壳寡糖对异育银鲫生长性能、肠道组织结构和非特异性免疫功能的影响
2018-12-13叶元土蔡春芳吴代武周露阳高敏敏张艳芳
孙 飞 何 杰 叶元土* 蔡春芳 吴 萍 吴代武 周露阳 高敏敏 郁 浓 张艳芳
(1.苏州大学基础医学与生物科学学院,江苏省水产动物营养重点实验室,苏州215123;2.中泰和(北京)科技发展有限公司,北京100027)
壳寡糖(chitosan oligosaccharides,COS)是首先由甲壳素(chitin)脱乙酰化生成分子质量为几十万到几百万几道尔顿、吸收率为1%~5%的壳聚糖(chitosan),再经生物酶解后得到的分子质量<2 000 u、可被吸收进入体内的低聚糖,它由2~10个氨基葡萄糖通过β-1,4糖苷键连接而成,是天然糖中唯一大量存在的碱性氨基多糖。壳寡糖是一种具有水溶性好、生物活性高、易吸收等特点的低分子质量的寡糖类产品[1],与壳聚糖在分子质量大小、水溶性和吸收性等方面有差异[2]。寡糖的结构、乙酰度、纯度以及溶解度的不同,会产生不同的作用效果。壳寡糖以添加剂应用于水产动物饲料中已经有报道。苏鹏等[3]报道,壳寡糖可促进红鳍东方鲀(Takifugurubripes)的生长,提高其非特异性免疫功能;田娟等[4]研究表明,壳寡糖能改善吉富罗非鱼(GIFT,Oreochromisniloticus)的肠道组织结构,调节其肠道主要菌群结构;还有研究表明,壳寡糖可以促进斑节对虾(Penaeusmonodon)的生长,提高机体的抗氧化能力和对氧化胁迫的抗性[5]。

本试验以异育银鲫为试验对象,在基础饲料中添加不同剂量的壳寡糖,来探讨其对异育银鲫生长性能、肠道结构、非特异性免疫指标的影响,同时研究壳寡糖是否对由饲料中氧化油脂引起的副作用有缓解作用。
1 材料与方法
1.1 试验材料
试验所用壳寡糖由中泰和(北京)科技发展有限公司公司提供,是以水产动物甲壳多糖为原料,采用酶法降解壳聚糖与膜分离耦合技术生产的一种吸收性寡糖,壳寡糖纯度为10%,载体为麦芽糊精,脱乙酰度大于90%(2位氨基寡糖),pH为7.0~9.0,壳寡糖分子质量<2 000 u,溶于水。
1.2 试验饲料
配制基础饲料所用原料由江苏省大丰市华辰水产实业有限公司提供。试验用豆油为中粮公司生产的“福临门”牌一级大豆油。氧化豆油制作过程:在正常豆油中添加七水合硫酸亚铁30 mg/L、五水硫酸铜15 mg/L、30%的过氧化氢600 mg/L和0.3%的水,混合后,放在(80±2) ℃的水浴锅中,每30 min充氧气1 min(循环),制作过程共14 d。
首先配制油脂原料分别为正常豆油和氧化豆油的2种基础饲料,然后在含有正常豆油和氧化豆油的基础饲料中分别添加0、0.02%、0.04%和0.06%的壳寡糖,共配制8种试验饲料(CG、OO、CG-200、CG-400、CG-600、OO-200、OO-400、OO-600),以油脂原料为正常豆油的未添加壳寡糖的饲料(CG)作为正对照组,以油脂原料为氧化豆油的未添加壳寡糖的饲料(OO)为负对照组。试验饲料组成及营养水平见表1。各组饲料水分、粗蛋白质以及粗脂肪含量无显著差异(P>0.05)。
饲料原料经粉碎过60目筛,先将大料(比例>4%)置于混合机混匀10 min;再将混合小料与上述混合好的大料逐级稀释混匀,置于混合机混匀10 min;最后将豆油(或氧化豆油)和添加的水(制粒需要)按逐级稀释混匀的办法与上述混合物混合(混合后过40目筛,颗粒物用粉碎机粉碎),再置于混合机混匀20 min。混合好的饲料用小型环膜制粒机(温度65 ℃)制成直径1.5 mm,长2~3 mm的颗粒状饲料,晾至水分在13%左右后,置于-20 ℃冰箱保存备用,使用前按需要量取出饲料自然解冻后投喂。

表1 试验饲料组成及营养水平(风干基础)
1)预混料为每千克饲料提供The premix provided the following per kg of diets:Cu (as copper sulfate) 5 mg,Fe (as ferrous sulfate) 180 mg,Mn (as manganese sulfate) 35 mg,Zn (as zinc sulfate) 120 mg,I (as potassium iodide) 0.65 mg,Se (as sodium selenite) 0.5 mg,Se (as cobalt selenite) 0.07 mg,Mg (as magnesium selenite) 300 mg,K (as potassium selenite) 80 mg,VA 10 mg,VB18 mg,VB28 mg,VB620 mg,VB120.1 mg,VC 250 mg,VD34 mg,VK36 mg,泛酸钙 calcium pantothenate 20 mg,烟酸 nicotinic acid 25 mg,叶酸 folic acid 5 mg,肌醇 inositol 100 mg。
2)营养水平均为实测值。Nutrient levels were all measured values.
