橡胶树ADC1的克隆、表达及生物信息学分析
2018-11-30侯岚菲杨洪邓治代龙军门中华李德军
侯岚菲 杨洪 邓治 代龙军 门中华 李德军
(1. 包头师范学院生物科学与技术学院,包头 014030;2. 中国热带农业科学院橡胶研究所/农业部橡胶树生物学与遗传资源利用重点实验室/省部共建国家重点实验室培育基地-海南省热带作物栽培生理学重点实验室/海南省热带作物栽培生理学重点实验室,儋州 571737)
橡胶树(Hevea brasiliensis)原产于亚马逊河流域,是2 000多种产胶植物中唯一商业化种植收获胶乳(天然橡胶原料)的植物。天然橡胶作为四大工业原料之一,是关系国计民生的基础产业和重要战略物资。我国天然橡胶供需矛盾日益凸显,自2009年以来自给率已不足20%。我国属非传统植胶区,橡胶树在生长周期内不可避免会受低温、台风、季节性干旱、病虫害等逆境条件影响。作为橡胶树生长环境中的重要影响因子,逆境胁迫必然对橡胶树的生长、发育和胶乳代谢等造成影响[1-4]。因此,如何有效提高橡胶树对逆境胁迫的耐受性,是橡胶树研究领域中重要且有潜在应用前景的课题。
多胺(Polyamines,PAs)是一类具有强生物活性的低分子脂肪族含氮碱,它可调节植物胚胎发生、根茎生长和衰老、花发育和果实成熟等生理过程[5-8]。PAs生 物 合 成 起 始 于 腐 胺(Putrescine,Put),在由精氨酸脱羧酶(Arginine decarboxylase,ADC)和鸟氨酸脱羧酶(Ornithine decarboxylase,ODC)催化的2条Put合成途径中[9],已报道的多数植物中以ADC合成途径为主。ADC是一种依赖于吡哆醛-5'-磷酸(Pyridoxal 5'-phosphate,PLP)的酶[10],作为PAs合成的重要酶,ADC在调控PAs动态平衡中发挥重要作用,同时ADC还与植物的抗逆相关。继燕麦中克隆ADC基因后[11],又相继从苹果、水稻、桃、棉花、颠茄等物种中克隆和鉴定出[12-16]。ADC基因表达一般无组织特异性,它通过调节PAs代谢参与植物抗逆反应。酸胁迫下大豆ADC活性增加,并与ADCmRNA水平呈正相关[17]。在水稻中过表达曼陀罗ADC基因可提高Spd和Spm水平,进而增强水稻抗旱性[18]。在金柑中,转录因子FcWRKY70通过调控ADC基因表达水平来调节Put合成,以此增强植株抗旱性[19],ADC基因的表达调控PAs的水平,增强杜梨抗旱性[20]。胁迫诱导的AtADC2调节植株体内Put积累,从而增强转基因拟南芥的抗旱和耐盐性,AtADC2突变体对盐敏感需外施Put缓解[21-22]。