广西红水河外来红腹罗非鱼形态差异分析
2018-11-30汪开成吴志强谭虹雨赵立朝刘博文赵学倩高胜男程光平
汪开成,吴志强,武 琳,谭虹雨,赵立朝,刘博文,赵学倩,高胜男,程光平,张 曼,黄 钧
(广西大学动物科学技术学院/广西高校水生生物健康养殖与营养调控重点实验室,广西 南宁 530004)
【研究意义】红腹罗非鱼,曾用名齐氏罗非鱼(Tilapiazillii),学名为Coptodonzillii(Gervais,1848),隶属鲈形目(Perciformes)、丽鱼科(Cichlidae)、Coptodon属[1]。该物种原产于以色列和埃及,1978年我国从泰国引进养殖,但因其生长速度慢,个体小,现生产上已被其它罗非鱼品种替代[2]。红腹罗非鱼性成熟早、繁殖周期短、产卵量高且具有较强的生态适应性,目前已成功入侵我国南方部分内陆水域,在生态系统中具有明显竞争优势,威胁到本土物种的生存及生态平衡[2-5]。红水河是珠江流域西江水系的干流,自广西河池市天峨县进入广西境内,至来宾市象州县石龙镇三江口与柳江汇合,流域面积广,水量充沛,渔业资源丰富[6]。国内外研究表明,外来物种能否成功入侵与该物种的生理特性密切相关,入侵物种在新的生态环境中通常会产生一定程度的形态差异[7-9]。红腹罗非鱼虽在广西区内未有正式的引种记录,但前期调查研究发现其已扩散到红水河沿线水域。因此,研究和分析广西红水河不同地理条件下红腹罗非鱼的形态差异,对深入了解该物种生态适应性的分化特征、探讨其入侵现状和危害风险评价等均有重要意义。【前人研究进展】近年来,针对入侵鱼类在不同生境条件下的形态分化已有不少研究成果。万安等[8]通过对大别山区入侵种绿太阳鱼(Lepomiscyanellus)进行形态测量和数据分析,弄清了该群体内产生形态差异的主要原因及其为适应山区溪流环境而产生的体型变化状况;魏朝军[9]探讨了青海省黄河段外来麦穗鱼(Pseudorasboraparva)形态特征与气候变化的关系,表明气温和降水量等环境因子的变化对各样点麦穗鱼雌雄个体的形态分化有不同程度的影响;管嘉俊等[10]对广西6个养殖区域的逃逸尼罗罗非鱼(Oreochromisniloticus)进行了多变量形态度量学研究,通过主成分分析和判别分析均可较好地识别逃逸尼罗罗非鱼来源,并能通过全长、体长和体质量的相关性分析评估其野外生存状态。