壳聚糖酶的研究进展
2018-12-13鲁晶娣韦盘秋张兴猛黄锦祥黎娅伍时华易弋
鲁晶娣,韦盘秋,张兴猛,黄锦祥,黎娅,伍时华,易弋*
(1.广西科技大学 生物与化学工程学院,广西 柳州 545006;2.广西科技大学 广西糖资源绿色加工重点实验室,广西 柳州 545006;3.广西科技大学 广西高校糖资源加工重点实验室,广西 柳州 545006)
1 概述
壳聚糖是一种具有独特功能特性的天然阳离子聚合物,是由自然界中广泛存在的甲壳素经过脱乙酰作用得到的,主要由D-氨基葡萄糖通过β-1,4-糖苷键连接而成。大量研究表明壳聚糖在食品、化工、医药及农业领域都有广泛的应用,但由于壳聚糖的分子量较大,只能溶于某些酸性溶液中,阻碍了壳聚糖的有效利用。壳寡糖又名壳聚寡糖,是壳聚糖经壳聚糖酶降解后的产物,具有分子量低,溶解性高,更容易被生物体吸收等特点,壳聚糖某些特殊功能只有将其降解成壳寡糖才能表现出来。
壳聚糖易受多种酶类的影响,这些酶类主要包括特异性(壳聚糖酶)和非特异性(碳水化物酶、蛋白酶、脂肪酶等)[1]。自1973年,Shimosaka等首先提取出壳聚糖酶以来,科研人员相继从多种微生物(包括细菌、放线菌、真菌及病毒等)中分离、纯化得到壳聚糖酶。2004年,酶命名委员会将壳聚糖酶(EC3.2.1.132,壳聚糖N-乙酰氨基葡萄糖水解酶)定义为能够在部分乙酰化壳聚糖中催化水解β-1,4糖苷键的酶。另外,还有一些非特异性酶如常见的碳水化合物酶、蛋白酶和脂肪酶也显示出了对壳聚糖的水解能力。
2 壳聚糖酶的来源与分布
壳聚糖酶主要是由微生物和植物产生的,该酶在这些微生物和植物的营养和防御功能上起着重要的作用,于1973年在研究土壤中的细菌和真菌时首次发现。在过去30多年时间里,已经有大量有关细菌、真菌以及植物中壳聚糖酶的结构、纯化和理化特性等方面研究的论文被报道[2]。
目前,从很多细菌中已经发现胞外壳聚糖酶,包括Bacillussp.[3-5],Serratiasp.[6,7],Janthinobacteriumsp.[8],Paenibacillussp.,Acinetobactersp.[9],Streptomycessp.[10]等。在这些细菌中,Bacillussp.和Streptomycessp.的研究最为广泛,研究范围包括酶的结构和分子水平。真菌壳聚糖酶的研究很少,已经报道的来源于真菌的壳聚糖酶包括Aspergillussp.,Gongronellasp.以及Trichodermasp.[11-14]。
蓝藻是一类具有光合作用的原核生物,它们在生理、形态和发育特征上存在可变性。近期报道中,研究人员在AnabaenafertilissimaRPAN1菌株中发现抗真菌壳聚糖酶的存在,并且可以起到抑制真菌的作用[15]。
上述壳聚糖酶大多数来源于微生物,然而也有少数来源于植物。在洋葱和韭菜根部发现壳聚糖酶活性物质。Hsu等从冬笋中发现了2种具有热稳定性的壳聚糖酶[16]。
3 壳聚糖酶的定义与分类
