海拔梯度下元谋干热河谷植物群落特征
2018-12-05余娇娥司宏敏吴雪涛张彧苏文华周睿
余娇娥,司宏敏,吴雪涛,张彧,苏文华,周睿*
1. 云南大学生态学与地植物学研究所,云南 昆明 650500;2. 吉林大学环境与资源学院,吉林 长春 130023
植物群落是植物在长期环境变化中自我调节和适应而形成的集合体(Lieth,1975),物种多样性是群落最基本的特征,是形成群落结构的基础。研究群落的物种组成及多样性是认识植物群落结构、生态、动态的基础(王国明等,2017;许传阳等,2014)。一般认为,物种多样性是由资源可利用程度决定的(Morin,2000),会随着生境的干旱的程度而降低(Bello et al.,2006;刘小丹等,2014)。α多样性以及β多样性构成物种的总体多样性。在植物群落多样性研究中,群落α多样性是反映群落内物种组成状况的一个重要指标,即物种丰富度和均匀度,β多样性表示物种沿环境梯度发生替代的程度或物种变化的速率,表示生境的变异程度或指示生境被物种分隔的程度(马克平等,1995)。植物群落α、β多样性是当前群落生态学乃至整个生态学研究中十分重要的内容(David,1994;马斌等,2008;陈杰等,2012;张晓龙等,2017)。研究植物群落多样性特征对揭示群落、生态系统结构和功能演变具有极为重要的意义(Pandey et al.,2003)。
植被盖度(Vegetation coverage)通常是指植被冠层垂直投影面积占基准地表单位面积的比例或百分比(Purevdorj et al.,1998),是表示陆地表面植被数量及反映生态系统变化的重要基本参数,也是地面蒸散、光合作用等多种地表过程研究的控制性因子,也是反映森林资源的一个重要参数(Price,1993;张云霞等,2003)。植被覆盖及其变化不仅是区域生态系统环境变化的重要指标,而且对水文、生态、全球变化等都具有重要的意义(陈晓光等,2007)。
干热河谷是对分布于中国西南地区,具备干、热两个基本属性河谷带状区域的总称,该区域由于地形深陷封闭而产生的雨影作用以及海拔下降绝热增温和谷地焚风效应而形成了西南地区特殊的地形气候类型(金振洲等,1987)。所谓“干热”就是极端水分条件与热量条件的结合。其中,“干”方面,全年的蒸发量大于降水量的3~6倍;“热”方面,年均温>20 ℃,>10 ℃年积温>7000 ℃。全年干湿季分明,水分和光热条件失衡,热量高、降水量少以及蒸发强是干热河谷气候的典型特征(何永彬等,2000;张建平等,2000)。这一特殊气候特点导致该区域谷底(海拔约1100 m)植被为稀树灌草丛组成的“河谷型”萨瓦纳植被(Savanna of valley type,金振洲,1999),而河谷顶部(海拔约2100 m)则分布有常绿阔叶林。