1.3 试验鱼与养殖管理
养殖试验在江苏省大丰市华辰水产实业有限公司华垦池塘网箱中进行。在面积为40 m×60 m的池塘中设置24个试验网箱(规格为1.5 m×1.5 m×2.0 m)。为了保证池塘溶氧量均匀,池塘中间设置2台叶轮式增氧机,同时设置1台微孔增氧鼓风机,每2个网箱之间放置1个纳米曝气管(直径20 mm)制成的圆形微孔增氧盘,增氧盘直径为0.5 m,安装在水下1.8 m的深度,投喂期间关闭增氧设备,投喂前使用微孔增氧1 h,其余时间一直使用叶轮式增氧。
试验用异育银鲫鱼种购自江苏省大丰市华辰水产实业有限公司,运输前停食24 h。选取规格整齐的异育银鲫960尾,平均体重为(7.60±0.05) g。将上述试验鱼消毒后,随机分成8组,每组设3个重复(网箱),共计24个网箱,每个网箱40尾鱼。将试验鱼分配到试验网箱中暂养,用相应对照组基础饲料每天早、晚各过量投喂2次,以基础饲料驯化适应2周后开始正式投喂。日投喂2次(06:00—8:30、17:00—19:30),日投喂量为鱼种体重的3%~5%,每10 d估算1次鱼体增重,调整投喂量,正式投喂72 d。每天06:00、18:00测试、记录水温。每5 d测定1次水下30 cm处水质。整个试验期间水温22~36 ℃,溶解氧浓度>5.0 mg/L,pH 8.2~8.6,氨氮浓度<0.2 mg/L,亚硝酸盐浓度<0.01 mg/L,硫化物浓度<0.05 mg/L。
1.4 样品采集
正式养殖试验结束后,停食24 h,进行采样工作。
1.4.1 全鱼采集
正式养殖试验开始前,随机抽取10尾异育银鲫作为初始样本,进行全鱼常规营养成分测定。养殖试验结束后,禁食24 h,对每个网箱的鱼称重,统计数量,计算成活率、增重率、特定生长率和饲料系数,并在每个网箱中随机抽取2尾鱼为全鱼留样,进行常规营养成分分析。
1.4.2 血清采集
用1 mL无菌注射器尾柄静脉采血,置于2 mL Eppenddorf管中自然凝固30 min后,离心(4 ℃,3 500 r/min)15 min后,每管血液取上层血清200 μL,混匀,分装(每管200 μL)于0.5 mL Eppenddorf管中(每个网箱至少分装12管血清),液氮速冻之后置于-80 ℃超低温冰箱保存,用于分析血清非特异性免疫指标。
1.4.3 组织切片样品采集
每个网箱随机选取2尾鱼,取长度为1 cm左右的中肠,于灭菌的0.75%生理盐水中洗净后,置于10%甲醛中固定,用于制作组织切片。各组试验鱼取样的肠段位置保持一致。
1.5 样品分析
1.5.1 常规指标分析
样品用冷冻干燥机干燥至恒重测其水分,之后用于其他指标测定;粗蛋白质、粗脂肪含量以及酸价、过氧化值和丙二醛含量(饲料和油脂)均采用国标方法测定。
1.5.2 血清非特异性免疫指标分析
血清超氧化物歧化酶、过氧化氢酶活性以及丙二醛含量均采用南京建成生物工程研究所生产的试剂盒测定,测定步骤按照试剂盒说明书操作。
1.5.3 肠道组织切片
经过甲醛固定的肠道组织经洗涤、酒精梯度脱水、透明、透蜡、石蜡包埋后切片,切片厚5 μm,苏木精-伊红(HE)染色,光学显微镜下观察肠道组织结构,并采用NikonCOOL-PIX4500型相机进行拍照,Smart-4500软件进行数据量化处理,测量肠皱襞高度、肠皱襞宽度以及肠壁厚度。