壳聚糖酶是对壳聚糖具有专一性水解的酶,而不降解甲壳素。一些特定的酶,例如普通的碳水化合物、蛋白酶和脂肪酶对壳聚糖也有水解能力。根据壳聚糖酶水解底物的不同,将壳聚糖酶分为3类:第I类壳聚糖酶水解(GlcN-GlcN)和(GlcNAc-GlcN)两种糖苷键,第II类壳聚糖酶只能水解(GlcN-GlcN)一种糖苷键,第III类壳聚糖酶能水解(GlcN-GlcN)和(GlcN-GlcNAc)两种糖苷键(见图1)。内切水解GlcN-GlcN糖苷键是壳聚糖酶常见且唯一的特性,而不能催化GlcNAc-GlcNAc糖苷键。几丁质酶与壳聚糖酶差别很小,都可作用于不同程度的脱乙酰化壳聚糖。两种酶的区别在于:壳聚糖酶易降解脱乙酰化的壳聚糖,几丁质酶更易降解乙酰化的壳聚糖。
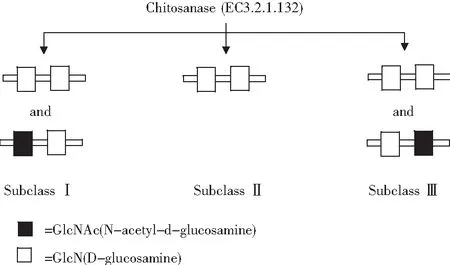
图1 壳聚糖酶活性位点分类方式
根据氨基酸序列不同,可将壳聚糖酶分为5个糖苷水解酶家族,分别为:GH-5、GH-8、GH-46、GH-75和GH-80。在这些家族中,GH-75的成员主要来自真菌和放线菌,GH-46家族来自于Bacillus和Streptomyces的壳聚糖酶,其催化特性、酶反应机理和蛋白结构已被广泛研究。
近年来,科研人员发现一种新型壳聚糖酶,该酶可从壳聚糖/壳寡糖的非还原末端水解GlcN,产生这种外切壳聚糖酶的微生物群已经被报道[17]。2008年,酶命名法委员会定义了一种新的酶类——壳聚糖外切(1,4)-β-D-氨基葡萄糖苷酶),这种酶也被称为外切壳聚糖酶,该酶可以水解GlcN-GlcNAc,但不能水解GlcNAc-GlcNAc。
4 壳聚糖酶发酵生产
通过微生物液态发酵或固态发酵得到胞外壳聚糖酶的产量。迄今为止,已报道的大多数壳聚糖酶都是可诱导生产的。胶体壳聚糖的诱导作用已经被证实,在大多数的壳聚糖发酵工艺中都通过添加这一物质来诱导壳聚糖酶的产生。在液态发酵中,壳聚糖粉、虾壳粉也被作为诱导剂、碳源和氮源添加到培养基中。
为了大量生产壳聚糖酶以满足其商业化需求,培养基配方以及发酵工艺条件的优化非常重要。为优化发酵条件,Zhou 等对碳源、氮源、金属离子、pH值、温度等因素进行了经典的单因素研究。这些多重生长参数间的交互作用使我们很难控制发酵结果。响应面方法学(RSM)是一种非常有用的统计学技术,可用于优化复杂的化学、生物和食品的加工过程,在微生物酶生产的研究中也得到了广泛关注,2012年,Zhang等人报道了利用统计学方法优化壳聚糖酶的生产,结果显示采用Aspergillussp. QD-2发酵生产壳聚糖酶,经响应面法优化,壳聚糖酶活性从26.5 U/mL提高到了85.8 U/mL 。
固态发酵通常被定义为微生物在没有或基本没有游离水的固态基质上生长的发酵方式。固态发酵主要用于真菌发酵,得到的代谢物浓度较高,纯化成本较低,Da 等报道了以昆虫病原真菌Trichodermakoningi为菌种,采用固态发酵的方式生产壳聚糖酶,优化后的壳聚糖酶产量达到了4.84 U/g(以培养基干重计),其研究表明了固体发酵对微生物壳聚糖酶生产的可行性。
5 酶的纯化
在制备壳寡糖的工业生产中采用粗酶液,不需要采用纯化的壳聚糖酶,但是为进一步研究该酶的生化特性、功能结构需要对该酶进行纯化。大多数情况下,该酶的纯化采用硫酸铵或聚乙二醇进行分级盐析,通过使用离子交换及亲和技术达到纯化目的,最后通过SDS-PAGE进行蛋白检测纯化。已经有大量关于来自不同细菌和真菌的上清液中壳聚糖酶纯化和特性的研究被报道。Zitouni等从Paenibacillussp.的培养滤液中分离、纯化了一种耐热的壳聚糖酶,采用冷丙酮沉淀、Q-SepHarose FF离子交换的方法从Paenibacillussp.1794的发酵上清液中纯化了比酶活为122 U/mg的壳聚糖酶。
