猪MITF-M的转录调控分析
2018-11-30张廷焕
张廷焕,柴 捷,陈 磊,龙 熙*
(1. 重庆市畜牧科学院,重庆 402460; 2.农业部养猪科学重点实验室,重庆 402460)
家畜在长期的驯化过程中受到了不同选择,毛色呈现出不同的变化[1]。毛色在动物生理调节和环境适应过程中起着重要作用,比如不同毛色会导致热交换出现差异,从而影响动物体温的调控[2]。除此之外,在家畜育种中,毛色更是作为一种经济性状受到强烈的人工选择。家猪毛色变异复杂,如果能明确毛色变异分子机制,可以通过遗传育种手段定向改变家猪毛色,形成特定品种特征,提高群体的表型一致性。
毛色是由表皮、毛囊内黑色素细胞的分布和活性决定的[3-4]。小眼畸形相关转录因子-M型(microphthalmia associated transcription factor M,MITF-M)选择性表达于黑色素细胞中,控制着黑色素细胞的发育、功能和凋亡,在黑色素生成通路中居于核心位置[5-6]。MITF-M还调控着3大主要色素合成酶包括酪氨酸酶(TYR)、酪氨酸相关蛋白1(TYRP1)和酪氨酸相关蛋白2(TYRP2)[7]的转录。目前在多个物种(比如,鸡、牛、羊)上已证明,MITF-M对动物毛色变化起着至关重要的作用[8-10],而猪毛色全基因组关联分析发现,MITF基因位点在不同程度毛色白化的猪种中受到强烈选择,比如,巴马香猪、宁乡猪、五指山猪等。同时,在荣昌猪中的研究证实,MITF-M转录调控区一个位点的插入突变引起了SOX蛋白的结合,影响了MITF-M的转录活性降低其表达水平,最终导致猪毛色白化现象[11]。然而,猪MITF-M转录调控区域的结构和功能尚不清楚,关于猪MITF-M主要转录调控区域及其转录因子的研究尚未见报道。
本试验以猪DNA为模板,通过PCR扩增得到MITF-M的转录调控区域;采用生物信息学对调控区的序列特征进行分析;根据序列结构特征分布情况构建5′端缺失报告基因重组载体;利用双荧光素酶报告基因试验对重组载体进行转录活性分析,寻找活性差异的调控区域;再利用干扰RNA技术手段对差异区域内潜在的转录因子结合位点进行验证。本试验结果可为猪MITF基因的表达机制研究奠定基础,也为深入研究猪毛色变化的调控机理提供理论支持。
1 材料与方法
1.1 试验材料
1.1.1 试验动物 采集3只荣昌猪新生小母猪耳组织样,提取基因组DNA,用于扩增MITF-M基因的转录调控区。荣昌猪来自重庆市畜牧科学院安富猪场。
1.1.2 细胞 B16细胞(小鼠黑色素瘤细胞系)、贴壁细胞购于上海酶研生物科技公司。
1.1.3 试剂 DNA提取试剂盒、RNA提取试剂盒、Q5超保真DNA聚合酶和内切酶均购自NEB公司;蛋白酶K、GoTaq®qPCR Master Mix和双荧光素酶报告基因系统试剂盒购自Promega公司;胶回收试剂盒和质粒提取试剂盒购自Omega公司;干扰RNA购自Ribobio公司;Lipofectamine 2000购自Invitrogen公司;琼脂糖、DNA marker购自TaKaRa公司。
1.2 试验方法