ADC与枳ICE1相互作用调节PAs水平增强抗寒能力[23]。提高小麦中依赖ADC的Put含量从而降低根细胞壁的铝残留率,增强小麦植株的耐铝性[24]。除与非生物逆境反应相关外,ADC基因还参与细胞凋亡、抗病和防御反应。胡椒ADC1通过调节PAs和γ-氨基丁酸代谢调控细胞凋亡和防御反应[25]。拟南芥ADC基因参与植株对抗黄绿假单胞菌的反应[26]。柑橘中过表达枳PtADC导致转基因植株矮化,气孔密度降低,并极显著地提高了植株对溃疡病的抗性[27]。
鉴于PAs代谢重要基因—ADC在进化和功能上的保守性,推测其可能在橡胶树逆境胁迫耐受性中发挥重要作用。本实验克隆了一个橡胶树ADC基因、研究了其表达模式,为深入解析该基因在橡胶树中的功能奠定基础。
1 材料与方法
1.1 材料
所用橡胶树的组织样品(雄花、雌花、茎尖、树皮、胶乳和叶片)、不同发育时期叶片、过氧化氢(H2O2)、伤害及乙烯利(Ethephon,ET)处理的胶乳样品,均采自中国热带农业科学院试验农场定植的橡胶树品系热研7-33-97。低温、干旱、高盐胁迫处理取材为移栽6个月且长势基本一致的热研7-33-97组培苗叶片。
1.2 方法
1.2.1 材料处理 选取长势基本一致的未开割热研7-33-97进行创伤、H2O2、ET处理,以不作任何处理的未开割树为对照。每个处理时间点取3次重复,每次重复取样5棵树,采集的胶乳在液氮中速冻并保存,用于后续RNA提取。参考Hao等[28]方法,用1%的ET涂于割线及其上方约2 cm割面处,并采集处理0、4、8、24、48及72 h后的胶乳。伤害和H2O2处理参考Zhu等[29]和 Tang等[30]的方法,将2% H2O2浸泡的棉花包在割线及其上方约2 cm割面处,采集处理0、6、24及48 h后的胶乳,用于提取RNA。对橡胶树品系热研7-33-97组培苗进行低温、干旱、高盐胁迫处理,洗净组培苗根部泥土并放入清水中,在温度为30℃、湿度为80%、光照12 h(光照强度 480 μmol·m-2·s-2)及黑暗 12 h环境中静置培养2-3 d,随后进行胁迫处理。参考安泽伟等[31]方法进行低温(4℃)处理,参考刘辉等[32]方法进行干旱及高盐胁迫处理,干旱和高盐胁迫条件分别用30% PEG 6000及1 mol/L NaCl,采集处理0、3、24及48 h后的叶片并立刻用液氮冻存提取RNA。
1.2.2 总RNA提取、cDNA合成和基因克隆 样品总RNA提取用北京百泰克生物技术有限公司的通用植物总RNA提取试剂盒,提取方法参照说明书。用分光光度法进行定量检测,用琼脂糖凝胶电泳进行完整性检测。样品cDNA合成操作参照PrimeScriptTMRT reagent Kit with gDNA Eraser试剂盒说明。