1.6 数据处理与统计分析
试验数据以平均值±标准差表示。采用SPSS 22.0软件对试验数据进行处理和统计学分析,组间若有显著差异,则进行Duncan氏法多重比较,显著性水平为P<0.05。
2 结 果
2.1 壳寡糖对异育银鲫生长性能的影响
经72 d的池塘网箱养殖,得到各组异育银鲫的生长性能数据,具体见表2。各组异育鲫鱼在试验期间均无死亡,成活率均为100.00%。与CG组相比,CG-200组的特定生长率升高了16.20%,差异显著(P<0.05);CG-400、CG-600组的特定生长率未发生显著变化(P>0.05);OO组的特定生长率降低了8.10%,差异显著(P<0.05)。与OO组相比,OO-200、OO-400、OO-600组的特定生长率分别升高了5.36%、5.75%、4.21%,但差异均不显著(P>0.05)。此外,OO-200、OO-400、OO-600组的特定生长率与CG组相比无显著差异(P>0.05)。CG-200组的增重率显著高于其他各组(P<0.05),其他各组间差异不显著(P>0.05)。
对于饲料系数,CG-200组与CG组相比降低了6.33%,但差异不显著(P>0.05);CG-400、CG-600、OO组较CG组分别升高了28.58%、80.38%、17.72%,差异显著(P<0.05);OO-200、OO-400、OO-600组较OO组分别降低了6.45%、2.15%、9.14%,但差异均不显著(P>0.05)。
2.2 壳寡糖对异育银鲫体成分的影响
由表3可知,壳寡糖添加量以及豆油是否氧化对全鱼水分、粗蛋白质以及粗脂肪含量均无显著影响(P>0.05)。
2.3 壳寡糖对异育银鲫肠道组织结构的影响
将各组试验鱼中肠同一位置的肠段做成组织切片,并通过显微镜进行观察,结果见图1。与CG组相比,OO组肠皱襞高度减小,宽度增大,肠壁变薄;CG-200组肠道绒毛排列整齐,肠皱襞高度高,宽度小;OO-400组较OO组肠皱襞高度变高,宽度减小,肠壁厚度增厚,达到CG组水平。
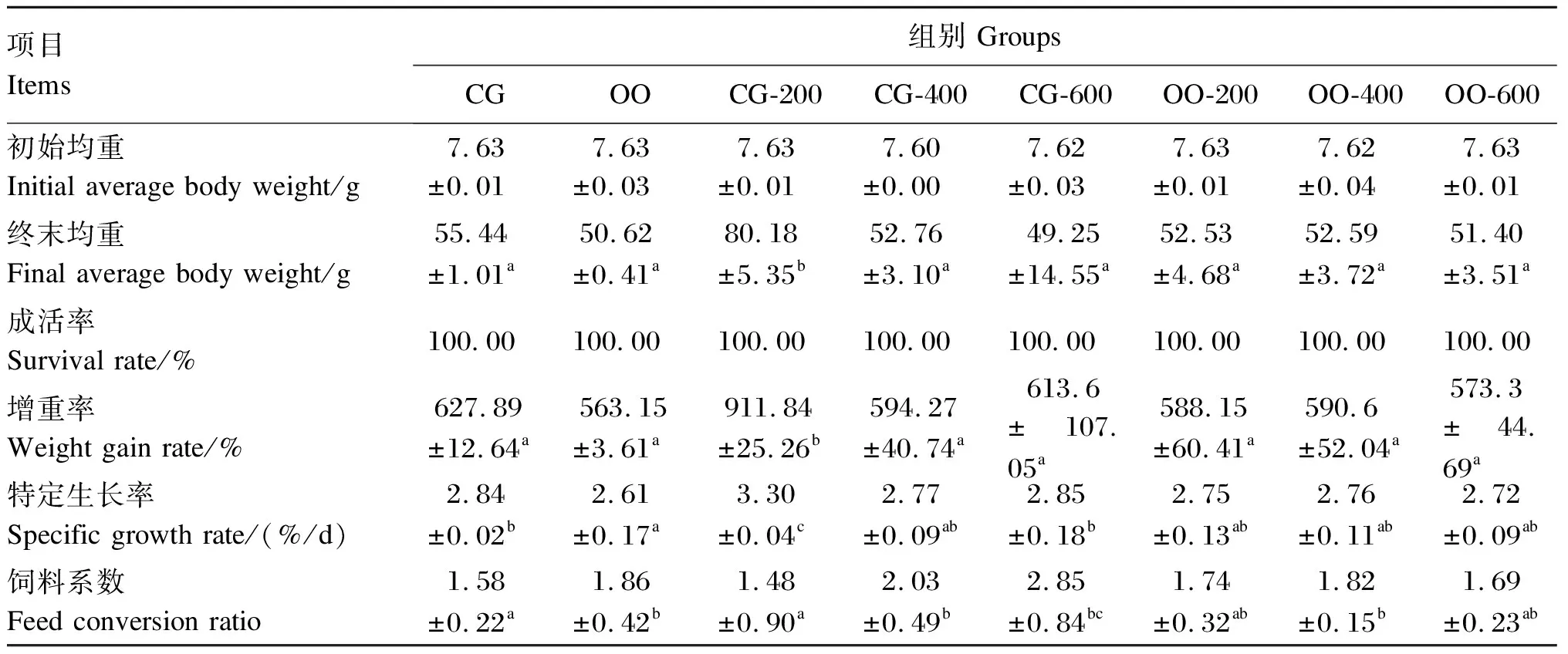
表2 壳寡糖对异育银鲫生长性能的影响
增重率=100×(Wt-W0)/W0;特定生长率=100×(lnWt-lnW0)/t;饲料系数=Wf/(Wt-W0)。式中:Wt、W0分别表示终末均重、初始均重;Wf表示投喂饲料的总量。
同行数据肩标不同字母表示差异显著(P<0.05)。下表同。
Weight gain rate=100×(Wt-W0)/W0; specific growth rate=100×(lnWt-lnW0)/t; feed conversion ratio=Wf/(Wt-W0). In the formula,WtandW0represented the final average body weight and the initial average body weight, respectively, andWfrepresented the total weight of the diet.