1.2.1MITF-M基因转录调控区克隆以及5′端缺失重组载体构建 本试验以荣昌猪DNA为模板,采用PCR扩增得到6 414 bp的MITF-M转录调控区域。50 μL反应体系:5×Q5 reaction buffer 10 μL,dNTP mixture(2.5 mmol·L-1)1 μL, Q5高保真酶(10 U·μL-1)0.5 μL,正、反向引物(10 μmol·L-1) 各1.5 μL,ddH2O 35.5 μL。PCR反应程序:98 ℃ 30 s;98 ℃ 10 s,60 ℃ 30 s,72 ℃ 4 min,35个循环;72 ℃ 5 min。为了进一步产生后续的克隆片段,将XhoI和NheI两个酶切位点加到引物的5′端。再以6 414 bp片段为模板,成功扩增另外8个5′端缺失片段,大小分别为1 140、1 397、 2 470、 3 753、5 201、5 429、 6 036 和 6 274 bp。随后将所有片段经过XhoI和NheI酶切后,利用T4连接酶亚克隆到pGL3-basic骨架载体上,构建了9个调控区5′端缺失重组载体:pGL3-1140、pGL3-1397、pGL3-2470、pGL3-3753、pGL3-5201、pGL3-5429、pGL3-6036、pGL3-6274、pGL3-6414,再通过测序和酶切电泳鉴定(图1),确认载体构建成功。所用引物信息见表1。
1.2.2MITF-M转录调控区生物信息学分析 从ENSEMBL(http://www.ensembl.org)数据库中得到了其他4个物种(人、鼠、牛、狗)的MITF-M转录调控区序列;在线软件Neural Network Promoter Prediction(http://www.fruitfly.org/seq_tools/promoter.html)用于预测MITF-M转录调控区核心启动子;MEME在线软件(http://meme-suite.org/tools/meme)发掘5个物种间MITF-M转录调控区保守的序列(Motif);在线软件JASPAR(http://jaspar.genereg.net/)用于预测转录因子结合位点。
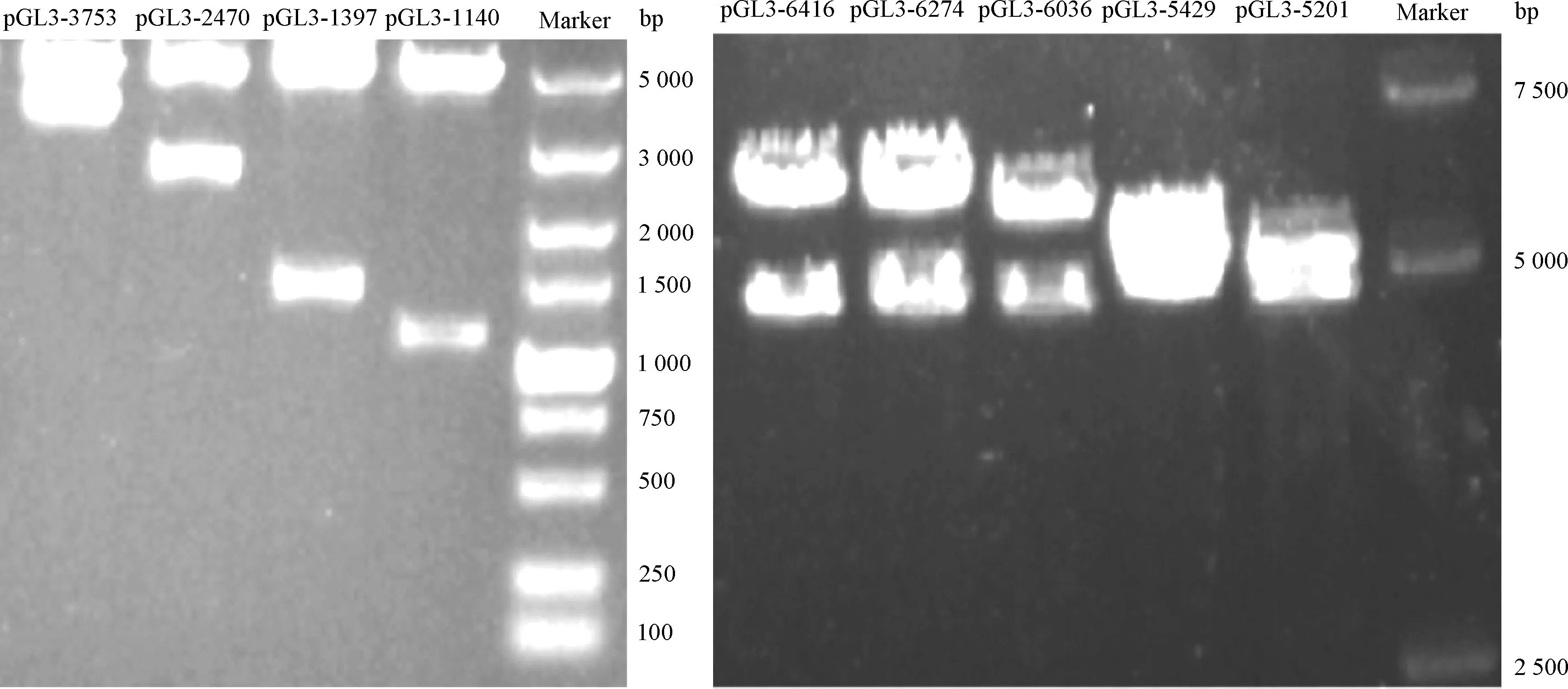
图1 猪MITF-M转录调控区域重组载体酶切电泳图
Fig.1 Enzyme digestion electrophoresis of pig MITF-M transcriptional regulatory regions
表1MITF-M重组载体构建引物
Table1PrimersforthevectorsconstructionofMITF-M

引物名称Name引物序列(5′→3′)Sequence产物长度/bpLength重组载体Recombinant plasmidTSSR+Xho ⅠCCGCTCGAGCGGccggaaactttatcacagcagctc-1140F+Nhe ⅠCTAGCTAGCTAGtgtctggaccctttatgaaactcaca1 140pGL3-1140-1397F+Nhe ⅠCTAGCTAGCTAGaagcttactctgataaccccaactta1 397pGL3-1397-2470F+Nhe