独蒜兰或美丽独蒜兰的杂交后代,但由于独蒜兰、美丽独蒜兰与大花独蒜兰均没有重叠分布区,遂将流苏独蒜兰归并为大花独蒜兰.中国植物志英文版(FOC)和世界植物名录(World Checklist of Selected Plant Families)也将P.barbaraeBraem作为大花独蒜兰的异名处理[2-3].此外,曾有学者采用DNA分析认为,流苏独蒜兰是以大花独蒜兰与独蒜兰复合体(该复合体包含独蒜兰、美丽独蒜兰、台湾独蒜兰和四川独蒜兰,因各种之间极易杂交和出现多倍化的现象,难以通过简单的形态学和细胞学的方法进行准确分类,故归为独蒜兰复合体类群)之一为亲本的天然杂交种,但该结果并未正式发表,仅见Paul[10]的报道.由此可见,流苏独蒜兰的分类和界定在学术界尚存在争论,其与大花独蒜兰的分类关系混乱,将其视作大花独蒜兰的一个异名还是大花独蒜兰与其他种的天然杂交种尚有待进一步分子水平的验证.因此,本试验拟通过核基因(nrITS)和叶绿体基因序列(cpDNA)构建独蒜兰属系统发育树,探讨流苏独蒜兰与大花独蒜兰及其疑似亲本的亲缘关系,旨在为独蒜兰属下分类研究提供依据.
1 材料与方法
1.1 材料
供试野生流苏独蒜兰采自云南玉溪(23.36°N,100.42°E,海拔1 360 m),大花独蒜兰采自云南文山(23.23°N,104.45°E,海拔1 480 m).
1.2 形态特征比较
以采集到的流苏独蒜兰和大花独蒜兰的花瓣为主要形态学观察对象,解剖花器官,观察比较花色、萼片、花瓣、唇瓣形状、唇瓣上的斑点颜色及附属物形态,并将流苏独蒜兰与其疑似亲本大花独蒜兰的模式标本进行花部形态特征的比较.
1.3 DNA提取及PCR扩增
参照Zhang et al[11]的方法进行DNA提取、PCR扩增、序列的读取和拼接.PCR扩增使用的引物参照Li et al[12]所用引物,利用1个核糖体基因序列(nrITS)和4个叶绿体基因片段(matK、trnT-trnL、trnL、trnL-trnF)进行引物扩增,用于系统发育分析.