从植物区系来看,1600 m以下河谷中植物有明显的热带性质,而河谷顶部亚热带成分增加(欧晓昆,1988),表现出明显的物种更替现象。剧烈的气候和群落变化被压缩在1000 m的海拔落差中,其间物种适应特征、物种组成和生物多样性也必然发生重大改变。该地区早期就有人类活动,是人类文明的发源地之一,是中国西部人口密集、开发较早的山地农业区,是云南农业经济发展热区(朱钟麟等,2006)。社会资源的高度开发和利用以及其他人为因素(如放牧、开垦耕地等)直接或间接造成了生态环境的进一步退化和破坏(张建利等,2010)。且从 20世纪 50年代至今,该地区大肆开展植物造林工程,主要引进种植了以赤桉(Eucalyptus camaldulensis)、柠檬桉(Eucalyptus citriodora)、大叶相思(Acacia auriculaeformis)以及豆类等物种(李昆等,1999;李昆等,2004),迅速而直接地改变了干热河谷的景观类型、组分结构及空间格局。该区域干湿季分明的气候特征也造就了植被外貌明显的季节性变化,旱季很多物种采用避旱对策呈现假死状态,只有很少完全适应当地极端干热环境的物种能在旱季保持常绿,这些物种是组成当地植物群落的重要物种,是维持生物多样性的基石。此外,雨季有人工种植物种逸出,这些物种未必适宜当地气候,会对生物多样性调查造成一定干扰。
本研究从干热河谷底部到两侧山地沿海拔梯度设置样方,探索沿海拔梯度群落结构的变化。试图回答以下科学问题:该地区的群落结构变化是怎样的?物种替代现象主要发生在哪个海拔区间?如何划分干热河谷与常绿阔叶林植被的分布区域?研究结果不仅可为干热河谷地区物种多样性的深入了解提供科学依据,还可为干热河谷的植被恢复提供依据。
1 材料与方法
1.1 研究区概况
元谋干热河谷位于金沙江流域干热亚区(何毓荣 等 , 1999 ) , 地 理 位 置 25°31′~26°07′N ,101°36′~102°07′E,是金沙江干热河谷的典型区域。属南亚热带干热季风气候。随海拔高度升高及顺河而上气温下降,湿度和降水量增加以及蒸发量减小,辐射增强。谷底干热及沿海拔梯度的水热急剧变化是干旱河谷生态环境的基本特点之一(周志琼等,2008)。年均降雨量为642.2 mm,年平均蒸发量为 3911.2 mm,蒸发量大约是降雨量的6.2倍,表现了干热河谷干旱的特征;多年平均气温为21.9 ℃,≥10 ℃年积温为8003 ℃,年日照时数约为 2670.4 h(崔永忠等,2006)。其气候与稀树草原(Savanna)气候相似(Ratnam et al.,2016)。基带土壤为燥红土,另有变性土、薄层土及紫色土。自然植被以坡柳(Dodonaea viscosa)+扭黄茅(Heteropogon contortus)群系为主(金振洲等,1987)。