ⅠCTAGCTAGCTAGatggccactttatattaaggactttg2 470pGL3-2470-3753F+Nhe ⅠCTAGCTAGCTAGcaaacagtggatggactaggaggact3 753pGL3-3753-5201F+Nhe ⅠCTAGCTAGCTAGgcattttcattcttttaacggctgag5 201pGL3-5201-5429F+Nhe ⅠCTAGCTAGCTAGagtcccaaactcctaatttatccttc5 429pGL3-5429-6036F+Nhe ⅠCTAGCTAGCTAGgcttctacatgataggaaaattggat6 036pGL3-6036-6274F+Nhe ⅠCTAGCTAGCTAGcatacacacttctatgaattcctcct6 274pGL3-6274-6414F+Nhe ⅠCTAGCTAGCTAGctttccagagttgctggaagttttgt6 414pGL3-6414
引物序列中的大写字母为限制性酶切位点;TSS代表转录起始位点
Uppercase letters in the primer sequences are restriction sites andTSSrepresent transcription start site
1.2.3MITF-M转录调控区活性分析 采用脂质体LipofectamineTM2000转染试剂将构建好的MITF-M转录调控区域9个重组载体(每孔1 000 ng)和阴性对照pGL3-basic(每孔1 000 ng)分别与内参载体pRL-TK(每孔700 ng)共转染到B16细胞内,48 h后裂解细胞,采用Promega公司的双荧光素酶活性检测试剂盒及Thermo公司的荧光和化学发光检测仪(Thermo Scientific FLuoroskan Ascent FL)来测量分析。用每个孔的海肾荧光素酶发光值(pRL-TK)为内参校正萤火虫荧光素酶发光值(重组载体),得到每个重组载体的转录活性值,再除以阴性对照pGL3-basic的活性值获得每个重组载体的相对转录活性值。
1.2.4 转录因子KLF4和Meis1干扰RNA试验 由于在本试验研究过程中筛选出可能影响MITF-M转录的两个调控因子,故设计了KLF4和Meis1基因干扰RNA的试验(表2),分别将其与重组载体和内参载体pRL-TK共转染到B16细胞内,48 h后收集细胞分成两份,一份用于上述的双荧光素酶活性检测,获得重组载体的转录活性值;另一份用于提取RNA,反转录成cDNA,再进行RT-PCR检测干扰RNA的基因干扰效率,10 μL反应体系:GoTaq®qPCR Master Mix(2×)5 μL,正、反向引物(10 μmol·L-1)各0.2 μL,CXR 0.1 μL,cDNA 1 μL,ddH2O 3.5 μL。反应程序:95 ℃ 10 min;95 ℃ 15 s,60 ℃ 30 s,40个循环,其中KLF4正向引物:5′-GCCGCTCTCGCTTATCTTCG-3′,反向引物:5′-GTCTTGCTACCCTGCTCCTT-3′;Meis1正向引物:5′-AAGCAGTTGGCACAAGATACAGGA C-3′,反向引物:5′-TGACTGCTCGGTTGGACTGG-3′。采用2-△△Ct方法计算KLF4和Meis1基因表达量。