1.4 数据处理
使用DNASTAR软件包中的SeqMan完成DNA序列的编辑和拼接,从GeneBank上获取独蒜兰属15种植物相应的核基因(nrITS)和叶绿体基因片段(matK、trnT-trnL、trnL、trnL-trnF)(表1),选择长叶足柱兰和笋兰作为外类群.
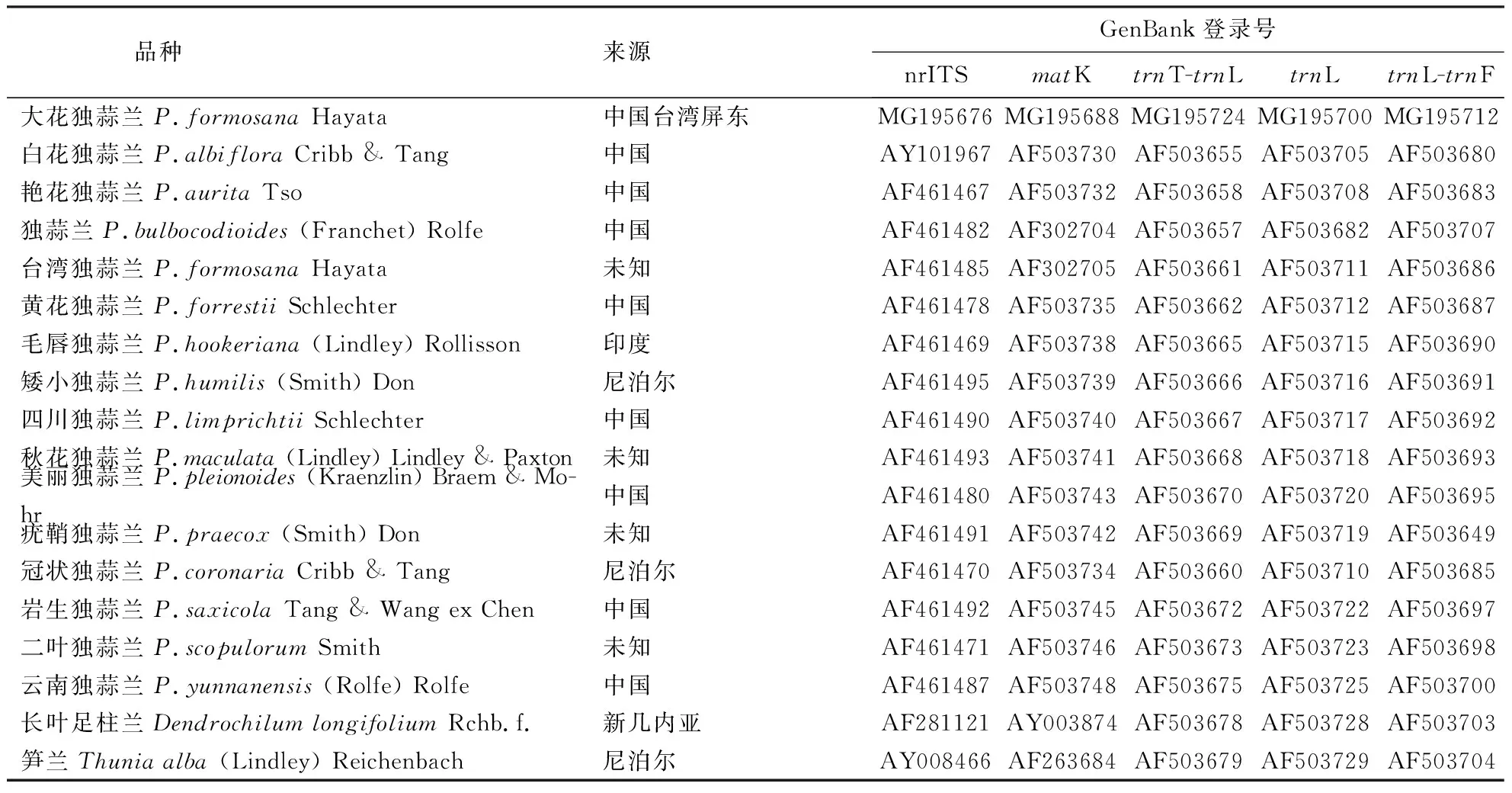
表1 序列数据的GenBank登录号Table 1 GenBank numbers of sequence data
使用最大似然法(ML)、最大简约法(MP)和贝叶斯法(BI)完成系统发育分析.BI和ML分析使用CIPRES Science Gateway web server (MrBayes on XSEDE 3.2.6和RA×ML-HPC2 on XSEDE 8.1.10)软件,参数设置参照Stamatakis et al[13]的方法;MP分析使用PAUP vertion 4软件[14].
2 结果与分析
2.1 流苏独蒜兰与大花独蒜兰的形态比较
流苏独蒜兰(图1A-1C)和大花独蒜兰(图1D-1F)的背萼片均呈镰状倒披针形,侧萼片均倾斜成弓形,呈狭椭圆状,大花独蒜兰的侧萼片比流苏独蒜兰略宽.流苏独蒜兰的花瓣呈狭椭圆状;大花独蒜兰的花瓣呈倒披针形,二者的唇瓣边缘均呈撕裂状,顶端微凹且有4~5条撕裂状附属物.流苏独蒜兰的唇瓣围绕合蕊柱呈囊状并在合蕊柱顶端重叠,大花独蒜兰唇瓣不在合蕊柱顶端重叠.流苏独蒜兰的花为紫红色,花瓣和萼片基部为灰白色,并着有纹脉,唇瓣展开呈倒卵近菱形,喉部白色,分布有深紫色不规则的斑点和条纹;大花独蒜兰的花为白色,唇瓣展开呈宽卵形,白色且分布有黄色斑点和条纹.二者的形态特征有诸多相似,但花部器官形态存在差异且花色差异显著,唇瓣上的斑点及唇瓣形状、颜色也有不同.