主要乔木树种有云南松(Pinus yunnanensis)、余甘子(Phyllanthus emblica)、滇刺枣(Ziziphus mauritiana);主要灌木有坡柳、疏序黄荆(Vitex negundo)、小石积(Osteomeles anthyllidifolia)、铁仔(Myrsine africana)、金合欢(Acacia farnesiana)、清香木(Pistacia weinmannifolia);主要草本物种有扭黄茅、孔颖草(Bothriochloa pertusa)。
1.2 样地设置
本研究采用样方调查法,于元谋干热河谷的2017年旱季时期(2017年4月)共设置了10个海拔梯度。从海拔1100 m左右开始,每隔100 m海拔根据地形条件设置4个10 m×10 m的样方。在10 m×10 m的大样方内再设置3个1 m×1 m的小样方内调查草本层。共设置40个10 m×10 m的样方,120个 1 m×1 m的小样方。乔木层记录所有胸径(DBH)≥1 cm的物种及株数、高度、胸径和盖度,灌木层及草本层记录所有种的株数、高度和盖度。胸径用围尺测量,高度和盖度采用目测法。其中,总盖度为10 m×10 m样方内所有植物垂直投影面积占样方面积的比例,灌木层盖度为10 m×10 m样方内灌木植物垂直投影所占的比例,草本层盖度为10 m×10 m样方内草本植物垂直投影所占的比例,分层盖度加和值大于或等于总盖度。同一海拔4个样地的盖度为4个样方的平均值。
1.3 数据分析
1.3.1 数据处理
运用Office WPS(北京金山办公软件股份有限公司和珠海金山办公软件有限公司)对实验数据进行初步整理与计算。运用SPSS 13.0(Chicago,IL,USA)对海拔与物种盖度、物种多样性进行Pearson相关性分析。
1.3.2 α多样性计算
本研究采用了4种常用的物种多样性指数(姜隽,2009;刘雨芳,2003;马克平等,1995;王飞等,2017):丰富度指数(S)、Shannon-Wiener物种多样性指数(H′)、Pielou群落均匀度指数(J)以及Simpson生态优势度指数(D)进行群落特征分析,其计算公式如下:
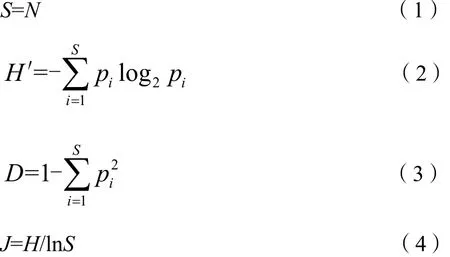
式中,S为物种数目;N为群落中的物种数;pi为属于种i的个体占全部个体的比例。
1.3.3 β多样性计算
该指标是反映不同环境梯下群落之间物种组成的相异性或物种沿环境梯度的更替速率,表示生境的变异程度或指示生境被物种分隔的程度。不同环境梯度中不同样地之间共有种越少,β多样性就越高。一般采用相异性指数和Cody指数进行表征,其计算公式如下(孙荣等,2011):
相异性指数(SI):

Cody 指数(βc):

