干旱胁迫对紫丁香幼苗抗氧化系统的影响
2018-12-05路雪梅
李 强, 路雪梅
(东北林业大学园林学院,黑龙江哈尔滨 150040)
水分是限制植物生长和分布的重要环境因子,随着全球气候的暖化,植物生长发育对干旱胁迫的响应和适应受到了越来越多的关注[1]。干旱胁迫诱导植物体内发生各种生理生化反应,造成活性氧的代谢失衡,引起细胞膜脂过氧化,导致生物膜受损,破坏蛋白质、核酸等的分子结构,从而对植物产生毒害[2-3];而增强抗氧化系统是植物体适应干旱胁迫的重要机制[4-5]。抗氧化酶、抗坏血酸和谷胱甘肽是植物抗氧化系统的重要组成部分[5],因此研究干旱胁迫对紫丁香(Syringaoblata)抗氧化系统的影响,对揭示紫丁香的抗旱能力和机制具有重要意义。
紫丁香是我国重要的城市园林绿化植物,具有广泛的应用价值。水资源匮乏是我国城市发展所面临的主要问题之一,在缺水地区紫丁香的栽培应用受到了极大限制[6]。因此研究干旱胁迫下紫丁香抗氧化系统中主要抗氧化酶和抗氧化物的变化规律,了解紫丁香的抗旱性,促进紫丁香在干旱及半干旱地区的推广应用,提高紫丁香在生态恢复和城市园林绿化等方面的应用价值,可为合理应用紫丁香提供科学依据。
1 材料与方法
1.1 材料
紫丁香的种子于2015年在哈尔滨市第二苗圃内采收。盆钵规格为51 cm×38 cm×35 cm(上径×下径×高),盆中装入10.3 kg试验用土。试验所用土壤为黑色泥炭土,购自哈尔滨市和平园林绿化公司,经消毒和过筛处理。试验用土pH值为7.6,有机质含量为145 g/kg,全氮含量为5.6 g/kg,全磷含量为12.3 g/kg,全钾含量为32.8 g/kg。试验前测得试验用土的最大田间持水量为46.7%。
1.2 试验设计
于2015年6月在东北林业大学苗圃内播种,于同年9月移入温室内。2016年5月15日,选择生长发育良好的紫丁香幼苗栽入盆内,每盆内移入1株幼苗,进行正常养护,每处理移植25盆。待移植的紫丁香恢复正常生长后,于2016年7月5日开始控水,试验共设4组处理,分别为对照(CK,土壤水分含量为试验用土最大田间持水量的75%)、轻度干旱胁迫(W1,土壤水分含量为试验用土最大田间持水量的60%)、中度干旱胁迫(W2,土壤水分含量为试验用土最大田间持水量的45%)、重度干旱胁迫(W3,土壤水分含量为试验用土最大田间持水量的30%)。架设防雨棚以防止自然降水对土壤水分的影响,于每日16:00用电子天平称质量,从而控制土壤含水量。7月10日各处理的土壤含水量达到标准,分别于该日之后5、10、15、20 d时取植株中上部叶片测定各项指标。
1.3 测定方法
细胞质膜相对透性和丙二醛(MDA)含量采用孔祥生等的方法[7]测定;过氧化氢(H2O2)含量采用Jiang等的方法[8]测定;超氧化物歧化酶(SOD)、过氧化物酶(POD)、抗坏血酸过氧化物酶(APX)活性采用王学奎的方法[9]测定;过氧化氢酶(CAT)活性采用李合生的方法[10]测定;谷胱甘肽还原酶(GR)活性采用单长卷等的方法[5]测定;还原型抗坏血酸(AsA)含量采用Hodges的方法[11]测定;还原型谷胱甘肽(GSH)含量采用Griffith的方法[12]测定。
1.4 数据处理
对采集的数据采用SPSS 19.0进行统计分析。采用单因素方差分析(One-Way ANOVA)和最小显著性差异法(LSD)检验不同土壤干旱胁迫处理间的差异显著性。