【本研究切入点】国内虽已有包括红腹罗非鱼在内的不同罗非鱼养殖品种间的形态差异比较[11],但对同一河流不同生境条件下外来红腹罗非鱼的形态差异分析至今未见报道。【拟解决的关键问题】以广西红水河天峨、东兰、巴马、大化、都安、合山和来宾7个地理群体的红腹罗非鱼为研究对象,通过传统形态学和框架分析方法,系统探究红腹罗非鱼不同地理群体间的形态差异,了解该入侵种在不同生存环境下的野外生存状态,为广西红水河鱼类多样性保护及制定红腹罗非鱼入侵防控策略提供理论依据。
1 材料与方法
1.1 试验材料
供试红腹罗非鱼为分别通过拉网捕捞、垂钓和市场采购方法,于2017年3月至2018年5月间采自广西红水河7个区域的河流、水库以及当地农贸市场。各采样点的地理分布、坐标及采集样本数详见图1和表1(各采样点的地理名称均以拼音首字母表示,例如:“天峨”以“TE”表示)。样本采集后及时速冻并用冰袋保鲜运回实验室,为避免雌性红腹罗非鱼因处于不同的怀卵期导致的体型变化对实验结果产生影响,通过解剖观察性腺以判断性别的方法,最终筛选出865尾雄性个体作为研究样本。
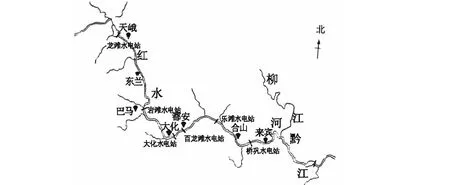
图1 采样点的地理分布Fig.1 Geographic distribution of sampling site
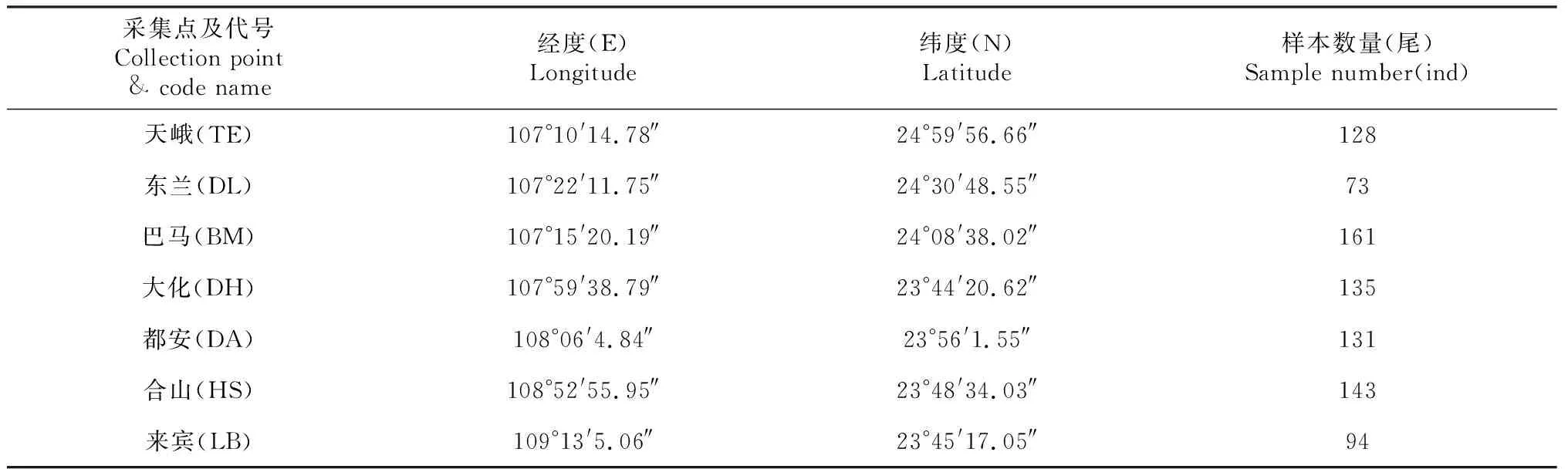
采集点及代号Collection point& code name经度(E)Longitude纬度(N)Latitude样本数量(尾)Sample number(ind)天峨(TE)107°10'14.78″24°59'56.66″128东兰(DL)107°22'11.75″24°30'48.55″73巴马(BM)107°15'20.19″24°08'38.02″161大化(DH)107°59'38.79″23°44'20.62″135都安(DA)108°06'4.84″23°56'1.55″131合山(HS)108°52'55.95″23°48'34.03″143来宾(LB)109°13'5.06″23°45'17.05″94