6 壳聚糖酶理化性质
来源不同的壳聚糖酶,其基本生化特性也会有所不同,这些酶的生化特性已经得到深入的研究和报道。
6.1 分子量
大部分壳聚糖酶的分子量较低,目前发现分子量约在20~75 kDa之间。Wang 等从AspergillusfumigatusKH-94中获得一个分子量大约为108 kDa的壳聚糖酶,具有很高的分子量。Zitouni等从PenicilliumchrysogenumAS51D中获得的壳聚糖酶存在2种形式,分别为30,31 kDa。
6.2 最适温度、PH、金属离子
从微生物中分离的壳聚糖酶的最适pH值在4.0~8.0之间。据报道,从Gongronellasp. JG和Aspergillussp. QD-2中分离得到的壳聚糖酶的最适的pH值为5.6,偏酸,紫色杆菌属中分离得到的壳聚糖酶最适的pH值在5.0~7.0之间。Gupta等从Anabaenafertilissima中获到的壳聚糖酶的最适的pH在7.5左右,偏碱性的酶。
壳聚糖酶的最适温度主要在30~60 ℃之间。Johnsen等从Janthinobacteriumsp. 4239中分离出一种冷活性壳聚糖酶在10~30 ℃条件下可以保留30%~70%的酶活性。大多数被报道的壳聚糖酶在自然界中都是嗜温的,然而热稳定的壳聚糖酶很少有报道。众所周知,耐高温的壳聚糖酶在工业生产中有很多优势,高温条件下,反应进程加快,液体黏度降低,微生物污染率也会降低。Chen等报道了从A.fumigates中分离的一种壳聚糖酶,80 ℃下半衰期为205 h,90 ℃下为1 h,100 ℃下为32 min,这种具有热稳定性的酶在工业生产的应用上更具有优势[18]。
不同金属离子对壳聚糖酶酶活性的影响,Mg2+,Ca2+,Zn2 +,Mn2+,K+,Na+和Cu2+等离子是该酶的激活剂,Fe2+,Hg2+,Co2+和Ag+等离子对该酶有强烈的抑制作用。例如Wang等在研究SerratiamarcescensTKU011菌株中得出Mn2+,Cu2 +也可作为该酶的抑制剂。
6.3 酶的水解
以从酶产物中获得的还原端糖的C1质子的异构体为基础,不同家族的壳聚糖酶分为:“retaining”和“inverting”(见图2)两种催化类型。采用“retaining”催化的糖苷水解酶在对底物进行水解时分为两步,发生了一个两步双置换反应,在这个反应中,两个必需氨基酸残基一个用作亲核试剂,另一个作为酸/碱基团参与了反应。与此相反,采用“inverting”催化型的糖苷水解酶在对底物进行水解时发生了一个单置换反应。该反应需要酸/碱基团的辅助,碱性基团极化了一个水分子形成更强的亲核基团去攻击糖苷键上的碳原子,而酸性基团质子化糖苷键上的氧原子来加速这个反应。Fukamizo等发现Glu22是质子供体,Asp40活化水分子以攻击底物中葡萄糖胺上的碳原子。
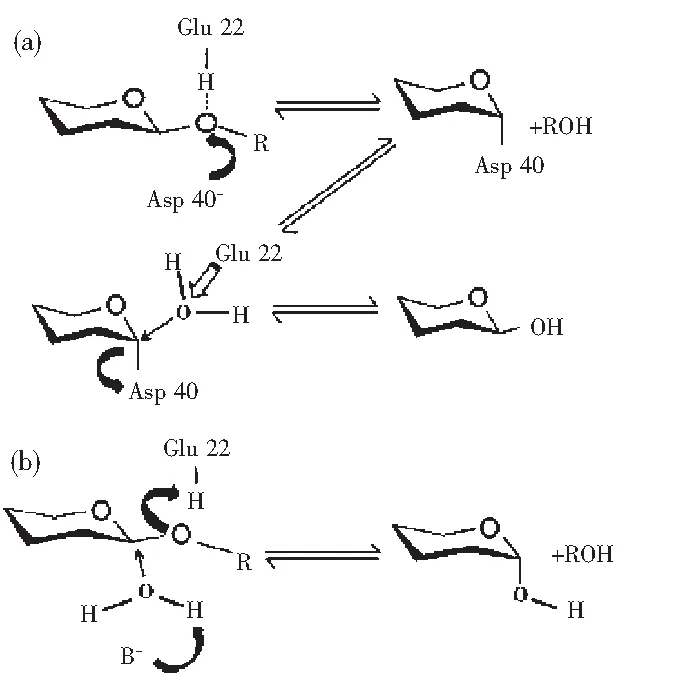
图2 壳聚糖酶催化机制
6.4 酶的诱导剂
