UV-B对大麦幼苗叶片部分生理和光合特性的影响
2018-12-11李晓科武玉珍张义贤
李晓科,武玉珍,张义贤
(1.晋中学院生物科学与技术学院,山西晋中030619;2.山西大学生命科学学院,山西太原030006)
近年来,随着工业化进程的加速,人类生产生活中排放的氟化物分解了大量臭氧,使地球臭氧层破坏,南极出现“臭氧空洞”现象,致使地面紫外线辐射(UV-B)增多,已引起当前各国学者的高度关注[1-2]。关于紫外线辐射(UV-B)增加对植物产生的影响,不少学者已通过模拟UV-B辐射进行了许多方面的研究,并发现UV-B辐射可以影响植物的形态、生物量、光合作用和呼吸作用等[3-8]。
大麦在世界上种植广泛,其蛋白含量高、脂肪和糖类含量低,是开发保健食品的作物来源,也是制造啤酒的重要原材料[9]。大麦还容易培养、生长速度快、对环境胁迫因子较为敏感,是研究环境胁迫因子对作物影响的理想试验材料。目前,紫外线辐射(UV-B)增加对大麦胁迫效应的研究已有较多报道[10-11],但有关UV-B辐射对大麦幼苗生理和光合特性影响的研究报道较少。
本试验以大麦为试验材料,从生理和光合特性角度研究UV-B辐射对大麦幼苗叶片生理和光合特性的影响,旨在揭示UV-B辐射对大麦幼苗的胁迫效应规律,并进一步阐明UV-B辐射对大麦的危害机制。
1 材料和方法
1.1 试验材料
试验所用材料为大麦(晋科571)种子,购于山西省农业科学院农作物品种资源研究所。
1.2 试验方法
1.2.1 材料培养 参考张义贤等[12]大麦种子培养方法,种子露白后整齐放入铺有数层滤纸的培养皿中,加入Hoagland营养液,置于恒温培养箱中培养生长,条件控制为:白天25℃,黑夜20℃,自然光照。
1.2.2 试验设计 UV-B辐射由植株上方包有0.13 mm醋酸纤维素薄膜的紫外灯管产生,处理时间为每天 8 h(9:00—17:00),对照采用自然光照。试验共设4个处理,分别为CK(0kJ/m2),T1(2kJ/m2),T2(4 kJ/m2)和 T3(6 kJ/m2),每个处理设 3 次重复。
1.2.3 测定指标及方法 UV-B辐射处理3,6 d后,分别于当日9:00—11:00随机选取不同辐射处理条件下的大麦幼苗叶片,用Li-6400XT便携式光合测定仪测定净光合速率(Pn)、气孔导度(Gs)、胞间 CO2浓度(Ci)、蒸腾速率(Tr),叶绿素含量、细胞膜透性(CMP)和丙二醛(MDA)含量采用李合生[12]的方法进行测定。细胞膜透性(CMP)用相对电导率(%)表示,采用文献[13]的方法进行测定。

1.3 数据分析
采用SPSS21.0和Excel 2003进行统计分析。
2 结果与分析
2.1 UV-B辐射对大麦幼苗叶片细胞膜透性(CMP)和丙二醛(MDA)含量的影响
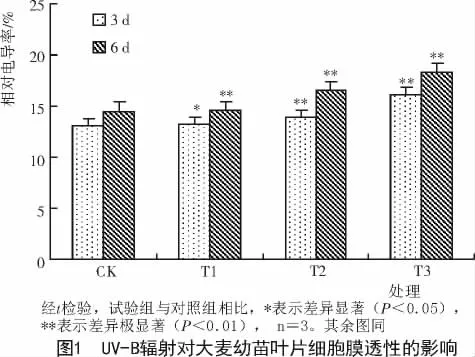