表2KLF4和Meis1基因的干扰RNA序列
Table2TheinterferingRNAsequencesofKLF4andMeis1genes

名称Name序列(5′→3′) Sequence正义链 Sense反义链 AntisensesiKLF4-1CACCCACACUUGUGACUAUTTAUAGUCACAAGUGUGGGUGTTsiKLF4-2GGUCAUCAGUGUUAGCAAATTUUUGCUAACACUGAUGACCTTsiMeis1-1GCUUUAAAGAGAGAUAAAGTTCUUUAUCUCUCUUUAAAGCTTsiMeis1-2AUGAAGAUAUAGCGGUGUUTTAACACCGCUAUAUCUUCAUTT
1.2.5 统计分析 使用Excel2013进行试验数据整理,用SPSS19软件中的Student′st-test或ANOVA进行差异显著分析,使用Publisher2013作图。
2 结 果
2.1 物种间MITF-M转录调控区生物信息学分析
本试验以荣昌猪DNA为模板,采用巢式PCR成功扩增出MITF-M转录起始位点(TSS)前6 414 bp调控区域。把6 414 bp的DNA序列输入在线软件Neural Network Promoter Prediction,预测发现4个核心启动子,都位于TSS~-1 427 bp内(表3)。
随后构建了猪与其他4个物种(人、小鼠、牛、狗)MITF-M6 414 bp调控区序列的进化树,发现猪和牛的亲缘关系较近,与小鼠最远(图2b)。同时,利用MEME软件分析5个物种间保守的序列模块(Motif),每一个Motif都可能存在与转录调控区域功能相关的调控序列。MEME分析结果表明(图2a),5个物种MITF-M转录调控区域序列存在规律性簇状排列的Motifs,比如,TSS~-1 100 bp区域内包括Motif 1、3、4、7、8、11、13,除了小鼠缺失Motif 8外,其他Motifs在物种间分布保守;在-1 200 ~-1 400 bp区域内也包括3个分布保守的Motif 2、5、10;而从-1 400 bp到-4 830 bp,Motif 6、9、12、14、15在物种间的分布出现较大的差异,同时小鼠缺失Motif 6、12、15。
通过上述生物信息学的分析发现,TSS~-1 400 bp区域存在大多数核心启动子和保守的Motifs,暗示这个区域对MITF-M的转录调控至关重要。
表3猪MITF-M调控区域内核心启动子分布
Table3ThedistributionofcorepromotersinMITF-Mregulatoryregion