大多数来源于微生物的壳聚糖酶都受到胶体壳聚糖的诱导,使用胶体壳聚糖作为培养基补充成分在壳聚糖生产中是一种常见的方法。另外,壳聚糖粉末、鱿鱼粉末和虾壳粉等不同底物能促使壳聚糖酶诱导产生。然而,壳聚糖黏度高,具有抗菌作用,会抑制微生物的生长,在加热杀菌过程中也会与其他化合物发生反应,因此采用不需要壳聚糖的诱导能产生壳聚糖酶的微生物发酵生产壳聚糖酶具有一定的技术优势,非诱导型壳聚糖酶在发酵过程中不需要在培养基中加入壳聚糖作为诱导剂具有一定的技术优势。从Bacillussp.MET 1299中分离出的组成型壳聚糖酶也有类似的报道。
6.5 酶的结构
Marcotte等首次介绍了一种从链霉菌N174中提取的抗真菌壳聚糖酶的x射线结构[19],该壳聚糖酶呈哑铃状,长度55Å,包含两个球状的区域,两个球状区域是由一段氨基酸组成的螺旋连接,形成一个10Å宽、12Å深的裂缝。壳聚糖酶的二级结构主要包括10α螺旋和3个β螺旋,α螺旋约占整个结构的64%。壳聚糖酶的表面电荷分析显示结合裂缝附近主要是负电荷,有利于结合带正电荷的壳聚糖底物,但是几丁质酶的裂缝是中性的。壳聚糖与活性位点的结合表明,底物结合和催化机制可能类似于其他糖水解酶。在壳聚糖酶中确定了6个糖结合位点,糖苷键断裂可能发生在其中两个位点和底物之间的糖苷键相互作用上。x射线结构表明,Glu22和Asp40对催化功能很重要。
随后Saito等[20]报道了从环状芽孢杆菌MH-K1中分离得到壳聚糖酶是一种由259个氨基酸组成的分量为29 kDa的胞外酶。该酶包括14个α螺旋和5个β螺旋,有上部和下部两个球状的结构域,产生底物结合活性位点裂缝。该壳聚糖酶与来源于链霉菌N174的壳聚糖酶在氨基酸序列上仅有20%的相似,但在空间结构上非常相似。通过比对上述两种壳聚糖酶结构,两者有三处明显差异,此外,该酶在Cys50和Cys124之间的二硫键是由β1折叠和α7螺旋连接,这样的结构在其他壳聚糖酶中并不保守。连接两个球状结构域的两条螺旋在方向上不同,这两条螺旋也决定着两种酶的裂缝在大小和形状上的不同。在裂缝活性位点的结构差异是酶特异性识别不同底物并降解不同类型壳聚糖的原因。
7 克隆和遗传改进
通过对壳聚糖酶基因的克隆,科研工作人员对壳聚糖酶的序列、特性、超表达、诱导模式以及酶工程等方面进行了大量研究。不同细菌和真菌中的壳聚糖酶基因已被克隆,主要在Escherichiacoli或Pichiapistoris中表达。近年来,来源于海洋中的Pseudomonassp.OUC1的壳聚糖酶基因被克隆,并在Yarrowialipolytica中实现了高表达,获得的转化株(9.0 U/mL)可以比非转化株(4.3 U/mL)分泌更多的壳聚糖酶[21]。在EscherichiacoliBL21(DE3)[22]中表达的N-末端融合了6×His标签的来源于芽孢杆菌的GH-8家族壳聚糖酶以可溶性形式存在,其表达水平高达500 mg/L,酶活性可达到140 U/mL。该酶可以有效地将壳聚糖转化为游离的壳寡糖:1 g酶可以水解约100 kg壳聚糖。
据报道,耐热的抗真菌B.subtilis壳聚糖酶(csnv26)已在Escherichiacoli中实现异源分泌表达[23],对编码壳聚糖酶的基因csnv26进行扩增并克隆到载体pBAD中,并在Escherichiacolitop10中表达。通过对重组蛋白的SDS-PAGE和酶谱分析,结果显示重组蛋白分子量大小约为27 kDa。重组菌所分泌的蛋白产量高达6.2 g/L。