式中,a和b分别为两样方的物种数;c为两样方的共有物种数;g(H)是沿生境梯度H增加的物种数目;l(H)是沿生境梯度 H失去的物种数目,即在上一个梯度中存在而在下一个梯度中没有的物种数目;相异性指数(SI)反映群落或样方间物种组成的相似性;Cody指数反映物种沿梯度的替代速率。
2 结果与分析
2.1 植被盖度的变化趋势
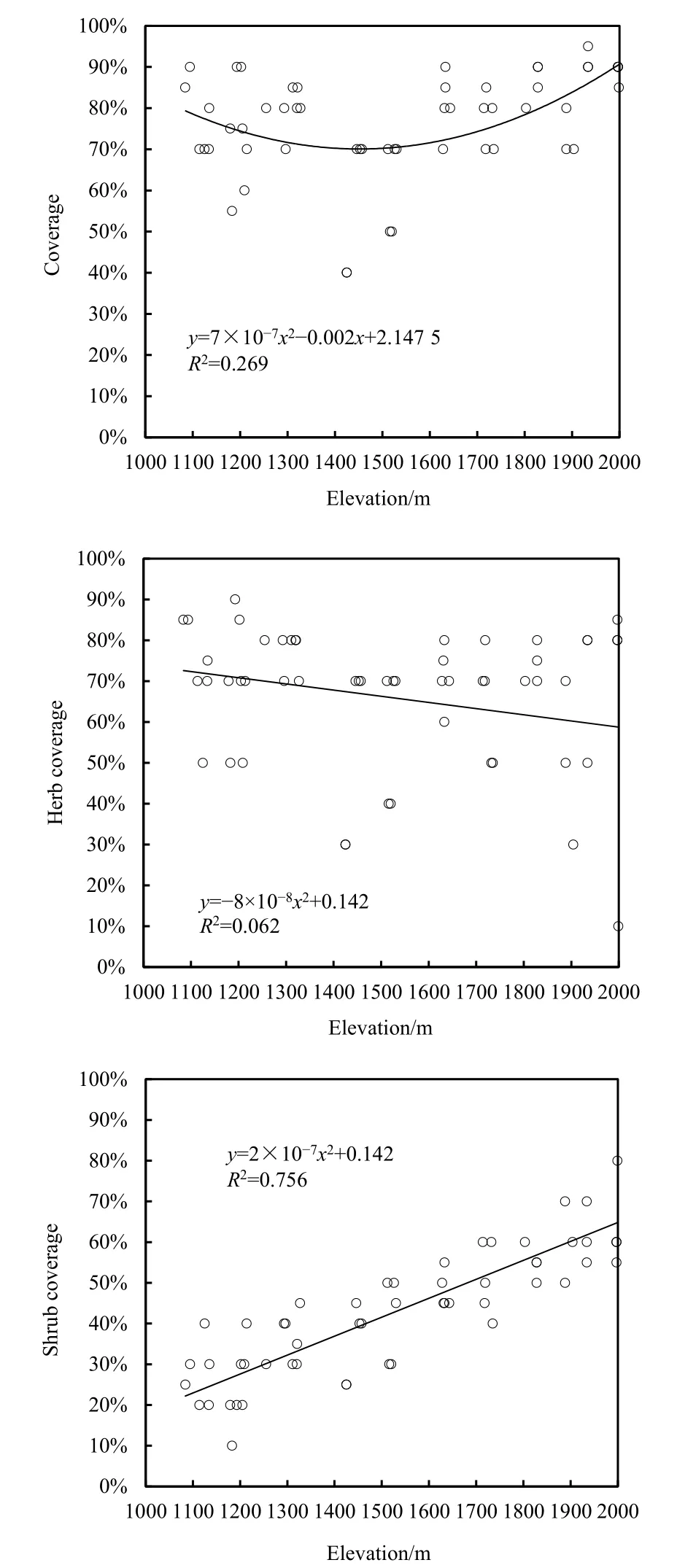
图1 盖度随海拔的变化趋势Fig. 1 Tendency of coverage to change with elevation
由图1可知,随着海拔的升高,群落总盖度与灌木层盖度整体上均呈现出了随之增高的趋势。植被总盖度整体波动相比灌木层的盖度波动大。在海拔1400~1500 m处植被总盖度出现一个急剧下降的趋势,海拔1500 m之后,盖度又有一个回升的趋势,且在海拔2000 m时,植被总盖度达到最大值。灌木层盖度随海拔高度逐渐上升,在海拔1100 m时,其盖度最低,海拔2000 m时,灌木层盖度同样达到最高值。草本层盖度则呈缓慢下降的趋势,但未有较大的变化。表1显示,海拔高度与群落总盖度在P<0.05水平上显著正相关;海拔高度和灌木层盖度在P<0.01水平上呈现极显著正相关;同时群落总盖度和灌木层盖度以及群落总盖度与草本层盖度在P<0.01水平上都存在极显著正相关。由相关性分析可以得知,海拔与总盖度存在显著正相关关系,证明海拔的变化影响了植物盖度的变化,海拔升高,植物总盖度增加。分析其原因,河谷海拔越高,其水热环境越好,越适宜植物生长故植物盖度越高。而灌木及乔木层盖度增加,导致了光热条件的重新配置,使得草本层盖度有所降低。海拔变化引起群落盖度的变化,群落总盖度受灌木层、草本层以及乔木层盖度的共同作用。
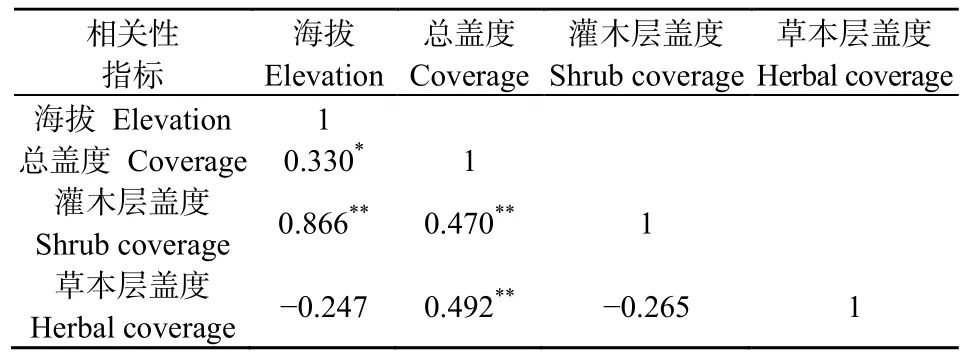
表1 海拔与盖度相关性分析Table 1 Analysis of the correlation between elevation and coverage
2.2 α物种多样性的变化趋势
表2显示了元谋干热河谷不同海拔梯度灌木群落的α物种多样性测定结果。分析可知,Gleason指数(D1)和Marglef指数(D2)变化最大,说明随着海拔升高,灌木群落物种多样性变化较大,且总体呈现出随着海拔升高,物种多样性增强的趋势。
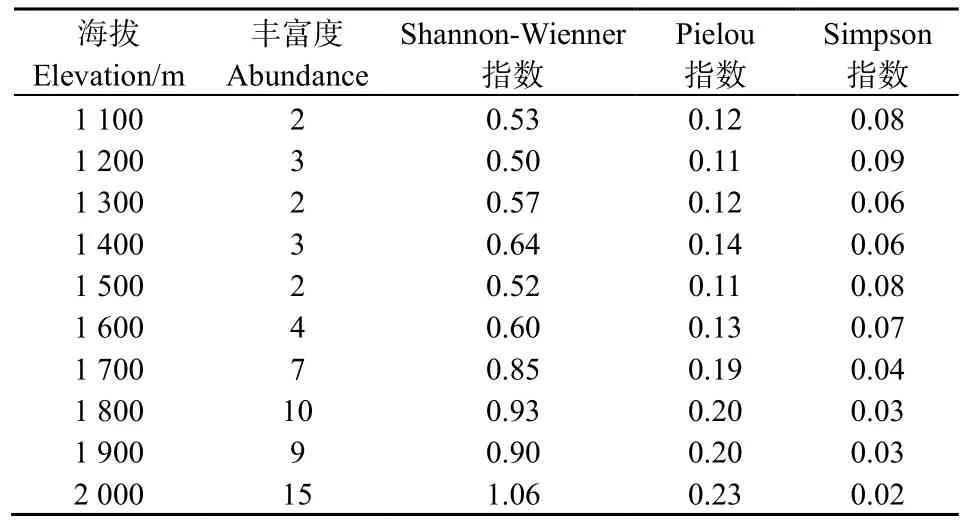
表2 元谋干热河谷不同海拔梯度灌木群落的的a多样性指数Table 2 A diversity index of shrub communities at different elevation in dry-hot valley of Yuanmou