Values in the same row with different letter superscripts indicated significantly different (P<0.05). The same as below.

表3 壳寡糖对异育银鲫体成分的影响
对显微镜所采集的图片进行测量,经过Smart-4500软件进行数据量化处理,得到各组异育银鲫肠皱襞高度、肠皱襞宽度、肠壁厚度的数据化结果,分别见图2、图3和图4。
由图2可知,与CG组相比,CG-200组的肠皱襞高度增高了86.84%,差异显著(P<0.05);CG-400、CG-600、OO组的肠皱襞高度均降低,其中CG-600、OO组分别降低了15.08%、17.81%,差异显著(P<0.05)。与OO组相比,OO-200、OO-400、OO-600组的肠皱襞高度分别增高了9.98%、18.89%、4.48%,其中OO-400组与OO组的差异达到显著水平(P<0.05)。此外,OO-200、OO-400组的肠皱襞高度与CG组相比无显著差异(P>0.05)。由图3可知,对于肠皱襞宽度,与CG组相比,CG-200、CG-400组均减小,但差异均不显著(P>0.05),但CG-600、OO组分别增大了47.38%、70.37%,差异显著(P<0.05);与OO组相比,OO-200、OO-400、OO-600组分别降低了36.65%、42.99%、33.72%,差异显著(P<0.05);此外,OO-200、OO-400、OO-600组与CG组相比无显著差异(P>0.05)。由图4可知,各组肠壁厚度的变化趋势与肠皱襞高度的变化趋势基本一致。
1~8分别表示CG、OO、CG-200、CG-400、CG-600、OO-200、OO-400和OO-600组异育银鲫的肠道组织切片。T代表肠壁厚度,H代表肠皱襞高度,W代表肠皱襞宽度。
1 to 8 represented the intestinal sections of crucian carp (Carassiusauratusgibelio) of CG, OO, CG-200, CG-400, CG-600, OO-200, OO-400 and OO-600 groups, respectively. T represented the thickness of intestinal wall, H represented the height of intestinal fold, and W represented the width of intestinal fold.
图1异育银鲫肠道组织切片
Fig.1 Intestinal tissue sections of crucian carp (Carassiusauratusgibelio)

数据柱标注不同字母表示差异显著(P<0.05)。下图同。
Date columns with different letters indicated significantly different (P<0.05). The same as below.