A~C:流苏独蒜兰(A:植株;B:花瓣解剖结构;C:唇瓣);D~F:大花独蒜兰(D:植株;E:花瓣解剖结构;F:唇瓣).图1 流苏独蒜兰和大花独蒜兰的花部形态特征Fig.1 Flower morphology of P.barbarae and P.grandiflora
将Braem[9]记载的流苏独蒜兰形态与大花独蒜兰模式标本进行比较(表2),其形态差异与本试验采集的流苏独蒜兰和大花独蒜兰一致.模式标本中的流苏独蒜兰背萼片呈狭椭圆形,大花独蒜兰的背萼片呈倒披针形;二者的侧萼片均呈狭椭圆形且倾斜成弓形;二者的花瓣形状不同,流苏独蒜兰的花瓣呈狭椭圆形,大花独蒜兰的花瓣呈镰状倒披针形,与本试验采集的材料差异一致;流苏独蒜兰的唇瓣上有白喉,且围绕合蕊柱重叠,明显区别于大花独蒜兰;二者唇瓣上的附属物均有5条,且呈撕裂状;二者的花色明显不同,流苏独蒜兰深紫色,大花独蒜兰白色.此外,流苏独蒜兰地生或岩生,大花独蒜兰岩生.
2.2 核基因序列分析
核基因nrITS序列的总长度为671 bp,包含53个信息位点,构建核基因片段的系统发育树.系统发育树结构(图2)表明,ML、MP和BI树中独蒜兰复合体之间的拓扑结构支持率低,且流苏独蒜兰明显与独蒜兰复合体分开,与大花独蒜兰、艳花独蒜兰、黄花独蒜兰聚为一支,与大花独蒜兰的亲缘关系最近,拓扑结构的支持率分别为84、66、52,nrITS序列明确将流苏独蒜兰与大花独蒜兰分开.

表2 流苏独蒜兰与大花独蒜兰模式标本形态特征比较Table 2 Comparison between the holotype of P.barbarae and P.grandiflora
2.3 叶绿体基因序列分析
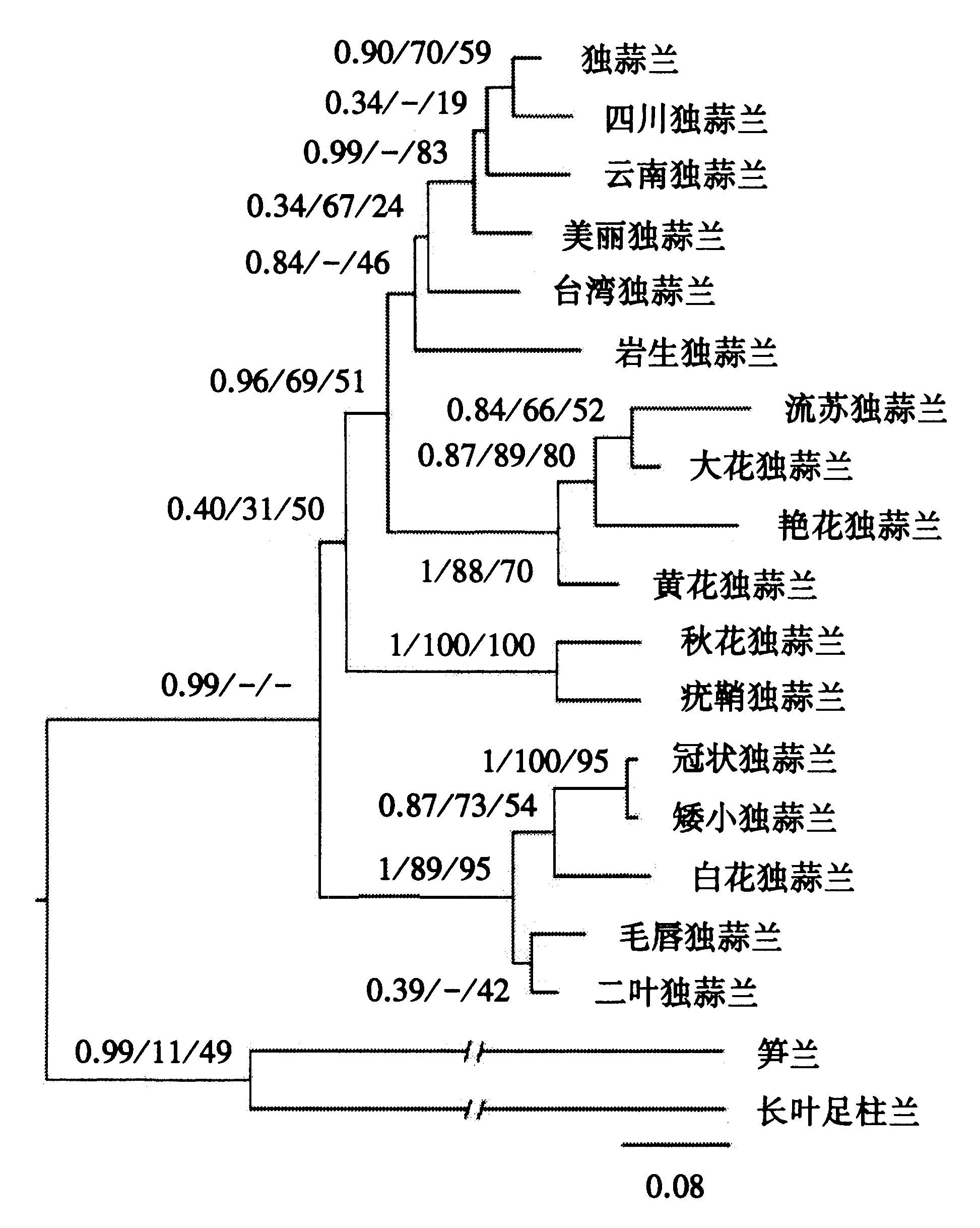
节点前的数字表示BI树的后验概率与ML/MP树的自展值百分比;/-表示ML、MP和BI树的拓扑结构不一致.图2 核基因序列的BI系统发育树Fig.2 Phylogenetic tree inferred from nrITS sequences using Bayesian inference