在海拔 2000 m时,物种丰富度达到最大值(15);海拔1100~1500 m时,物种丰富度比较稳定,未发生较大变化。自海拔1500 m之后,物种丰富度迅速上升,且随着海拔的升高呈现出物种丰富度增强的趋势。Shannon-Wiener指数、Pielou指数随海拔变化的趋势基本与丰富度指数一致,在海拔2000 m处物种最为丰富,分布均匀,但Simpson指数的变化趋势与之相反,整体上随着海拔的升高呈现出降低的趋势。
图2显示了元谋干热河谷不同海拔梯度α物种多样性中的Simpson指数和Shannon-Wiener指数的测定结果。从图2中可以看出,Shannon-Wiener指数随海拔梯度整体上呈现出上升的趋势,Simpson指数的变化趋势与 Shannon-Wiener指数的变化趋势大致相反,整体上呈现出随海拔梯度下降的趋势。两个指数随海拔的变化均不呈现出线性的趋势,且均在海拔1500 m处有较大的波动。
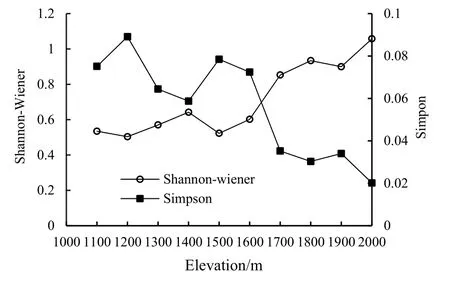
图2 元谋干热河谷不同海拔梯度α多样性变化趋势Fig. 2 Variation trend of alpha diversity at different Elevation gradients in dry-hot valley of Yuanmou
2.3 β物种多样性的变化趋势
元谋干热河谷不同海拔梯度的β多样性如图3所示。在沿海拔梯度上,植物群落SI指数随海拔的增加总体上呈现出“M”型变化趋势。在海拔1400~1500 m之间的相似系数最大,为0.8。而在海拔 1100~1200、1200~1300、1300~1400 m之间的相似系数最小,仅为0.4。Cody指数与SI指数随海拔的变化趋势大致相反,在海拔1400~1500 m之间Cody指数达到最小值(0.5),且随海拔的增加总体上呈现出先下降后上升的趋势。可见,随着海拔的增加,植物群落间的物种更替速率呈先降低再加快,最终达到最高点的特点。
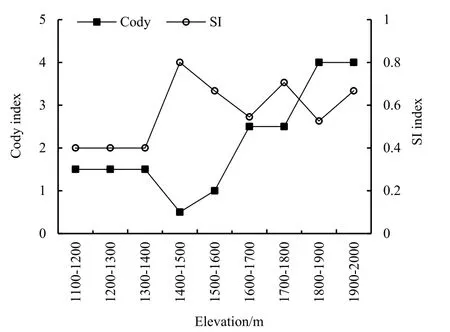
图3 元谋干热河谷不同海拔梯度β多样性变化趋势Fig. 3 Variation trend of beta diversity at different elevation gradients in dry-hot valley of Yuanmou
3 讨论
本研究聚焦于元谋海拔梯度上干热河谷植物的物种多样性。总体上,干热河谷植物群落结构简单,物种组成稀少,10个海拔梯度下的河谷群落植物只有4个群落的物种组成大于5种,这与该地区干旱恶劣的生境条件有关。本研究中,植被总盖度与灌木层盖度总体上呈现出随着海拔的升高,盖度增强的趋势,但两者的波动范围不一致,与王飞(2018)关于白龙江干旱河谷不同坡向主要灌丛群落随海拔梯度变化的物种多样性研究研究结果不一致,造成这种差异的可能原因是元谋干热河谷地区随着海拔的升高,水分越充足(周志琼等,2008),且灌木层物种趋于稳定,以坡柳为主,而木本植物与草本植物两者的变化相对较大。