2 结果与分析
2.1 干旱胁迫对紫丁香细胞质膜相对透性的影响
质膜的相对透性常用相对电导率表示,电导率越高表示细胞膜伤害程度越重。如图1所示,在轻度干旱胁迫下,紫丁香的相对电导率在胁迫20 d时与CK相比显著上升(P<0.05),其余处理时间差异不显著。在中度干旱胁迫10~20 d时,紫丁香的相对电导率与对照处理间差异显著(P<0.05)。在重度干旱胁迫下,紫丁香的相对电导率在整个胁迫过程中始终与对照处理间差异显著(P<0.05),同时也显著高于轻度、中度干旱胁迫处理(P<0.05),胁迫20 d时相对电导率已经达到对照处理的2.2倍。

2.2 干旱胁迫对紫丁香丙二醛、过氧化氢含量的影响
如图2、图3所示,在干旱胁迫下,紫丁香的MDA、H2O2含量随着胁迫时间的延长和胁迫程度的加剧而不断上升。在轻度干旱胁迫10~20 d时,MDA含量与对照处理间差异显著(P<0.05);在轻度干旱胁迫15~20 d时,H2O2含量与对照处理间差异显著(P<0.05)。在中度、重度干旱胁迫下,紫丁香的H2O2、MDA含量在胁迫过程中始终显著高于对照处理和轻度干旱胁迫处理(P<0.05)。

2.3 干旱胁迫对紫丁香抗氧化酶活性的影响
如图4-A所示,在轻度干旱胁迫下,随着胁迫时间的延长,紫丁香SOD活性呈上升趋势,在胁迫处理10~20 d时,与对照处理间差异显著(P<0.05)。在中度、重度干旱胁迫下,SOD活性先上升后下降,在中度干旱胁迫下,SOD活性在胁迫15 d后开始下降,但始终显著高于对照处理(P<0.05);在重度干旱胁迫下,SOD活性在胁迫处理10 d时达到最高值,为102.5 U/(min·g),此后开始迅速下降,在胁迫20 d时SOD活性仍显著高于对照处理(P<0.05),但显著低于轻度、中度干旱胁迫处理(P<0.05)。

如图4-B所示,紫丁香的CAT活性在轻度干旱胁迫下始终保持上升趋势,且与对照处理间差异显著(P<0.05)。在中度干旱胁迫下,CAT活性先上升后下降,在处理15~20 d时高于其他各组处理(P<0.05)。在重度干旱胁迫下,CAT活性在处理10 d时达到最高,随后迅速下降,在胁迫20 d时显著低于轻度、中度干旱胁迫处理(P<0.05),但仍显著高于对照处理(P<0.05)。
如图4-C所示,在轻度干旱胁迫下,紫丁香的POD活性呈现上升趋势,在处理15~20 d时与对照处理间差异显著(P<0.05)。在中度干旱胁迫下,POD活性先上升后下降,在处理 15 d 时达到最高,POD活性始终显著高于对照处理与轻度干旱胁迫处理(P<0.05)。在重度干旱胁迫下,在处理5~10 d时,POD活性显著高于其他各组处理(P<0.05);在处理15 d时,POD活性仍显著高于对照处理和轻度干旱胁迫处理(P<0.05),但低于中度干旱胁迫处理;在处理20 d时,POD活性仍显著高于对照处理(P<0.05),低于中度干旱胁迫处理,且差异显著(P<0.05)。
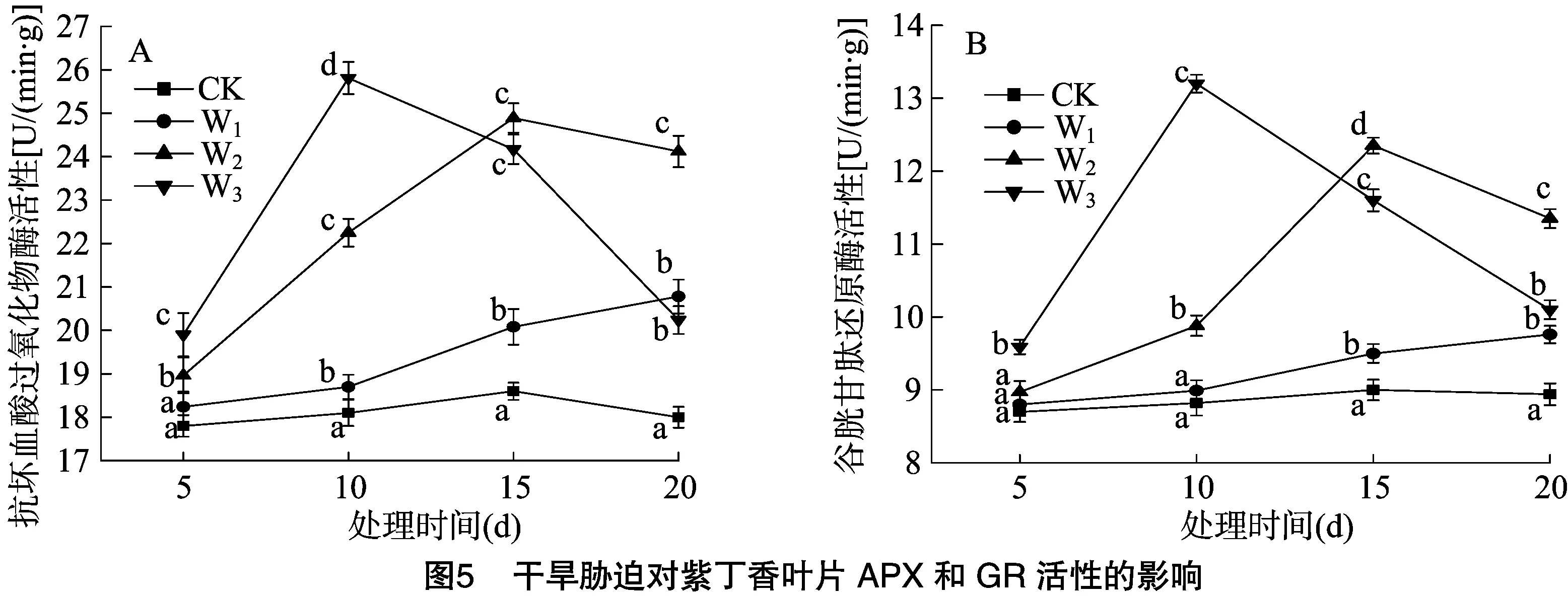
2.4 干旱胁迫对紫丁香抗坏血酸过氧化物酶、谷胱甘肽还原酶活性的影响