由于真核基因不易改造,所以有关真菌壳聚糖酶的基因改良方面的报道少于细菌。Liu等人报道了来自Fusariumsolani0114的壳聚糖酶基因在Saccharomycescerevisiae工业菌株中表达并实现大规模生产。Chen等人将烟曲霉菌内切壳聚糖酶基因在PichiapastorisGS115表达,酶活结果显示该酶在80 ℃下半衰期为205 h,在90 ℃下为1 h,在100 ℃下为32 min,是一种热稳定性极强的壳聚糖酶。
Zhu等[24]通过反向pcr克隆了从Penicilliumsp.D-1的壳聚糖酶编码,对其进行cDNA序列分析,在基因中并未发现内含子,因此推断酶由250个氨基酸组成,其中包括由20个氨基酸所组成的信号肽,并且该蛋白序列与TalaromycesstipitatusB8M2R4中的GH75家族的壳聚糖酶相似度达到了83.6%,且这种酶已在Escherichiacoli中实现了大量表达。
8 壳聚糖酶生物学功能及应用
微生物产生的壳聚糖酶具有多种生物学活性,壳聚糖降解微生物广泛分布于自然界中,微生物通过分泌壳聚糖酶降解壳聚糖以达到需求。壳聚糖酶常常和几丁质酶、几丁质脱乙酰酶和氨基葡萄糖酶等酶的共同使用,来降解回收甲壳类动物的贝壳。壳聚糖是接合真菌细胞壁中主要的结构成分,壳聚糖酶在其细胞壁降解和形态中起着重要作用。
壳聚糖酶应用广泛[25],最为重要的用途是生产壳寡糖。壳聚糖酶水解产生的壳寡糖分子量更易控制且条件温和,壳聚糖酶水解获得的壳寡糖分子量一般低于10 kDa,且更易溶于水,优于天然的壳寡糖。壳寡糖比壳聚糖有更好的生物功能,例如在生物医学领域对人体的免疫调节、抗肿瘤、降血脂、调节血糖、改善肝脏和心肺功能等。此外,壳寡糖在食品行业还具有重要的应用价值,有助于改善肠道,从而促进人体吸收营养物质;可用于生产调味品,代替市场上一些如苯甲酸钠等化学添加剂;可用于生产饮料,具有减肥、美容养颜、调节免疫功能等作用;可用于蔬菜、水果的保鲜,且具有抗菌防腐的功效。
大量的海产品工业发展产生的甲壳类废弃物,处理这些废弃物会引发一系列环境问题[26]。近年来,产壳聚糖酶的微生物在甲壳类废料处理中得到广泛的应用,如酶和抗氧化剂的生产。
壳聚糖是真菌细胞壁中主要的结构成分,其中一种应用是利用壳聚糖酶降解细胞壁中的壳聚糖制备原生质体[27]。壳聚糖酶也是壳聚糖介导的基因传递,提高细胞转化效率。
利用微生物或其组分抑制植物病害的生物防治可代替化学杀真菌剂,也是一种生态的、有效的农业病原菌防治方法。几个研究小组报道了壳聚糖酶的体外抗真菌活性,它们可用于提高植物对不同植物病原真菌的抗性[28-30]。Kouzai等人报道了壳聚糖酶活性在植物抗病性中的分子机制。植物病原真菌在感染过程中改变细胞壁成分,避免宿主裂解酶降解,细胞壁几丁质向壳聚糖的转化可能是病原体的感染原因之一。
9 展望
壳聚糖酶不仅是用于制备具有生物活性物质壳寡糖的关键酶,与此同时壳聚糖酶在植物病原体的生物防治方面也发挥着重要作用。壳聚糖酶得到广泛应用依赖于以合理的成本生产高活性酶。近年来,壳聚糖酶相关的研究热点也集中在嗜热壳聚糖酶的开发。其来源主要是通过对嗜热微生物的直接提取或在其基因水平进行修饰而获得具有热稳定性的壳聚糖酶。利用微生物产生的壳聚糖酶处理甲壳质废弃物不仅能消除其对环境的污染,还能增加经济效益。在不久的将来,逐渐深入地对壳聚糖酶的分子机制及生化性质的研究,将促使壳聚糖酶在各行业中得到更加广泛的应用。