植物群落多样性指数可以客观反映群落内物种组成特征,在元谋所有海拔梯度下,植物群落 α多样性指数偏低,物种丰富度指数、Shannon-Weiner多样性指数和 Simpson优势度指数最大值分别为15、1.06、0.09,植物群落物种多样性与干热河谷区植被物种多样性水平相当(王飞等,2018;张建利等,2010;杨振寅等,2008)。Shannon-Wiener指数与Simpson指数之间的关系沿海拔呈现出相反的趋势,Shannon-Wiener指数与海拔之间表现出较强的正相关关系,虽然受水热条件以及人为干扰等因素的影响出现波动性,但总体呈现出随着海拔升高,物种多样性增强的趋势(表 2,图 2),与杨阳(2016)关于三江并流地区干旱河谷植物物种多样性海拔梯度格局的比较研究结果,张大才等(2018)关于高寒草甸湿润与干旱化生境中植物多样性分布与植被的关系中的研究结果基本一致。
β多样性用于反映环境梯度下植物群落间的差异性,当一个群落向另一个群落过渡时,环境梯度变化较大,不同群落或样方间共有种越少,β多样性越大(王庆锁等,2000;张晓龙等,2017;韩路等,2017)。本研究中,Cody指数与SI指数之间的变化趋势基本相反。随着海拔的增加,SI相似性系数在海拔 1400~1500 m 的区域达到最大值(0.8),而在 1100~1200、1200~1300、1300~1400 m区域的相似系数最小,仅为0.4。同时,Cody指数在海拔1400~1500 m的区域值最小,为0.5,说明在该海拔区域的 β多样性最小,即在海拔1400~1500 m区域的植物群落结构最稳定,物种更替速率最小。
1400~1500 m总盖度最低,植被稀疏,物种丰富度偏低,推测该海拔是区域内环境条件最为严苛的地段,其稳定性亦说明,只有最适于当地环境的物种得以生存,其他物种难以进入。所以,这个地段的适生物种可以用于当地植被难恢复地区的生态建设。与杨锐等(2015)及宋璐璐等(2011)关于在海拔梯度上各种环境因子会表现出连续的梯度性变化的研究结果不一致,造成这些变化的主要原因可能与该地区的物种替代有关,即在海拔1400~1500 m处具有边缘效应,是干热河谷植被与常绿阔叶林植被的过渡带。同时该研究结果与欧晓昆(1988)关于海拔1600 m是干热河谷植被与常绿阔叶林植被的划分带的结论不一致,这可能是由于人为活动(植树造林、放牧、开垦耕地)的影响而导致过渡带下移。
4 结论
研究表明,元谋干热河谷植物多样性指数、植被盖度与海拔之间均并不呈现连续的线性的变化趋势,且都在海拔 1400~1500 m处出现明显的波动,表明这个区段是生态恢复关键区段,这里的适生物种最适宜在干热河谷环境中生存,是干热河谷中的建群种。