由图1,2可知,大麦幼苗叶片CMP和MDA含量均随UV-B辐射增强而上升,并且二者在T2,T3处理组与对照间差异达极显著。随着处理时间的延长,叶片相对电导率上升较快,在T1处理组就与对照差异达极显著水平,说明此时UV-B辐射对细胞膜透性破坏比较严重。而UV-B辐射处理6 d的大麦幼苗叶片丙二醛(MDA)含量只有在辐射强度较高的T2,T3处理组才快速增加,说明此时细胞膜透性的增加使细胞膜脂过氧化程度加大,大麦幼苗叶片已经受到UV-B辐射的伤害。

2.2 UV-B辐射对大麦幼苗叶片叶绿素含量的影响
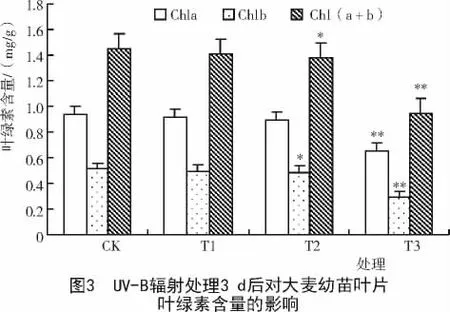
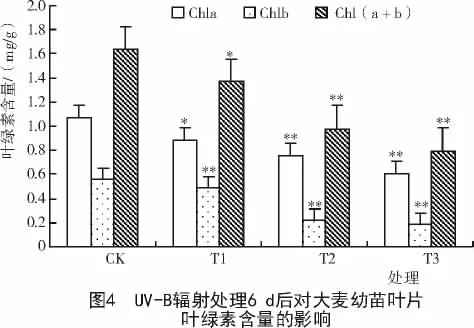
绿色植物中叶绿素含量的稳定对于植物光能的收集、传递和转换起着重要的作用[14]。从图3,4可以看出,大麦幼苗叶片中的叶绿素含量随着UV-B辐射处理强度的增加而下降,其中,UV-B辐射处理3 d的叶片叶绿素a含量和叶绿素b含量在T1,T2,T3处理组分别下降为对照的97.87%,95.20%,69.90%和96.09%,94.72%,56.95%。说明叶绿素b含量下降的幅度大于叶绿素a含量。统计学分析表明,在辐射处理3 d的T3处理组和辐射处理6 d的T2和T3处理组中,大麦幼苗叶片Chla,Chlb和Chl(a+b)的含量与对照间均达极显著差异(P<0.01),且大麦幼苗叶片叶绿素含量随着辐射处理时间的延长而下降。因此,UV-B辐射强度越大,辐射时间越长,对大麦幼苗叶片叶绿素含量抑制作用就越强。
2.3 UV-B辐射对大麦幼苗叶片净光合速率(Pn)和气孔导度(Gs)的影响
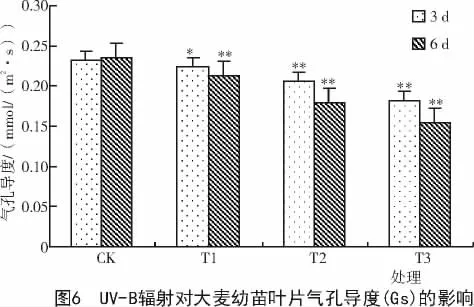
从图5,6可以看出,与CK相比,大麦幼苗叶片净光合速率(Pn)及气孔导度(Gs)随各处理强度的升高呈下降趋势。统计学分析表明,叶片气孔导度(Gs)与对照相比,UV-B辐射3 d的T1处理组表现为差异显著(P<0.05),T2,T3处理组表现为差异极显著(P<0.01),UV-B辐射6 d时3个处理组均表现为差异极显著(P<0.01)。而UV-B辐射3,6 d的净光合速率(Pn)与对照相比,均表现差异极显著(P<0.01)。此外,处理3 d时叶片Pn和Gs在T1,T2,T3处理组分别下降为CK的96.42%,78.85%,49.42%和96.55%,88.79%,78.45%。因此,净光合速率(Pn)下降的幅度大于气孔导度(Gs)下降的幅度,说明UV-B辐射对大麦幼苗叶片净光合速率(Pn)影响较大。从处理时间上看,图5,6表明,经UV-B辐射处理6 d后的大麦幼苗叶片净光合速率(Pn)和气孔导度(Gs)下降的幅度都大于经UV-B辐射处理3 d后的大麦幼苗叶片,说明UV-B辐射处理时间越长,对大麦幼苗叶片的伤害越严重。
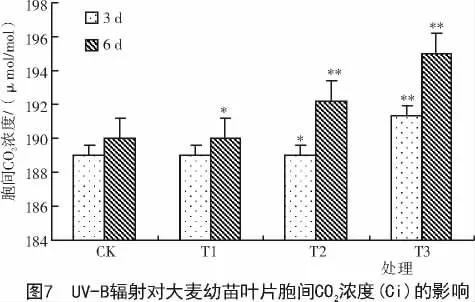
2.4 UV-B辐射对大麦幼苗叶片胞间CO2浓度(Ci)和蒸腾速率(Tr)的影响