图2壳寡糖对异育银鲫肠皱襞高度的影响
Fig.2 Effects of COS on intestinal fold height of crucian carp (Carassiusauratusgibelio) (n=6)
2.4 壳寡糖对异育银鲫血清非特异性免疫指标的影响
由图5可知,与CG组相比,CG-200、CG-400、CG-600组血清超氧化物歧化酶活性均有所上升,但差异均未达显著水平(P>0.05),其中CG-200组上升最高,上升了6.59%;OO组则下降了27.35%,差异显著(P<0.05)。与OO组相比,OO-200、OO-400、OO-600组血清超氧化物歧化酶活性分别上升了26.25%、40.90%、18.15%,差异显著(P<0.05),其中OO-400组与CG组之间无显著差异(P>0.05)。对于血清丙二醛含量,与CG组相比,CG-200、CG-600组下降,CG-400组上升,但差异均不显著(P>0.05),但OO组上升了25.72%,差异显著(P<0.05)。与OO组相比,OO-200、OO-400、OO-600组血清丙二醛含量分别下降了12.98%、16.20%、27.53%,但差异均不显著(P>0.05),且OO-200、OO-400、OO-600组血清丙二醛含量与CG组相比无显著差异(P>0.05)。血清过氧化氢酶活性各组之间均无显著差异(P>0.05)。
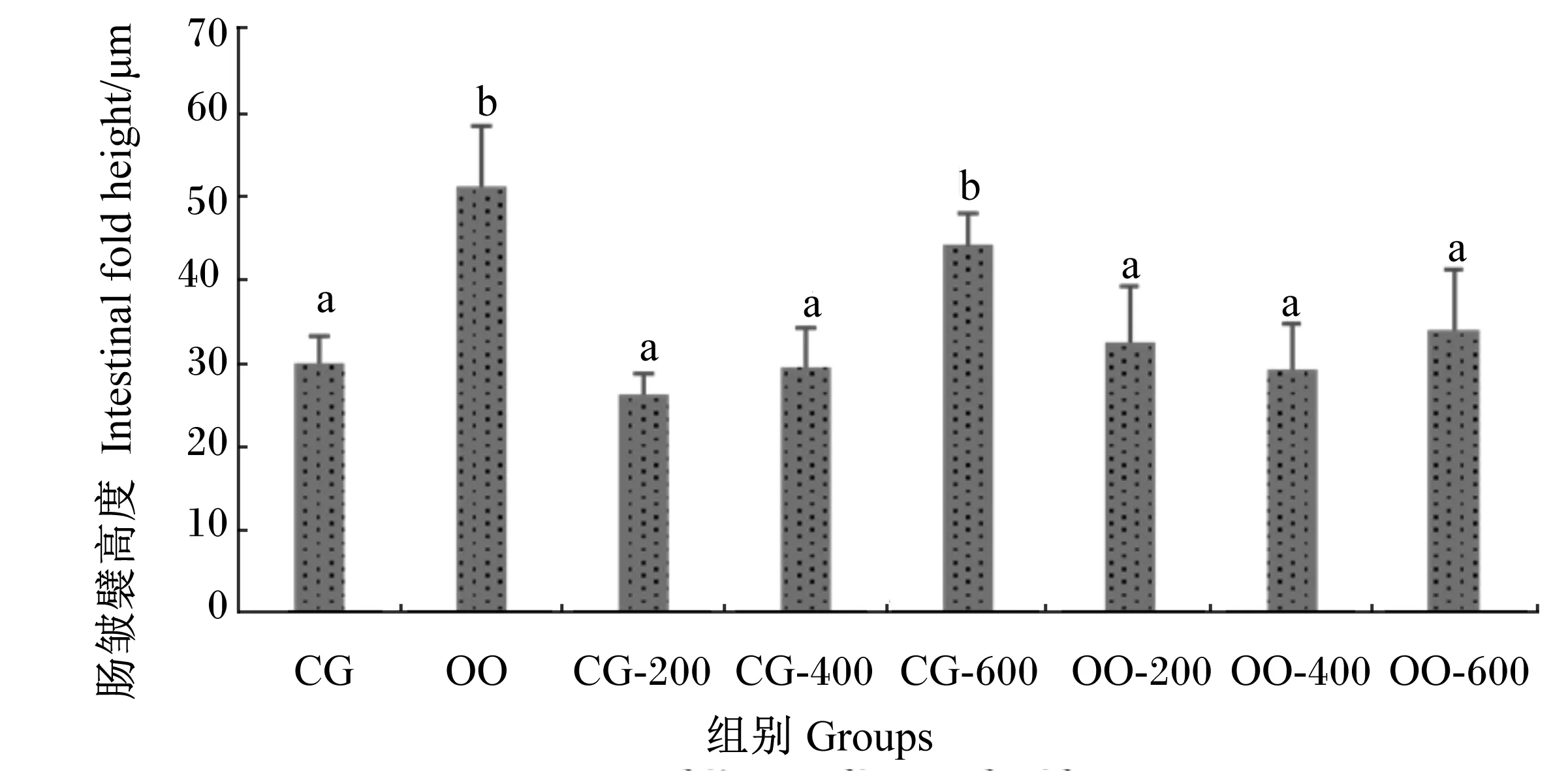
图3 壳寡糖对异育银鲫肠皱襞宽度的影响
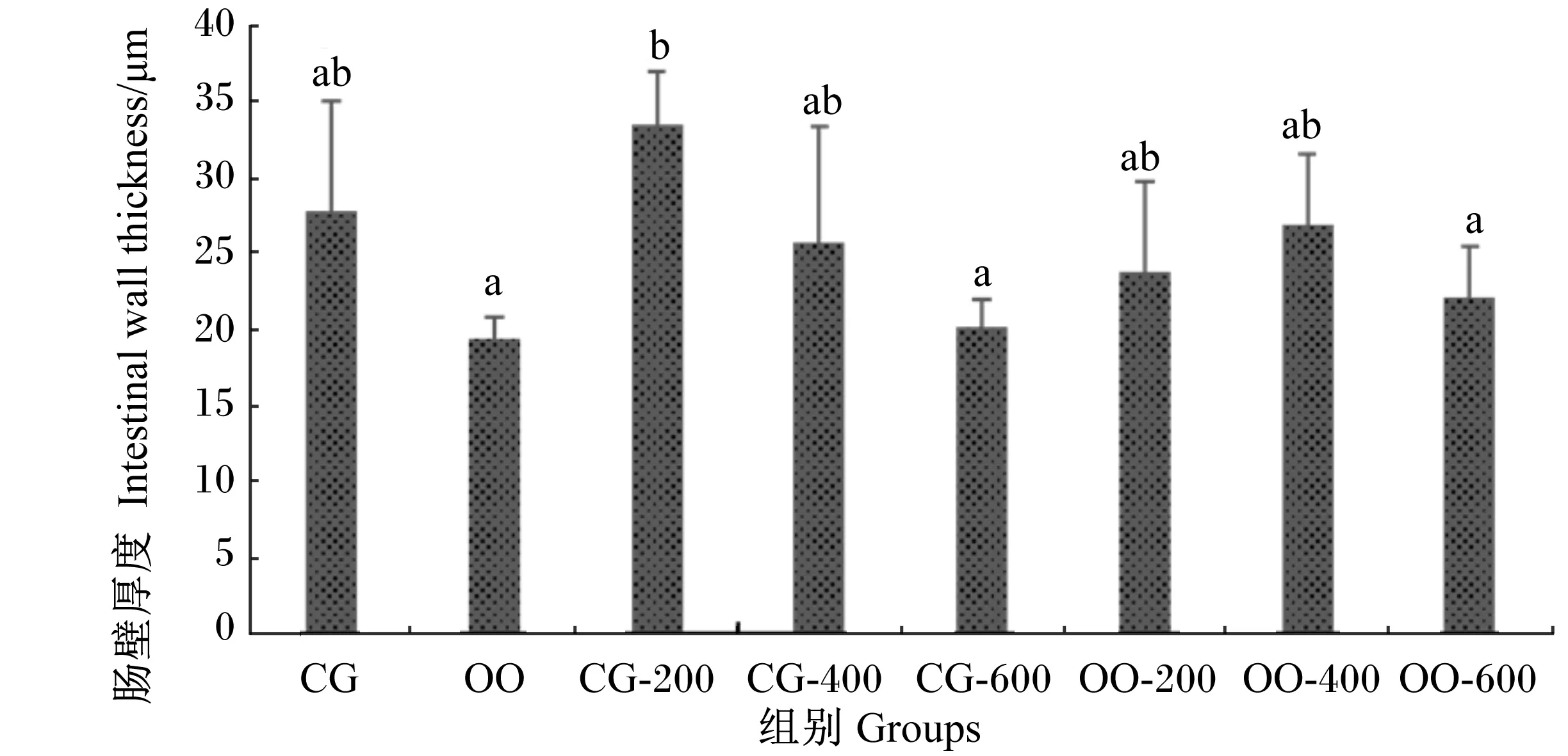
图4 壳寡糖对异育银鲫肠壁厚度的影响

图5 壳寡糖对异育银鲫血清非特异性免疫指标的影响
3 讨 论
本试验条件下,以CG组为对照,在含4%豆油的饲料(常规饲料)中添加0.02%壳寡糖使异育银鲫的特定生长率升高了16.20%,并且饲料系数降低了6.33%;但饲料中添加0.04%、0.06%壳寡糖时异育银鲫的特定生长率未产生显著变化;氧化豆油替代豆油使异育银鲫的特定生长率下降了8.10%,饲料系数增加了17.72%。以OO组为对照,在含4%氧化豆油的饲料中添加0.04%壳寡糖可以使异育银鲫的特定生长率升高了5.75%,饲料系数下降了2.15%,使异育银鲫生长速度和饲料系数恢复到饲喂常规饲料异育银鲫的水平。