1.2 体表形态测量
可量性状和框架性状采用精确度为0.01 mm的电子数显卡尺测量。其中,可量性状包括全长、体长、体高、体厚、头长、吻长、眼径、躯干长、尾长、尾柄长和尾柄高共11项指标。框架测量定位点的选择和参数标注按照李思发等[12]的设定,共测量24项框架性状数据(图2),每一项均以“大写字母D+阿拉伯数字”表示,如:D1-2表示下颌骨最后端至吻前端的直线距离(mm)。
1.3 统计分析
为消除异速生长及体型差异对结果分析的影响,除全长外,其它形态数据均除以全长并取对数进行标准化处理[12],校正后仍以原性状命名(例如:log1/2体长/全长仍称为体长),最终得到34项标准化性状,其中包括10项可量性状和24项框架性状。应用统计软件SPSS 22.0对相关数据进行处理分析。
1.3.1 主成分分析 对各地理群体红腹罗非鱼的34项标准化性状进行主成分分析,通过不断剔除信息提取率(即主成分分析中所提取原始数据的有效信息比例)低于60 %的性状,筛选最优数据组合。采用KMO检验和巴特勒球度检验验证所选性状是否适合进行因子分析(KMO>0.6,P<0.01)。建立原始数据的得分系数矩阵并选取特征值大于1.0的因子作为主成分,采用最大方差正交旋转法对因子负荷矩阵进行旋转以获得各主成分的贡献率和累积贡献率[13]。对贡献率较高的前3个主成分(PCA1、PCA2和PCA3)的得分系数进行具体分析,随后对贡献率大于10 %的主成分绘制散点图以探讨不同地理群体间红腹罗非鱼的形态区分度。
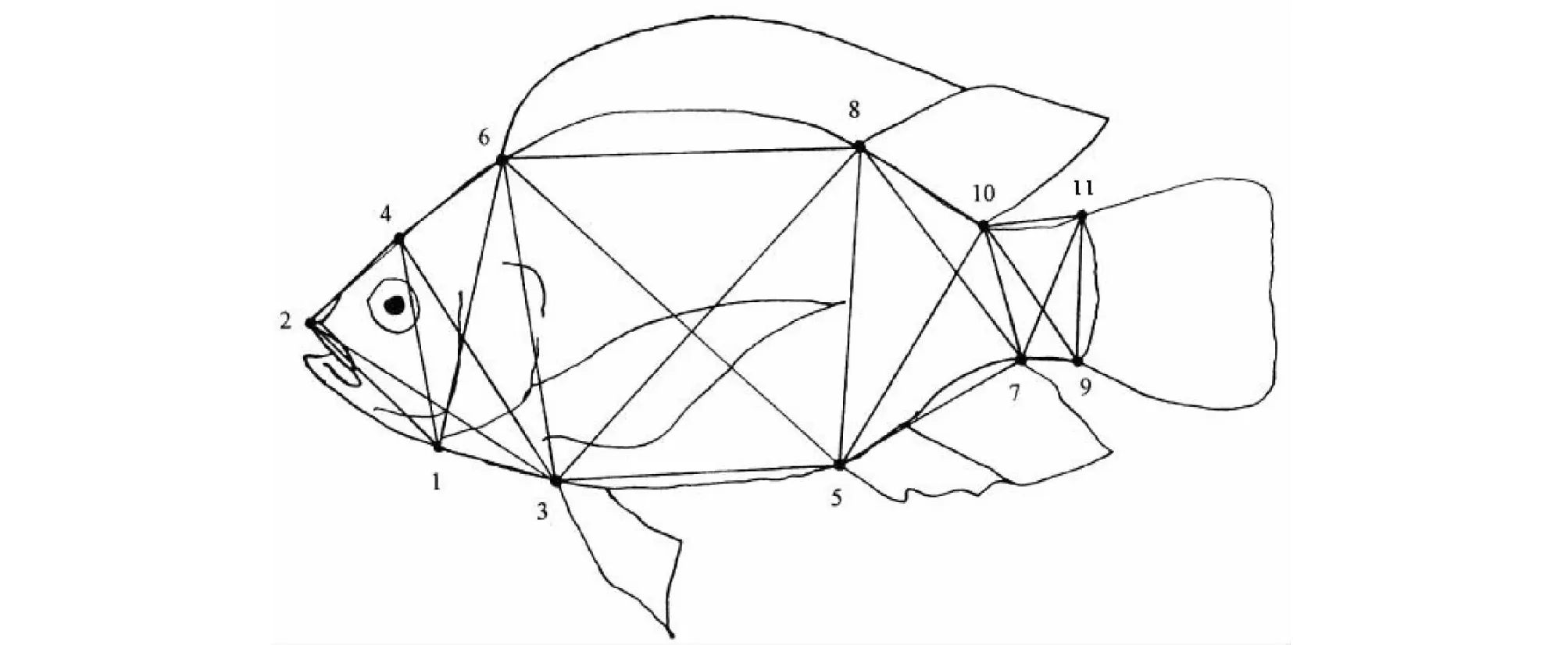
1:下颌骨最后端;2:吻前端;3:腹鳍起点;4:额部有鳞部最前缘;5:臀鳍起点;6:背鳍起点;7:臀鳍末端;8:背鳍鳍棘部末端;9:尾鳍腹部起点;10:背鳍末端;11:尾鳍背部起点1: Most posterior point of maxillary; 2: Tip of snout; 