根据橡胶树割面干涸机制实验室获得的部分橡胶树ADC基因序列设计特异性引物(F:5'-GCATA G A A C A A A G G C A C C A A T T G-3'和R:5'-GCATAGA-ACAAAGGCACCAATTG-3')扩增橡胶树ADC基因,并通过测序获得基因序列。
1.2.3 生物信息学分析 开放阅读框预测用NCBI的ORF fi nder;蛋白保守结构域预测用NCBI保守结构域数据库网站(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi);用 Smart分 析 蛋 白保守结构域(http://smart.emblheidelberg.de/);用Expasy网站的Prot-Param程序分析蛋白质理化性质(http://web.expasy.org/protparam/);同源性分析用NCBI中的BLAST和DNASTAR软件;信号肽预测用SignalP 4.1 Server软件(http://www.cbs.dtu.dk/services/SignalP/);用TMHMM 2.0程序预测跨膜结构域(http://www.cbs.dtu.dk/services/TMHMM/);通过在线工具Psort分析目的序列的亚细胞定位(https://www.genscript.com/psort.html)。 用 ClustalX 1.8及MEGA 5.1做多序列比对并构建进化树[33]。
1.2.4 实时荧光定量PCR 根据HbADC1序列设计特异性荧光定量引物(F:5'-GGTTTATACTATGGCAACGAG-3' 和 R:5'-GCACAAAACCAACCATACACG-3'),以不同样品反转录cDNA稀释5倍后作为模板,反应体系为模板2 μL、正反向引物各1 μL、SYBR Premix Ex Taq II(2×)10 μL 及 ddH2O 6 μL。内 参为橡胶树18S rRNA(F:5'-GCTCGAAGAC G A T C A G A T A C C-3'和 R:5'-TTCAGCCTTGCGACC-ATAC-3')。每个样品设置3个生物学重复,基因相对表达量用2-ΔΔCt法计算。用Origin 9.0软件进行数据处理并作图,相对表达量结果为3次重复的x-±s。
2 结果
2.1 HbADC1的克隆及序列分析