4个叶绿体基因片段(matK,trnT-trnL,trnL、trnL-trnF)的总长度为2 874 bp,包含56个信息位点,基于此构建系统发育树(图3).图3显示,ML、MP和BI树中独蒜兰复合体之间拓扑结构的支持率不高,与nrITS序列片段构建的系统发育树结果一致,独蒜兰复合体聚为一支,与流苏独蒜兰明显分开,与大花独蒜兰、艳花独蒜兰聚在一起.叶绿体基因序列中,流苏独蒜兰与大花独蒜兰的亲缘关系近且拓扑结构的支持率高,分别为100、94、89.
3 讨论与结论
Braem记载描述的P.barbarae形态特征(表2)为:花大,呈深紫色,花瓣和萼片上无斑点,唇瓣呈倒卵形,喉部白色,唇瓣上分布有形状不规则的微红色斑块,中间有4~5条白色的唇瓣附属物,其中的4条呈明显的撕裂状,中间一条仅在顶上部分呈轻微的撕裂状[9].除唇瓣形状外,本试验采集的流苏独蒜兰与Braem描述的P.barbarae形态特征吻合.经查证,Bream记载的线描图中的唇瓣形状与本试验采集的流苏独蒜兰的唇瓣形状相近,呈倒卵近菱形[9].因此,确定本试验采集的流苏独蒜兰确系P.barbara.
中国植物志中大花独蒜兰的形态特征为:单花,花大呈白色,唇瓣上时有分布深紫略红或棕色的斑点和条纹,还有4~5条不规则的撕裂褶片,唇瓣呈卵形、椭圆形、扇形,3浅裂[2].本试验采集的大花独蒜兰形态特征与之吻合.
流苏独蒜兰与大花独蒜兰的花部形态存在以下不同:流苏独蒜兰的背萼片呈狭椭圆形,大花独蒜兰的背萼片呈倒披针形;流苏独蒜兰的花瓣呈狭椭圆形,大花独蒜兰的花瓣呈镰状倒披针形;流苏独蒜兰的唇瓣呈倒卵状近菱形,唇瓣上有白喉,且围绕合蕊柱重叠呈囊状,大花独蒜兰的唇瓣呈宽卵形,围绕合蕊柱但不重叠;二者的花色明显不同,流苏独蒜兰紫红,大花独蒜兰白色.此外,流苏独蒜兰地生或岩生,而大花独蒜兰岩生,生态习性的不同表明它们对环境的适应性也存在差异.