3: Origin of pelvic fin; 4: Most anterior of scales on skull; 5: Origin of anal fin; 6: Origin of dorsal fin; 7: Terminus of anal fin; 8: Terminus of spines in dorsal fin; 9: Ventral origin of caudal fin; 10: Terminus of dorsal fin; 11: Dorsal origin of caudal fin图2 罗非鱼框架测量图Fig.2 Drawing for measuring truss network of Tilapias
1.3.2 判别分析 采用逐步判别法从34项标准化性状中筛选出对区分7个地理群体红腹罗非鱼有显著贡献的性状,并根据性状相关系数构建各地理群体的判别函数。将形态性状数值分别代入7个判别函数中,所得最大值对应的地理群体即为该样本的判别分类[11],再按赵优等[14]的方法分别计算判别准确率和综合判别率。
判别准确率( %)=(某地理群体判别正确的尾数/该地理群体实际样本数)×100
综合判别率( %)=(所有地理群体判别正确的尾数/所有地理群体实际样本总数)×100
1.3.3 聚类分析 分别求出各地理群体红腹罗非鱼34项标准化性状的平均值,采用欧氏距离矩阵和最短距离法[12]对7个地理群体进行系统聚类分析,探讨各地理群体间红腹罗非鱼的形态距离。
1.3.4 体长—体质量回归分析及肥满度分析 采用Keys公式W=bLa对各地理群体的体长—体质量进行回归拟合分析,式中W为体质量(g),L为体长(cm)。根据幂指数a值判断红腹罗非鱼的野外生长速度,当a值在3.00附近时,表示鱼体为匀速生长,当a值在2.00或4.00附近时,则表示鱼体为异速生长[15]。
以K=W×100/L3计算鱼体肥满度并进行方差分析,比较各地理群体间肥满度的差异性[16],式中K为肥满度,W和L同上式。
2 结果与分析
2.1 各地理群体红腹罗非鱼形态性状的主成分分析