在分析橡胶树转录组测序数据时发现一条与植物ADC基因高度一致的序列(TSA接受号:GDFU01108758)。根据该序列设计特异性引物,以橡胶树cDNA为模板进行PCR。扩增产物切胶回收并连接到pMD18-T载体,挑选阳性克隆PCR扩增验证后进行测序。测序结果(图1)表明,该序列长2 594 bp且包含完整ORF,经Blastx比对,确定该基因为植物ADC家族成员,并命名为HbADC1。HbA-DC1最长开放阅读框2 175 bp,编码724个氨基酸。
2.2 HbADC1的生物信息学分析
HbADC1预测理论分子量约为77.7 kD,理论等电点为5.14。HbADC1第119-687位为ADC保守结构域,第130-404位和第436-604位残基分别为鸟氨酸/赖氨酸/精氨酸和相关底物的结合位点,属于2-磷酸吡哆醛依赖性的Ⅳ型(鸟氨酸/二氨基庚二酸/精氨酸)ADC家族[37]。信号肽预测结果(图2)显示,HbADC1无信号肽。亚细胞定位预测结果表明该蛋白属叶绿体运输肽、线粒体靶肽和分泌途径信号肽外的其他类蛋白。
为分析HbADC1与其他植物ADC蛋白的进化关系,通过ClustalX 1.8和MEGA 5.1软件,用N-J(neighbor-joining)法进行1 000次Bootstrap构建橡胶树(Hevea brasiliensis)、木薯(Manihot esculenta)、麻风树(Jatropha curcas)、蓖麻(Ricinus communis)、毛白杨(Populus trichocarpa)、苹果(Malus domestica)、枣树(Zizyphus jujuba)、烟草(Nicotiana tabacum)、拟南芥(Arabidopsis thaliana)、可可(Erythroxylum coca)、 水 稻(Oryza sativa)、 玉 米(Zea mays)、高粱(Sorghum bicolor)、乌拉尔图小麦(Triticum urartu)ADC蛋白系统进化树。结果(图3)表明,HbADC1与木薯 ADC(XP_021610770.1)、麻风树ADC(XP_012084432.1)蛋白为一个分支,与玉米、水稻和小麦等ADC蛋白属不同分支,暗示HbADC1与木薯和麻风树ADC蛋白亲缘关系较近,而与玉米、水稻和小麦等ADC蛋白亲缘关系较远。
2.3 HbADC1的表达分析

图1 HbADC1核苷酸序列及推导的氨基酸序列

图2 HbADC1同源蛋白序列比对
以橡胶树18S rRNA作为内参,对不同组织、不同发育时期叶片、逆境胁迫和乙烯处理条件下的HbADC1表达模式进行实时荧光定量PCR分析。