如图5-A所示,在轻度干旱胁迫下,紫丁香APX活性在胁迫5~20 d内始终保持上升趋势,在胁迫10~20 d与对照处理差异显著(P<0.05)。在中度干旱胁迫下,APX活性在胁迫5~15 d呈逐渐上升趋势,在胁迫5~20 d内,APX活性均显著高于对照处理和轻度干旱胁迫处理(P<0.05)。在重度干旱胁迫下,APX活性在胁迫5~10 d时,显著高于其余各组处理(P<0.05);此后,随着胁迫时间的延长,紫丁香APX活性开始下降,在胁迫15 d时,紫丁香的APX活性已比胁迫10 d时下降6.4%,但仍与轻度干旱胁迫处理和对照处理间差异显著(P<0.05);在胁迫 20 d 时,APX活性进一步降低,已显著低于中度干旱胁迫处理(P<0.05),但仍显著高于对照处理(P<0.05)。
如图5-B所示,在轻度干旱胁迫下,紫丁香的GR活性于处理15~20 d时显著高于对照处理(P<0.05)。在中度干旱胁迫下,GR活性呈现先上升后下降的趋势,在胁迫15 d时达到最高;GR活性始终显著高于对照处理和轻度干旱胁迫处理(P<0.05);在处理15~20 d时显著高于重度干旱胁迫处理(P<0.05)。在重度干旱胁迫下,GR活性先上升后下降,但始终显著高于对照处理(P<0.05);在胁迫5~15 d时,GR活性显著高于轻度干旱胁迫处理(P<0.05),但在胁迫20 d时二者间已无显著差异;在胁迫5~10 d时,GR活性随胁迫时间的延长而上升,且显著高于中度干旱胁迫处理(P<0.05);但胁迫15~20 d时,GR活性迅速下降,且显著低于中度干旱胁迫处理(P<0.05)。

2.5 干旱胁迫对紫丁香还原型抗坏血酸、还原型谷胱甘肽含量的影响
如图6所示,在轻度干旱胁迫下,随着胁迫时间的延长紫丁香的AsA、GSH含量呈上升趋势,且随着胁迫时间的延长,AsA、GSH含量上升的速度加快;在胁迫15~20 d时,AsA、GSH含量均显著高于对照处理(P<0.05)。在中度干旱胁迫下,紫丁香叶片的AsA、GSH含量均呈现先上升后下降的趋势;在胁迫15 d时,AsA、GSH含量均达到最高值,且与其他各组处理间差异显著(P<0.05);在胁迫20 d时,AsA含量显著高于对照处理与重度干旱胁迫处理(P<0.05),GSH含量仍显著高于其他各组处理(P<0.05)。在重度干旱胁迫下,紫丁香的AsA、GSH含量在干旱胁迫10 d内呈上升趋势,随后开始迅速下降;在胁迫10 d时,AsA、GSH含量显著高于其他各组处理(P<0.05);在胁迫 15~20 d时,AsA含量开始迅速下降,到胁迫20 d时,AsA含量已显著低于其余各组处理(P<0.05);GSH含量在胁迫15 d时,仍显著高于对照处理和轻度干旱胁迫处理(P<0.05),但显著低于中度干旱胁迫处理(P<0.05),在胁迫20 d时,仍显著高于对照处理(P<0.05),但显著低于其他2组胁迫处理(P<0.05)。

3 讨论与结论
在植物的生长发育过程中,干旱胁迫往往是最常见的逆境之一,植物会产生一系列的生理活动以响应和适应干旱胁迫[13]。植物质膜相对透性是反映细胞膜结构和功能的重要指标,干旱胁迫下植物细胞质膜相对透性的增加往往是由细胞过氧化引起的[14]。本研究中紫丁香质膜相对透性与干旱胁迫的强度及胁迫时间长度整体呈正相关关系,表明高强度的干旱胁迫破坏了紫丁香细胞膜结构的完整性,造成质膜相对透性的增加,影响了细胞的代谢和功能,从而诱导紫丁香各器官发生不良反应。