通过逐步剔除信息提取率较低的性状,选取红腹罗非鱼头长(X1)、眼径(X2)、躯干长(X3)、尾长(X4)、尾柄高(X5)、D1-4(X6)、D2-4(X7)、D3-5(X8)、D3-6(X9)、D4-6(X10)、D8-3(X11)、D8-5(X12)、D8-7(X13)、D8-10(X14)、D7-9(X15)、D9-10(X16)、D9-11(X17)和 D10-11(X18)共18项标准化性状进行主成分分析,最终共提取出6个主成分(PCA1~PCA6),贡献率分别为32.455 %、13.950 %、8.000 %、7.278 %、6.376 %和5.797 %,累计贡献率为73.855 %。其中,PCA1、PCA2和PCA3的贡献率最高,各标准化性状的得分系数如图3所示。
PCA1中得分系数较高的是主要反映鱼体头尾轴和躯干部特征的D8-3(X11)、D3-5(X8)、躯干长(X3)、D3-6(X9)、尾长(X4)、D8-10(X14)和头长(X1);PCA2中得分系数较高的是主要反映鱼体前侧尾部特征的X13(D8-7)、X14(D8-10)和X12(D8-5);PCA3中得分系数较高的有X18(D10-11)、X15(D7-9)、X9(D3-6)、X2(眼径)和X16(D9-10),主要反映鱼体尾柄以及部分头部特征。从以PCA1为横坐标、PCA2为纵坐标绘制的散点图(图4)可见,来宾群体和巴马群体重叠最多,天峨群体和东兰群体有部分重叠,都安群体则与周边各群体均有重叠。各地理群体总体上形态离散明显,与实际地理分布具有一定的相关性,PCA1坐标轴上体现的差异较PCA2坐标轴更为明显。

图3 PCA1、PCA2和PCA3得分系数Fig.3 Scoring coefficient of PCA1, PCA2 and PCA3
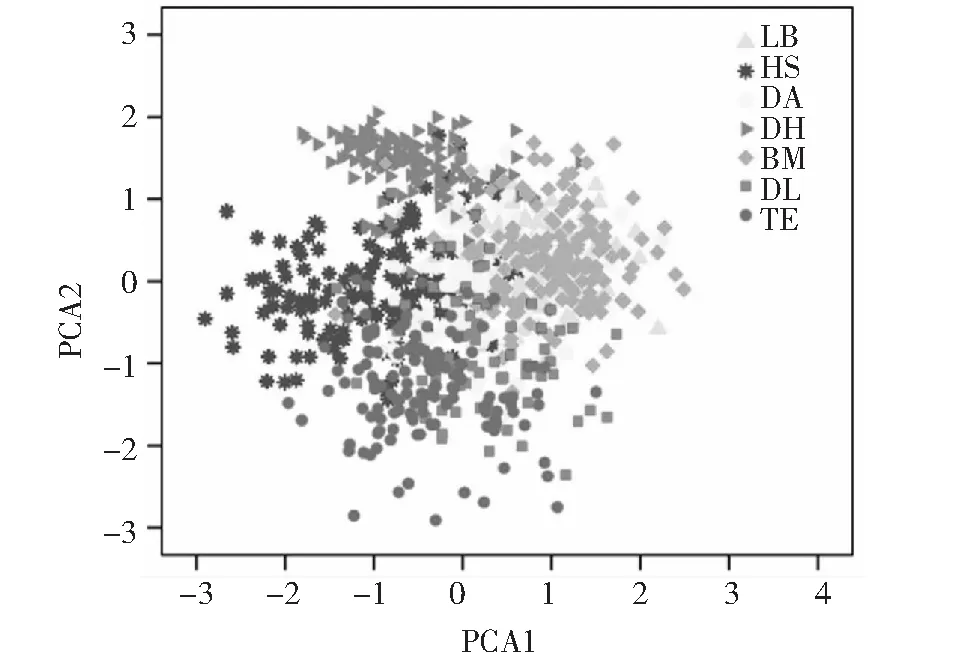
图4 各地理群体红腹罗非鱼在主成分PCA1和PCA2的分布Fig.4 Distribution of redbelly tilapia (Coptodon zillii) from each geographical population on PCA1 and PCA2