起始位点/bpStart结束位点/bpEnd启动子序列(5′→3′)Promoter sequence 分数Score (0~1)-1 427-1 377CTCAATTATTTAAAAAGGCACTGACAATAAAAGCTTACTCTGATAACCCC0.98-1 262-1 212AACATTTATTTTTAAATGTGCAGCCCTTTCTTTTTTAAGTGACATAGTGA0.86-1 001-951ATTCTTGTGCTTAAAATACCTCACCATTTCAGCAGTGAAAATCGGCCATT0.84-535-485AGGATGTTTGTACATATATCCAGCTGAGAATAAGATGCACATTGAGACCA0.88
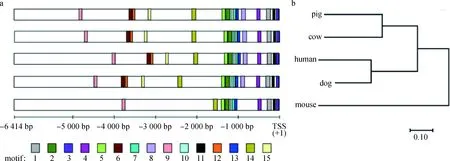
a.MEME数据库预测的保守Motif的分布情况,Motif 1至Motif 15表示调控区域存在的序列模块(Motif),保守的Motif用相同颜色表示,平均长度为50 bp,不同的颜色代表不同的Motif(TSS代表转录起始位点);b. MEGA7构建的物种间系统发育树
a.The distribution of conserved Motifs were predicted by MEME database, Motif 1 to 15 showed sequence modules existing in regulatory regions and conservative Motif had the same color, different Motifs were represented by different colored boxes, the average length of the Motif was 50 bp(TSS represented transcription start site); b. The Neighbor-joining phylogenetic tree was constructed using MEGA7 with 6 414 bp sequences of MITF-M
图2 5个物种MITF-M 6 414 bp转录调控区的系统发育树以及Motif分布情况
Fig.2 Phylogenetic tree and Motif composition of 6 414 bp promoter region of MITF-M in 5 species
2.2 猪MITF-M基因转录活性分析
为了确定猪MITF-M基因的重要转录区域,本试验根据上述核心启动子和Motif的分布情况,成功构建出9个5′端缺失重组载体,分别为pGL3-1140、pGL3-1397、pGL3-2470、pGL3-3753、pGL3-5201、pGL3-5429、pGL3-6036、pGL3-6274、pGL3-6414(图1)。
通过细胞转染试验和双荧光素报告基因试验,得到了每个重组载体的相对转录活性值。如图3所示,所有9个重组载体的转录活性都高于空载体pGL3-basic,但随着插入片段5′端延长,MITF-M调控区域的转录活性发生了变化。