壳寡糖的生物活性取决于自身的物理化学性质,如脱乙酰度、电荷分布以及化学修饰[15]。壳寡糖主要通过以下途径对动物生长性能产生影响:1)促进矿物元素的吸收。壳寡糖分子上含有氨基(—NH2)和羟基(—OH)等活性基团,很容易与矿物元素结合后在小肠被吸收[16]。2)改善肠道结构。研究表明,饲料中添加适量壳寡糖后,肠道绒毛变高、变细,同时可以增加绒毛密度,使绒毛与食物接触面积更大,促进肠道对食物的消化与吸收[17]。
对肠道黏膜组织结构的改善是壳寡糖的一个重要的作用位点。鱼类的肠道是消化与吸收营养物质的主要场所,也是鱼类最大的黏膜免疫器官,肠道形态结构的正常是营养物质吸收和肠道免疫正常的基础[18]。本试验发现:在常规饲料中壳寡糖添加量为0.02%时,异育银鲫肠道绒毛排列整齐均匀,与CG组相比,其肠皱襞高度增加了86.84%,肠壁厚度增加了20.45%,肠皱襞宽度减少了12.18%,肠道结构改善效果最好;氧化豆油使异育银鲫肠道结构遭到损害,肠皱襞高度减少了17.81%,肠皱襞宽度增大了70.37%,肠壁厚度减少了30.33%;在氧化豆油饲料中添加0.04%壳寡糖时,与OO组相比,其肠皱襞高度增加了18.89%,肠皱襞宽度减少了2.98%,肠壁厚度增加了38.35%,有效地改善了饲料中氧化油脂带来的负作用,使鱼体肠道结构达到饲喂常规饲料水平。这表明,饲料中添加适量壳寡糖可以有效地改善肠道结构,增大绒毛与食物的接触面积,促进对营养物质的吸收。Dimitroglou等[19]在饲料中添加适量的甘露寡糖使金头鲷(Sparusaurata)肠道皱襞高度、密度都有一定增加;Pryor等[20]在饲料中添加适量甘露寡糖同样使墨西哥湾鲟的肠道皱襞高度、密度增加,这与本研究结果相似。
壳寡糖对肠道结构的改善主要可能是在于壳寡糖能抑制病原菌在肠道黏膜上的定植,改善肠道菌群结构,促进肠道上皮细胞增殖,有利于动物消化道形态结构正常发育,其作用途径有2个:1)壳寡糖具有高亲和力的配体,能提供与细菌外源凝集素特异性吻合的结合位点,从而阻断病原菌与肠黏膜上皮细胞结合[21];2)壳寡糖能促进有益菌的增殖,使其能更广泛地附着在肠壁表面,从而减少有害菌与肠壁的接触[22]。也有研究表明,壳寡糖对肠道内环境改善的同时,也有利于鱼体对营养物质的消化吸收。