2.3.1HbADC1在不同组织及叶片不同发育时期的表达分析 实时荧光定量PCR结果(图4)表明,HbADC1在橡胶树不同组织(叶片、雌花、雄花、树皮、胶乳、茎尖)中均有表达,雌花中表达最高,接下来是茎尖、雌花、叶片、树皮和胶乳。
HbADC1在叶片不同发育时期表达存在差异,淡绿期表达量远高于其他4个时期,古铜期、变色期和衰老期表达量相当,稳定期表达量最低(图5)。
2.3.2HbADC1在不同逆境处理条件下的表达分析与其他植物ADC基因在逆境条件下表达模式一致,低温、干旱、伤害、高盐及H2O2处理均能调控HbADC1表达,各处理HbADC1表达模式各异。
低温胁迫下,HbADC1表达呈现先微降后显著上升模式(图6-A)。干旱处理后,HbADC1表达水整体呈升高趋势,24 h持续上调并在48 h保持平稳(图6-B)。伤害处理,HbADC1表达先上升,后下降再上升,处理后24 h和48 h表达水平均低于处理前水平(图6-C)。HbADC1的表达受高盐胁迫调控,HbADC1表达在高盐胁迫后先下降,随后上升并在24 h达到最高后又下降(图6-D)。HbADC1的表达还受H2O2诱导,处理后的表达水平先升后降,表达最高和最低点分别出现在6和48 h(图6-E)。综合HbADC1在逆境条件下表达模式,推测HbADC1参与橡胶树多种逆境胁迫反应,并在其中发挥重要作用。
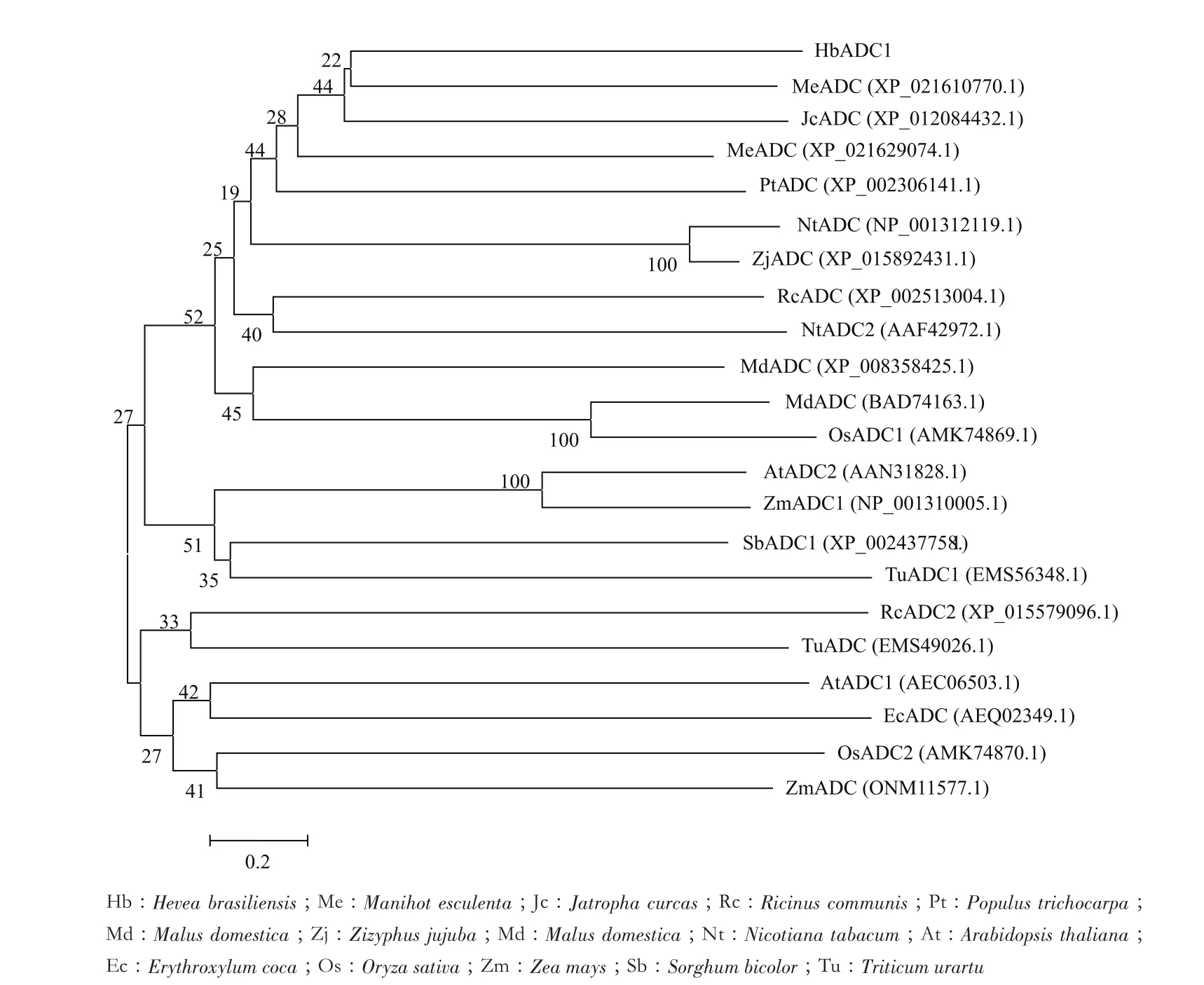
图3 HbADC1与其他植物ADC蛋白的系统进化分析
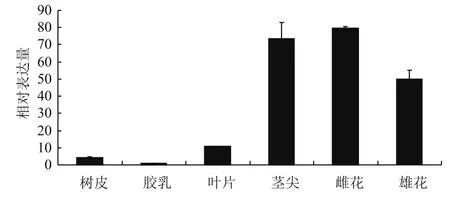
图4 不同组织中HbADC1的表达
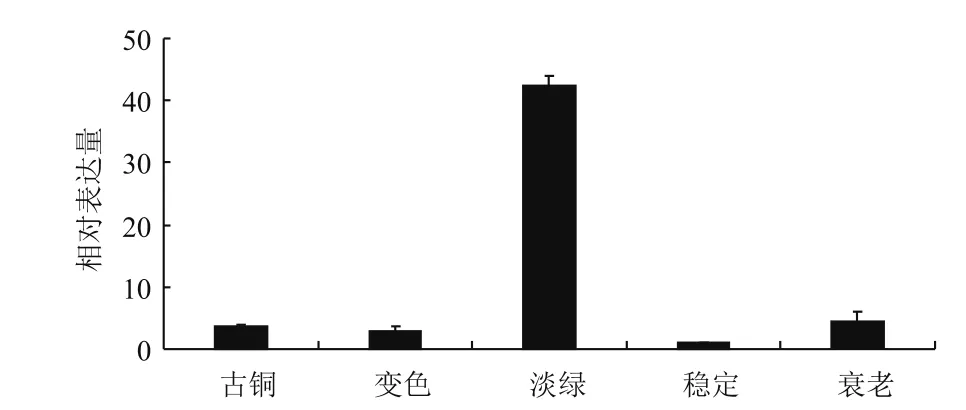
图5 叶片不同发育时期HbADC1的表达
2.3.3HbADC1在乙烯处理下的表达分析 ET是橡胶树中研究最深入且应用最广的植物激素,作为橡胶树增产刺激剂,ET通过延长排胶时间提高橡胶树胶乳产量。实时荧光定量PCR结果(图7)显示HbADC1表达受ET调控。ET处理后HbADC1表达存在波动,呈先升后降再升模式,但整体呈上升趋势,处理后各时间点表达量均高于处理前,8 h达到表达高峰,上述结果暗示HbADC1参与橡胶树ET反应。
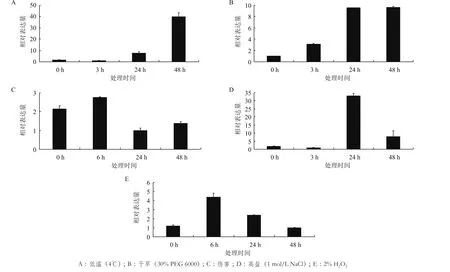
图6 不同胁迫下HbADC1的表达
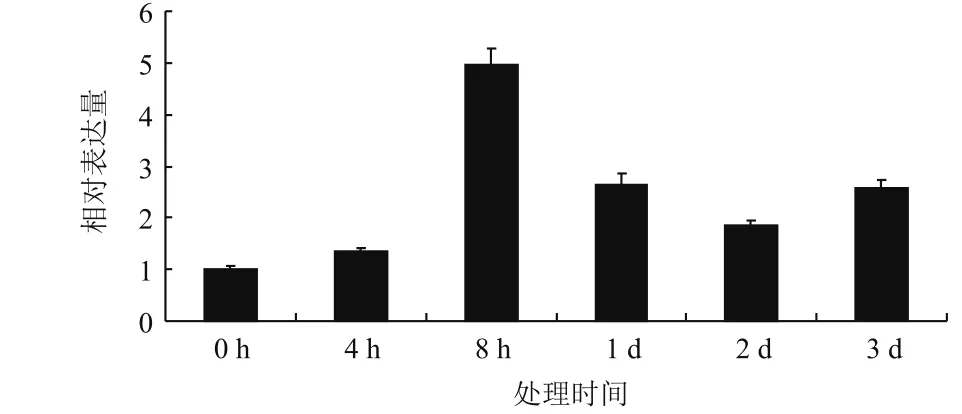
图7 ET处理下HbADC1的表达
3 讨论