相比pGL3-1140,pGL3-1397的活性有显著的上升,说明片段-1 140~-1 397 bp内含有正向调控元件;pGL3-1397和pGL3-2470的活性没有显著差异,都具有很高的转录活性;而pGL3-2470和pGL3-3753之间的活性出现了显著的下降,甚至高达5~6倍的活性变化,说明-2 470~-3 753 bp这个区域可能存在极强的负向调控元件;pGL3-3753、pGL3-5201、pGL3-5429之间的活性没有显著差异,而且维持较低的转录活性;但与pGL3-5429相比,pGL3-6036活性呈现显著的上升,说明片段-5 429~-6 036 bp内含有正向调控元件;而pGL3-6036、pGL3-6274、pGL3-6414之间活性没有显著差异。
由此,鉴定出猪MITF-M基因3个重要的转录活性显著变化的区域,两个转录激活区域,一个转录抑制区域。
2.3 转录因子KLF4调控猪MITF-M基因转录活性
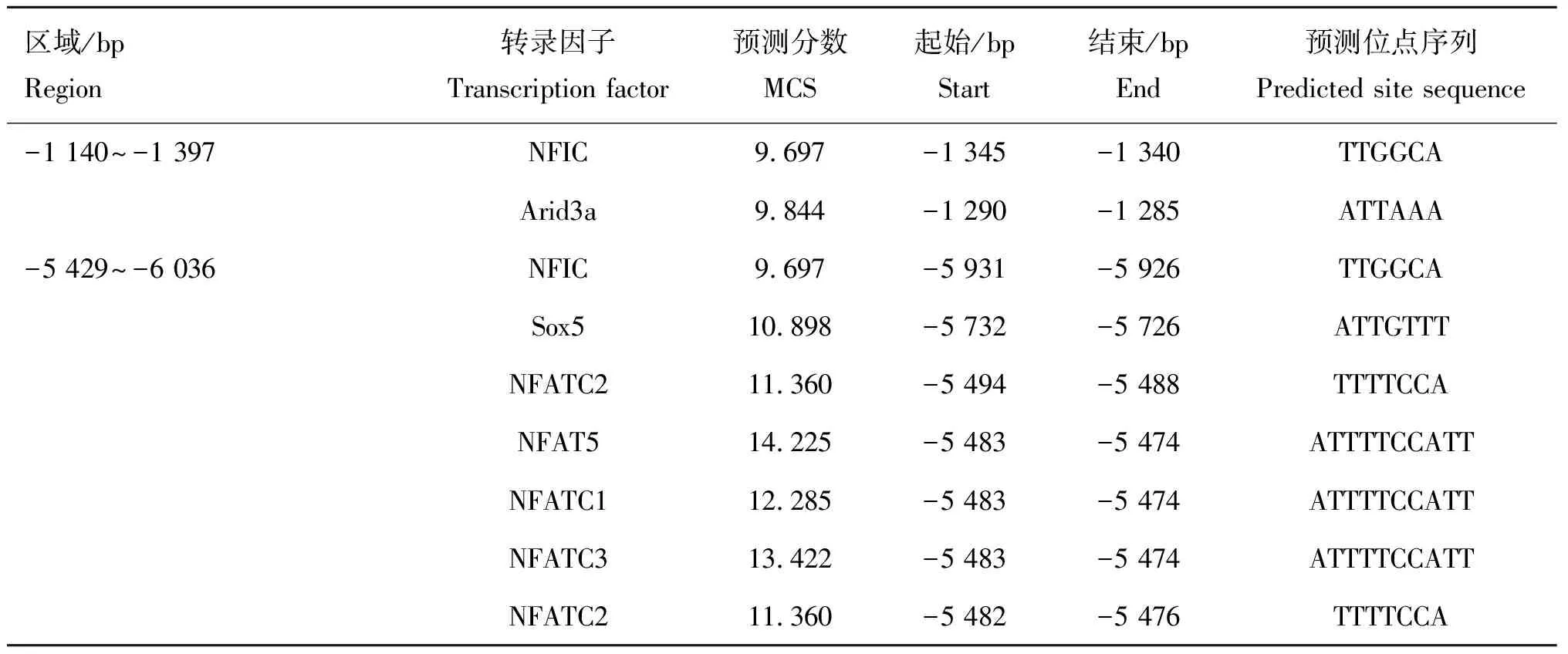
为了进一步确定上述显著变化区域内的重要调控元件和转录因子,本试验运用JASPAR软件预测这3个区域中潜在的转录因子结合位点(表4)。转录因子KLF4预测得分最高,而且已有研究报道KLF4与黑色素的合成直接相关[11];再结合前面MEME分析结果,发现只有转录因子Meis1的结合位点位于Motif(Motif 12)上;而且KLF4和Meis1均未报道参与MITF-M转录调控。
为了证实它们与MITF-M的转录调控关系,本试验合成了KLF4和Meis1基因的干扰RNA各2条(siKLF4和siMeis1)(表2),将其转入B16细胞48 h 后,利用RT-PCR发现基因干扰效率均在90%以上(图4),其中siKLF4-1和siMeis1-2的干扰效率更高,可用于后续干扰试验。随后,MITF-M重组载体和干扰RNA(siKLF4-1或siMeis1-2)共转染试验发现,与对照组(无义干扰RNA)相比,干扰KLF4基因表达时,pGL3-2470 与pGL3-3753活性的比值有显著的下降,由原来约5倍的比值下降到约3倍,说明KLF4确实负向调控这个区域的活性变化。然而,干扰Meis1后,pGL3-2470 与pGL3-3753活性的比值却没有发生显著变化,说明Meis1对这个区域的活性没有调控作用(图5)。
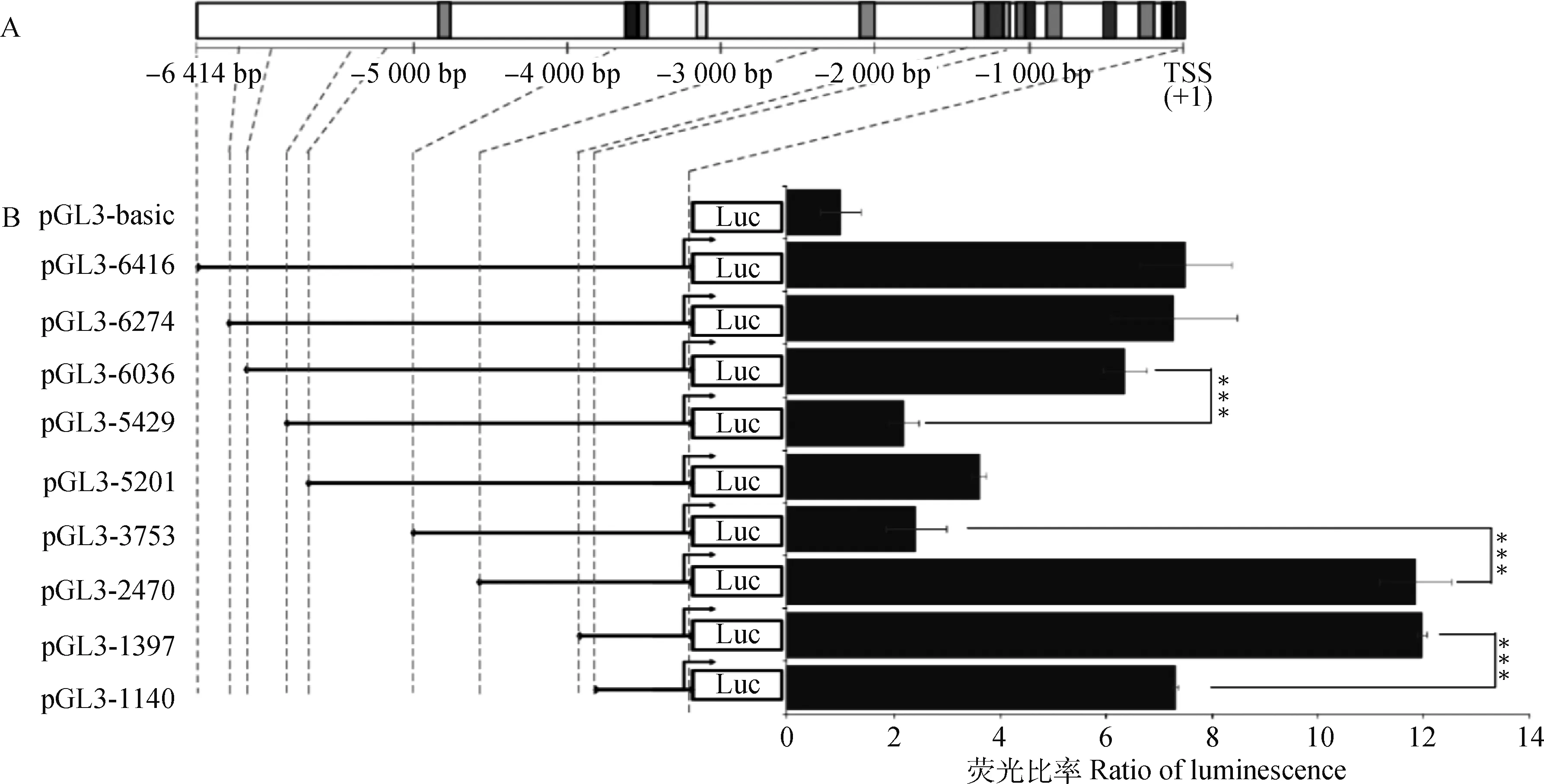
A. 猪MITF-M调控区Motif分布图以及5′截短片段示意图;B.9个5′截短片段重组载体以及空载体pGL3-basic的转录活性,***表示P<0.001
A. The diagram of Motif distribution and 5′ truncated fragments in the regulaory region of pig MITF-M gene; B.The transcriptional activity of recombinant vectors containing 9 5′truncated fragments and pGL3-basic, *** indicate P<0.001
图3 猪MITF-M调控区转录活性分析
Fig.3 The transcriptional activities of the pig MITF-M regulatory regions