本试验构建的两种系统发育树中各分支具有相对较高的支持率.nrITS和cpDNA系统发育树的结构均表明,流苏独蒜兰与大花独蒜兰互为姐妹支,同时,流苏独蒜兰没有与其疑似亲本之一的独蒜兰复合体中任何种聚为一支,否定了独蒜兰复合体中任何种是流苏独蒜兰亲本之一的猜想.
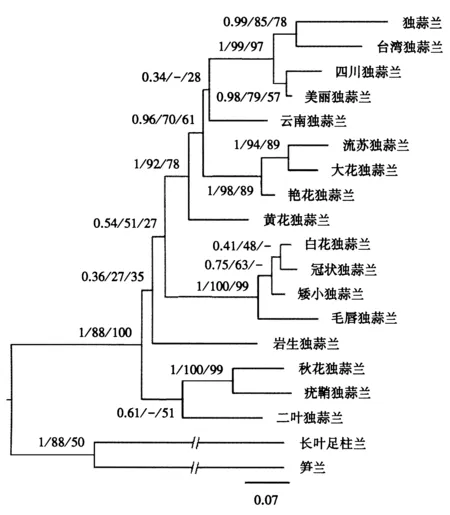
节点前的数字表示BI树的后验概率与ML/MP树的自展值百分比;/-表示ML、MP和BI树的拓扑结构不一致.图3 叶绿体基因序列的BI系统发育树Fig.3 Phylogenetic tree inferred from plastid sequences using Bayesian inference
nrITS系统发育树的支持率相对cpDNA系统发育树低,但nrITS系统发育树中ML、MP和BI树的拓扑结构一致性高,可能因为该属分化时间较短,ITS信息位点较少,且大花独蒜兰和流苏独蒜兰由于分布地存在重叠(模式标本均采自云南),二者可能存在核基因的渐渗,基因之间发生频繁交流[15].因此,流苏独蒜兰和大花独蒜兰表现出形态特征的相似性,如花大,唇瓣上相同的附属物形态及数量,花亭长等;二者同分布于云南地区,同属春花类独蒜兰,开花物候一致,花期4—5月份.对杂交物种而言,形态的相似性被认为是证明其杂交起源有力的证据,但杂交后代可能通过协同进化或回交固定某一亲本的序列,从而导致杂交并不总能在分子水平上被鉴别[16].流苏独蒜兰与大花独蒜兰在分子水平上明显被分开,表明流苏独蒜兰不是天然杂交种.此外,除了早期Braem发现并命名的P.barbarae标本,另有P.moelleriBraem、P.mohriiBraem、P.harberdiiBraem和P.milaniiBraem[9]同被归并入大花独蒜兰[3].研究者将这些不同变异视作流苏独蒜兰.由于本试验取材的局限性,确定本试验采集的深紫色花、唇瓣上有白喉且着有深紫色斑点的流苏独蒜兰与大花独蒜兰有很近的亲缘关系,推测在物种进化历史中流苏独蒜兰与大花独蒜兰可能发生过杂交,导致核基因的双亲遗传与叶绿体母系遗传不一致[17].结合nrITS系统发育树与cpDNA系统发育树拓扑结构的差异,猜测独蒜兰属植物很有可能发生网状进化而导致形态变异的多样性.目前尚无法确定是否存在以大花独蒜兰为亲本的天然杂交种,但明确本试验采集的流苏独蒜兰不是以大花独蒜兰和独蒜兰复合体之一为亲本的天然杂交种.
综上,本试验采集的流苏独蒜兰与大花独蒜兰的花瓣和唇瓣形状存在差异,流苏独蒜兰的唇瓣围绕合蕊柱重叠明显区别于大花独蒜兰,二者的花色存在显著差异;结合nrITS和cpDNA系统发育树,排除了大花独蒜兰和独蒜兰复合体是其亲本的可能;确定流苏独蒜兰与大花独蒜兰的亲缘关系最近,但是否存在以大花独蒜兰为亲本的天然杂交种尚需进一步验证.本试验初步揭示了大花独蒜兰与流苏独蒜兰的复杂关系,明确流苏独蒜兰不是一个天然杂交种,为独蒜兰属下分类研究提供依据.
致谢:诚挚感谢中国科学院昆明植物研究所张伟博士为本试验提供材料,福建农林大学园林学院江鸣涛博士、刘仲健教授在文章写作上给予的建议和指导.