从图7可以看出,处理3,6 d的大麦幼苗叶片随UV-B辐射强度增加胞间CO2浓度(Ci)呈上升趋势。统计学分析表明,在UV-B辐射处理3 d时,大麦幼苗叶片胞间CO2浓度(Ci)在T1处理组与对照间无显著性差异,T2处理的幼苗叶片胞间CO2浓度(Ci)显著高于对照幼苗(P<0.05),而 T3处理的幼苗叶片胞间CO2浓度(Ci)与对照达极显著差异(P<0.01)。此外,从处理时间上看,经UV-B辐射处理6 d后的大麦幼苗叶片胞间CO2浓度(Ci)上升的幅度都大于经UV-B辐射处理3 d后的大麦幼苗叶片,其原因可能是UV-B辐射处理时间越长,光合作用利用的CO2浓度在减少所导致。
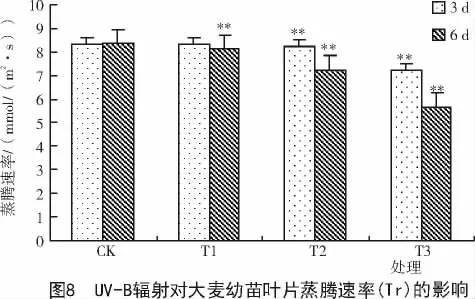
从图8可以看出,处理3,6 d的大麦幼苗叶片蒸腾速率(Tr)随UV-B辐射强度增加呈下降趋势。统计学分析发现,在UV-B辐射处理3 d时,大麦幼苗叶片蒸腾速率(Tr)在T1处理组与对照间无显著性差异,而在T2,T3处理组幼苗叶片蒸腾速率(Tr)与对照间差异均达极显著水平(P<0.01)。从处理时间上看,经UV-B辐射处理6 d后的大麦幼苗叶片蒸腾速率(Tr)下降的幅度都大于经UV-B辐射处理3 d后的大麦幼苗叶片,说明UV-B辐射处理时间越长,大麦幼苗蒸腾速率(Tr)越慢。
3 结论与讨论
照射到地面的太阳光中的紫外线包含UV-A(波长 320~400 nm),UV-B(波长 280~320 nm)和UV-C(波长200~280 nm)。在这3种紫外辐射中,UV-C辐射对植物的伤害最为严重,能使多数被辐射的植物死亡,UV-A辐射基本没有伤害,而大多数植物在UV-B辐射下会产生一些伤害症状[15-16]。
细胞在正常情况下自由基含量很低,对细胞无伤害。可当植物处于逆境时,自由基含量增加,膜结构和保护酶系统受到破坏,积累丙二醛(MDA)[17]。MDA可与蛋白质、核酸等活性物质结合,从而影响细胞正常新陈代谢,其含量能间接反映细胞损伤程度[18]。本试验研究发现,强度不断增加的UV-B辐射使大麦幼苗叶片膜结构受到损伤,细胞膜透性(CMP)增加,积累大量自由基,产生大量过氧化产物丙二醛(MDA),此结果与晏斌等[19]在水稻试验和张新永等[20]在马铃薯试验中所得结果一致。
植物与大气环境进行气体交换的气孔会依据环境条件调节其开度大小,以便获得CO2最大进入量[21-22]。而胞间CO2浓度的变化可作为判断光合速率是否受气孔因素影响的一个依据[23]。本试验发现,随UV-B辐射增强,大麦幼苗叶片胞间二氧化碳浓度(Ci)升高了,但气孔导度(Gs)、蒸腾速率(Tr)和叶片净光合速率(Pn)却在降低,这表明,能用于光合作用的二氧化碳量是充足的,光合速率和气孔导度却下降了,这主要是由非气孔因素引起的[24]。这与董杰等[25]在小麦试验和时丽冉等[26]在小黑麦试验中所得结果相似。本试验还表明,影响植物光合作用的叶绿素含量随UV-B辐射强度增加和辐射时间延长而大幅下降,说明光合速率下降的原因可能是高强度的UV-B辐射导致大麦幼苗叶片叶绿素遭到破坏,致使含量降低,并且使光合酶如PEP羧化酶、RuBP羧化酶活性下降,光系统受到影响,特别是捕光色素,结果使叶绿体能吸收的光能降低[27-29]。