壳寡糖本身可以作为促生长因子被有益菌所利用,并能产生B族维生素,促进肠道蠕动,提高鱼体对营养物质的吸收[23]。本研究还发现,饲料中添加壳寡糖后,异育银鲫肠壁厚度增加,这与刘爱军君[22]的观点不一致,刘爱君等[22]研究发现饲料中添加壳寡糖后奥尼罗非鱼(Oreochromisniloticus×O.aureus)肠壁厚度减少,这可能与养殖动物的种类、地区、时间以及水体等不同有关,需要进一步的研究。
壳寡糖作为一类水溶性的、容易吸收的2-氨基寡糖,可能被鱼体吸收并在抗氧化损伤、维护免疫防御系统结构与功能完整等方面发挥生理作用,提升鱼体的免疫防御能力。鱼类是较低等的脊椎动物,非特异性免疫是鱼类主要的免疫系统。血清中超氧化物歧化酶、过氧化氢酶活性以及丙二醛含量是衡量鱼类非特异性免疫的重要指标。超氧化物歧化酶是生物体内重要的抗氧化酶,其主要功能是清除自由基;过氧化氢酶是机体生物防御体系的关键酶之一,其主要功能是清除体内的过氧化氢,使机体免受过氧化氢的毒害;丙二醛是生物体内脂质过氧化反应的氧化终产物,脂质过氧化会引起细胞损伤,因此血液中丙二醛的含量可以间接地反映机体内细胞的损伤程度。本试验结果表明,在常规饲料中添加0.02%壳寡糖后,以CG组为对照,血清超氧化物歧化酶活性上升了6.59%,丙二醛含量下降了16.94%;氧化豆油使血清超氧化物歧化酶活性下降了27.35%,丙二醛含量上升了25.72%;以OO组为对照,在含氧化豆油的饲料中添加0.04%壳寡糖后,血清超氧化物歧化酶活性上升了40.90%,丙二醛含量下降了16.20%,异育银鲫血清超氧化物歧化酶活性、丙二醛含量达到了饲喂常规饲料异育银鲫的水平。壳寡糖提升鱼类的非特异性免疫能力可能从以下方面来实现:1)壳寡糖具有清除羟自由基(·OH)的能力,从而提升鱼体的抗氧化能力,保护免疫器官,进而增强免疫功能[23]。2)壳寡糖可促进有益菌如双歧杆菌等的大量增殖,而双歧杆菌可以提高机体的抗体水平,激活巨噬细胞的吞噬活性,从而增强机体的免疫功能[24]。
在异育银鲫常规饲料中添加壳寡糖并非添加量越大效果越好,CG-400和CG-600组特定生长率相对于CG-200组显著降低,可能的原因有:1)饲料中添加的壳寡糖在鱼体肠道内不被消化,直接被肠道上皮细胞吸收,进入血液循环,与体内各种基团结合发挥其多种功能。随着壳寡糖添加量的增加,鱼体本身不能通过调节作用减少对壳寡糖的吸收,虽然增加量不多,但可能壳寡糖会在血液循环中与其他基团结合过度,导致内稳态失衡。2)饲料添加适量壳寡糖会增加鱼体的免疫能力,但随着壳寡糖添加量的增加,可能会使鱼体产生免疫抑制,影响鱼体免疫系统。