ADC是PAs合成途径中一种依赖于PLP的重要酶。本研究克隆了一个橡胶树的ADC基因,HbADC1序列长2 594 bp,最长ORF序列长2 175 bp,编码724个氨基酸。与其他植物ADC蛋白一致,HbADC1包含ADC保守结构域,具有鸟氨酸/赖氨酸/精氨酸和相关底物的结合位点,属于2-磷酸吡哆醛依赖性的Ⅳ型ADC家族[34],与已鉴定的植物ADC蛋白相同[12,22]。以上结构域和特征对ADC蛋白行使功能至关重要,推测HbADC1具有ADC蛋白活性,可行使ADC蛋白功能。拟南芥AtADC1和AtADC2在细胞质和叶绿体中均表现出双重亚细胞定位[35],与此不同,亚细胞定位预测HbADC1不定位于叶绿体,属其他类蛋白。当然,HbADC1亚细胞定位预测结果还有待于进一步实验确认。橡胶树属大戟科,进化分析显示HbADC1与大戟科植物木薯、麻风树等ADC蛋白为同一分支,与玉米、小麦和水稻等ADC蛋白属不同分支,表明其与大戟科植物木薯、麻风树等ADC蛋白亲缘关系较近。上述结果与传统分类一致,暗示HbADC1与上述植物ADC蛋白功能相似。
本研究发现HbADC1表达无组织特异性,该结果与其他植物ADC基因组织表达模式相似,如桃树ADC基因在根和老叶中表达量最低,在嫩枝和幼叶中的表达高于花和茎[14];棉花ADC基因在叶片的表达量远高于根和茎,在根中表达量最低[16];芥菜ADC基因主要在根和茎中表达,在叶片中几乎不表达[36];AtADC1为组成型表达,而AtADC2仅在莲座叶和角果中表达[37]。此外,HbADC1在橡胶树叶片淡绿期表达远高于其他4个时期,暗示HbADC1与橡胶树叶片发育密切相关。
植物ADC活性调节依赖于生理条件,且与多种非生物胁迫有关[18-21,23-25,27]。拟南芥中 ADC 活性低会减少PAs合成,从而降低植株耐盐性[38]。PbrMYB21在烟草中过表达使ADC表达上升,PAs积累增强了烟草对脱水和干旱胁迫的耐受性[20];拟南芥和芥菜中ADC活性分别受渗透胁迫和盐胁迫调控[36,39];AtADC2在细菌侵染后表达上升,蛋白活性增加[26]。与上述结果相似,本研究中HbADC1表达受干旱、低温、伤害、高盐和H2O2等逆境胁迫调控,表明HbADC1可能参与巴西橡胶树逆境胁迫反应。ET通过延长排胶时间提高胶乳产量[40]。本研究发现,ET处理后HbADC1表达存在波动,但整体呈上升趋势,暗示其可能参与橡胶树ET反应和产排胶过程。
4 结论
克隆获得橡胶树HbADC1。其蛋白属于2-磷酸吡哆醛依赖性的Ⅳ型ADC家族,与大戟科植物木薯、麻风树ADC聚为一支。HbADC1无组织特异性表达。HbADC1可能参与橡胶树的叶片发育过程及橡胶树逆境胁迫和乙烯应答过程。