植物体内活性氧的积累是干旱胁迫导致植物质膜损伤和质膜透性提高的根本原因[15],因此干旱胁迫下植物细胞膜受损程度可以通过测量植物体内MDA、H2O2含量进行衡量,质膜过氧化程度越高,MDA、H2O2含量则越高[16]。干旱胁迫下,植物体内的MDA、H2O2含量逐渐上升,但不同植物种类间存在差异,抗旱能力强的植物体内的这2种物质的增速较慢,增幅较小,而抗旱性弱的增加幅度较大[17-20]。本研究中在轻度干旱胁迫5 d时,紫丁香的MDA、H2O2含量均与对照处理差异不显著,而在胁迫10 d时,MDA、H2O2含量均上升,且MDA含量与对照处理间差异显著,说明此时干旱胁迫已对植物造成了一定程度的伤害;在中度、重度干旱胁迫下紫丁香损伤更严重,其体内的MDA、H2O2含量始终显著高于对照处理。
在干旱胁迫下,植物对体内活性氧的清除能力是衡量植物耐旱性的重要因素,抗氧化酶可以降低植物体内活性氧的累积,减缓氧化速度,有效抑制自由基氧化损伤[21-22]。SOD、CAT和POD是植物细胞内重要的抗氧化保护酶,对缓解干旱胁迫下植物体内的过氧化损伤和保持细胞膜结构、功能的完整性有重要作用[23]。干旱胁迫下,植物的抗旱能力与其体内的SOD、CAT、POD活性密切相关[23-26],抗旱性强的植物可以维持较高SOD、CAT、POD活性以缓解干旱胁迫所造成的活性氧伤害[27]。在一定时间和强度的干旱胁迫下植物SOD、CAT、POD活性会上升,而当胁迫时间和强度超过了植物的耐受范围则酶活性会下降[27],这与本研究所获得的紫丁香的这3种抗氧化酶活性在不同强度干旱胁迫下的变化趋势基本一致。说明在长时间和高强度的干旱胁迫下植物的抗逆性下降,作为植物防御体系的抗氧化酶活性也随之下降。
还原型抗坏血酸(AsA)是植物体内含量最丰富的抗氧化剂之一,可以氧化还原过氧化氢,减少自由基的生成[28-29]。APX对AsA的合成和循环代谢起重要作用,保持其代谢活性可以有效提高植物体内AsA含量,因此,APX是植物体内抗氧化的重要酶类[23,30]。本研究结果表明,通过提高还原型抗坏血酸的合成代谢活性,紫丁香幼苗在轻度干旱胁迫下可以有效地增强自身抗性,缓解胁迫伤害。但随着干旱胁迫程度的加剧,紫丁香的APX活性也迅速下降,并破坏了AsA的正常代谢,使其含量降低,这与马玉华等研究的苹果APX活性和AsA含量在不同强度干旱胁迫下的变化趋势[31]相一致,进一步表明在中度、重度干旱胁迫下紫丁香抗逆性下降。
GSH是植物体内重要的抗氧化剂[32],其功能是清除植物体内的H2O2,修复过氧化所致的细胞壁损伤,此外GSH对保持蛋白质或酶的活性以及细胞膜的完整性有积极作用[33]。在植物体内,GR可以调控植物体内的GSH水平,并使细胞保持在还原状态,因此,在干旱胁迫响应中具有重要作用[31]。本研究结果表明,在轻度干旱胁迫下紫丁香的GSH含量和GR活性均上升,但当胁迫强度加剧时,随着胁迫时间的延长GSH含量和GR活性先增加后降低。说明紫丁香幼苗只可以在一定强度和短时间内的干旱胁迫下,通过提高GR活性来增加GSH含量,从而提高紫丁香对干旱胁迫的耐受性,这与李州等研究的白三叶(Trifoliumrepens)叶片可通过提高GR活性和GSH含量来缓解其体内过氧化伤害的结果[34]基本一致。
根据以上分析,紫丁香幼苗可以在一定时间内(20 d)有效地抵御轻度干旱胁迫(土壤水分含量为试验用土最大持水量的60%)所造成的氧化胁迫,但随着胁迫程度的加强其抗氧化系统抵御严重干旱的能力会严重下降。