由此,本试验发现,KLF4可以通过两种不同的结合序列对MITF-M转录活性起负向调控作用。
表4MITF-M转录调控区转录因子的预测结合位点
Table4TheputativebindingsitesoftranscriptionfactorsinthetranscriptionalregulatoryregionsofMITF-M

区域/bpRegion转录因子Transcription factor预测分数MCS起始/bpStart结束/bpEnd预测位点序列Predicted site sequence-2 470~-3 753KLF415.416-2 836-2 827TGGGTGTGGCBhlha1513.595-2 846-2 839CCATATGTSox510.898-2 768-2 762ATTGTTASox510.898-2 739-2 733ATTGTTTMeis110.323-3 526-3 520TTGACAGNFIC9.697-3 177-3 172TTGGCAMZF19.085-2 664-2 659TGGGGASOX108.910-2 816-2 829CTTTGTMAFG::NFE2L18.812-2 840-2 835CATGAC
(续表4 Continued)
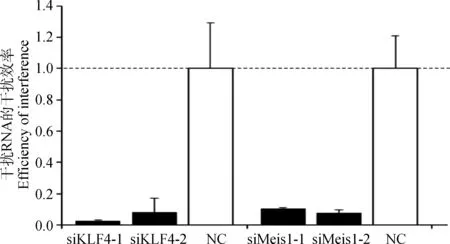
图4 KLF4和Meis1干扰RNA在B16细胞中干扰基因表达的效率
Fig.4 The interference efficiency of siKLF4 and siMeis1 in B16 cells

**表示差异显著(P<0.01),下同
** indicate the significant difference(P<0.01),the same as below
图5 KLF4 和Meis1干扰RNA对MITF-M转录活性的影响
Fig.5 Effect of siKLF4 and siMeis1 on the transcription activity of MITF-M

NS表示差异不显著(P>0.05)
NS indicate the difference is not significant(P>0.05)
图6 KLF4负向调控MITF-M转录
Fig.6 Negative effect of KLF4 on the transcription activity of MITF-M
3 讨 论
毛色是猪的一个重要形态学特征,由表皮、毛囊内黑色素细胞的分布和活性决定。而MITF-M基因控制着黑色素细胞的色素合成和沉积,被视作毛色主要调控因子。MITF-M调控着大量与色素合成和黑色素细胞分化相关的基因,同时还控制着黑色素瘤细胞的增殖、浸润和转移。Chen等[11]发现,敲除MITF-M的转基因小鼠,毛色出现全白的现象,与此同时毛色相关基因的表达水平也发生显著变化。