饲料中的氧化油脂对异育银鲫的生长有负面影响,饲料中补充壳寡糖可以一定程度地修复这类负面影响。饲料中油脂氧化后产生的氧化油脂会对水产动物的生长性能以及肠道健康等造成损害,主要原因有以下几点:1)氧化油脂会导致饲料适口性下降,降低摄食量[25];2)氧化油脂所含有的初级和次级氧化产物,如过氧化物、丙二醛等,会打破机体自由基代谢平衡,使自由基异常增加,破坏抗氧化酶活性,导致鱼类产生氧化应激损伤[26];3)氧化油脂会导致虹鳟(Oncorhynchusmykiss)胃肠无食物,出现积水[27],草鱼(Ctenopharyngodonidella)摄食含氧化油脂的饲料后肠道紧密连接结构打开,肠道通透性变大[28]。壳寡糖丰富的生物活性如抗氧化作用等可能是其促进异育银鲫生长同时缓解由氧化豆油引起的负面影响的主要原因。本试验所用壳寡糖分子质量<2 000 u,脱乙酰度>90%。Feng等[29]比较了不同分子质量的水溶性壳寡糖的抗氧化能力,结果显示分子质量越小其抗氧化能力越强,这可能是受分子间氢键的影响。高分子质量的壳聚糖结构紧凑,分子内氢键强,壳聚糖的抗氧化能力受其影响较大。低分子质量的壳聚糖结构较松散,分子内氢键弱,壳聚糖的抗氧化能力受其影响较小。此外,低分子质量的壳聚糖比高分子质量的壳聚糖拥有更多的自由羟基和氨基,其清除超氧阴离子自由基的能力明显高于高分子质量的壳聚糖。Je等[30]研究了不同脱乙酰度的壳寡糖清除自由基的能力,发现壳寡糖清除自由基的能力强弱取决于壳寡糖的脱乙酰度程度,结果显示脱乙酰程度最大的壳寡糖有最好的清除自由基的能力,这可能是由于壳寡糖能够提供正电子与自由基反应,将自由基转变成了更稳定的产物,从而终止自由基链式反应。以上研究结果表明本试验所用壳寡糖具有较强的抗氧化性,壳寡糖可通过提升异育银鲫的抗氧化能力,在一定程度上改善由氧化豆油带来的鱼体损伤。
4 结 论
① 在含4%豆油的常规饲料中添加0.02%的壳寡糖可以促进异育银鲫生长,并维护鱼体健康。
② 氧化豆油会对异育鲫鱼的生长和健康造成负面影响,在该饲料中添加0.04%的壳寡糖可改善由氧化豆油引起的负面影响,使异育银鲫的健康程度达到饲喂常规饲料异育银鲫的水平。