同时已有大量研究表明,MITF-M的转录调控区与动物毛色的改变有着密切的联系,在狗毛色的研究中发现,MITF-M的转录调控区4个位点的突变与狗的毛色变化高度相关[12-13];马MITF-M转录调控区近端的出现一个位点的插入突变导致了马毛色变化[14-16];不同毛色的猪全基因组关联分析证明,MITF所在区域与毛色高度相关[17-18],而且猪MITF-M转录调控区远端出现一个沉默子,导致MITF-M的表达缺失,从而引起毛色白化现象。这一切都证明MITF-M的转录调控区对动物毛色的重要性。本试验获得了猪MITF-M6 414 bp的转录调控区,生物信息学分析发现,MITF-M的启动子都位于TSS~-1 427 bp内,符合大多数基因启动子的分布,而这个区域包含了绝大多数Motifs的分布,后续的双荧光素报告基因试验也证明了pGL3-1397的转录活性是最高的,说明这段区域对于启动MITF-M的转录极为重要,这与Chen等[18]对MITF-M的研究结果相符。本研究中,在MITF-M基因的-1 140~-1 397 bp之间转录活性有显著的变化,但当干扰到结合在Motif 2上的转录因子的表达后,pGL3-1397的活性显著上升,说明物种间保守的Motif 2可能对MITF-M转录起着重要作用。本试验还发现,在pGL3-2470和pGL3-3753之间存在着一个高达5~6倍的转录活性变化,后续试验证实,干扰KLF4表达后,活性比值只减小到3倍,说明 片段-2 470~-3 753 bp之间还存在其他转录因子结合位点,有待进一步挖掘。除此之外,-5 429 ~-6 036 bp之间也出现转录活性显著变化,但此区域并未发现Motif的分布,说明这有可能是猪特有的调控区域。
目前研究发现,PAX3、SOX10、CREB等转录因子和典型的WNT3A信号通路都能够调控MITF-M的转录和表达[19-20]。本试验发现了KLF4与MITF-M的转录调控关系。KLF4(Kruppel-like factor 4)作为真核生物转录因子,是一种在人类多种组织中广泛表达的转录因子,参与调控细胞增殖、分化、胚胎发育等重要生命过程[21]。KLF4是一个双重功能转录因子,根据不同的靶基因,通过不同的机制,既可以激活又可以抑制转录作用。但KLF4在不同的肿瘤中是作为癌基因或肿瘤抑制因子起作用的[22]。本试验发现,在小鼠黑色素瘤细胞中转录因子KLF4对MITF-M基因转录有抑制作用,符合前人的研究结果。同时KLF4是体细胞重编程过程中诱导多能干细胞(iPS细胞)的重要诱导因子之一[23],通过与转录调控区结合元件(GC盒、CACCC盒和基础转录元件)结合来调控靶基因的转录[24]。本试验发现,KLF4通过两个结合位点来调控MITF-M转录活性,一个是近端(-1 140~-1 397 bp)的CACCC盒,一个是远端(-2 470~-3 753 bp)的GC盒,完全符合已有研究结果。除此之外,Peagaricano等[25]研究发现,KLF4的表达量与黑色素的生成有直接关系,与本试验的研究结果相结合可推测,KLF4通过抑制MITF-M的转录活性从而影响黑色素细胞的色素合成和沉积。
4 结 论
本研究发现了猪MITF-M基因3个重要的转录调控区域,2个正向调控区域,1个负向调控区域,还确定了KLF4抑制MITF-M转录活性的调控关系,为进一步研究MITF-M基因调控猪毛色的分子遗传机制奠定了理论基础。