2.2 各地理群体红腹罗非鱼形态性状的判别分析
采用逐步判别分析法筛选出28项对区分7个地理群体的红腹罗非鱼有显著贡献的性状,各群体样本的性状相关系数见表2,依此建立各地理群体的判别函数,例如:Z天峨= -5128.380-369.818Y1+76.425Y2+……+175.505Y27+75.160Y28。对所有样本进行逐一判别,结果表明巴马群体和来宾群体的判别准确率分别为80.12 %和81.91 %,其它地理群体的判别准确率为91.78 %~98.52 %,最终综合判别率为91.29 %(表3)。
2.3 各地理群体红腹罗非鱼形态性状的聚类分析
经聚类分析,天峨、东兰、都安、合山和大化5个群体的红腹罗非鱼形态较为相似,而来宾群体和巴马群体的红腹罗非鱼与其它5个群体的相似性较低。其中,大化群体、合山群体和都安群体的形态距
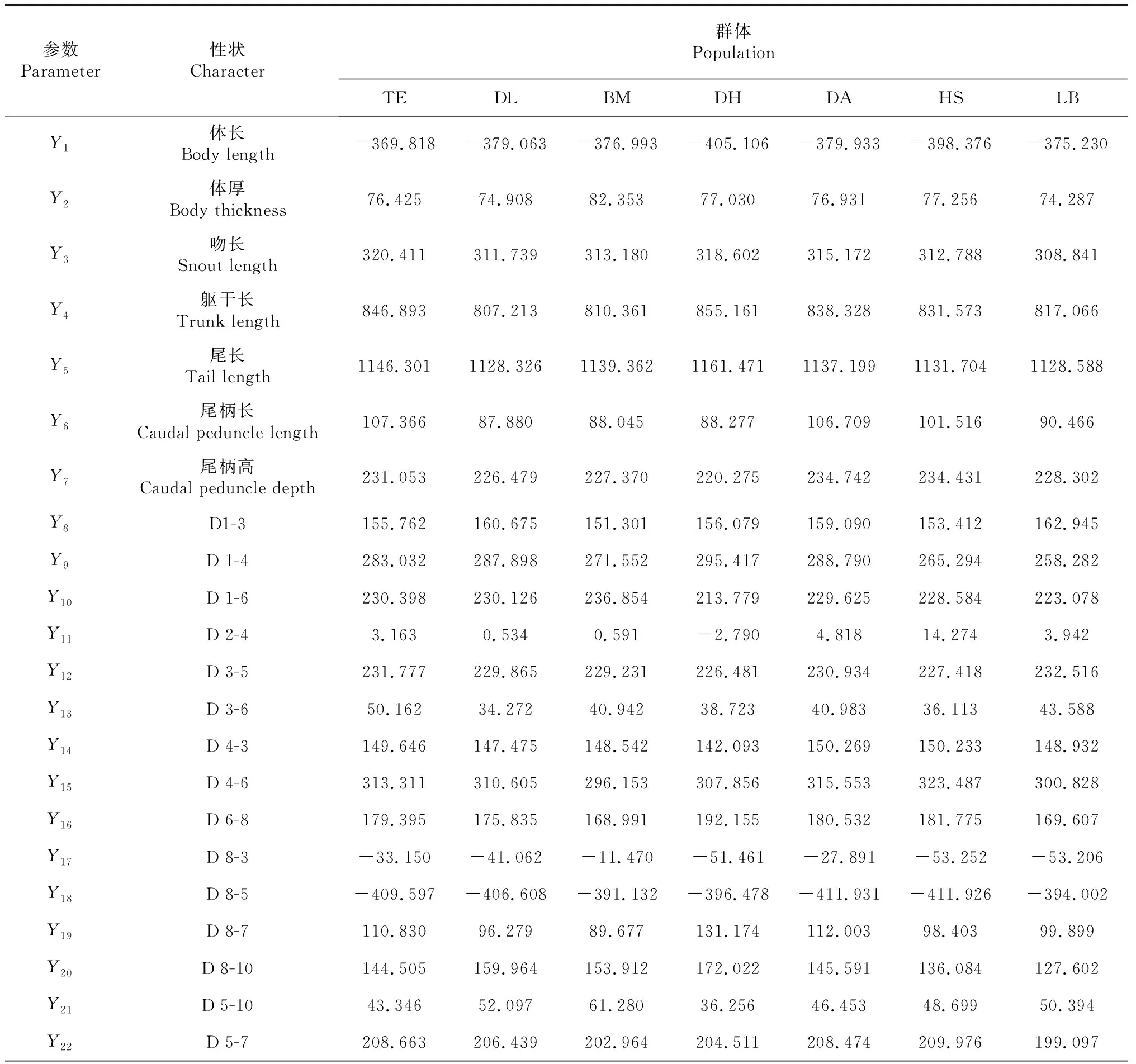
表2 各地理群体红腹罗非鱼样本的性状相关系数
续表2 Continued table 2

参数Parameter性状 Character群体 PopulationTEDLBMDHDAHSLBY23D 7-914.84119.88920.75315.84412.19510.97928.505Y24D 7-1068.45073.29165.89270.90678.91785.52766.576Y25D 7-11-22.540-14.600-14.296-20.125-26.082-22.752-11.962Y26D 9-10104.725111.520106.981105.399106.58799.164111.473Y27D 9-11175.505166.630165.954168.009175.137181.101170.955Y28D 10-1175.16074.05676.53770.98370.08266.54873.760-常量 Constant-5128.380-4955.400-4942.070-5087.870-5122.920-4970.420-4825.170
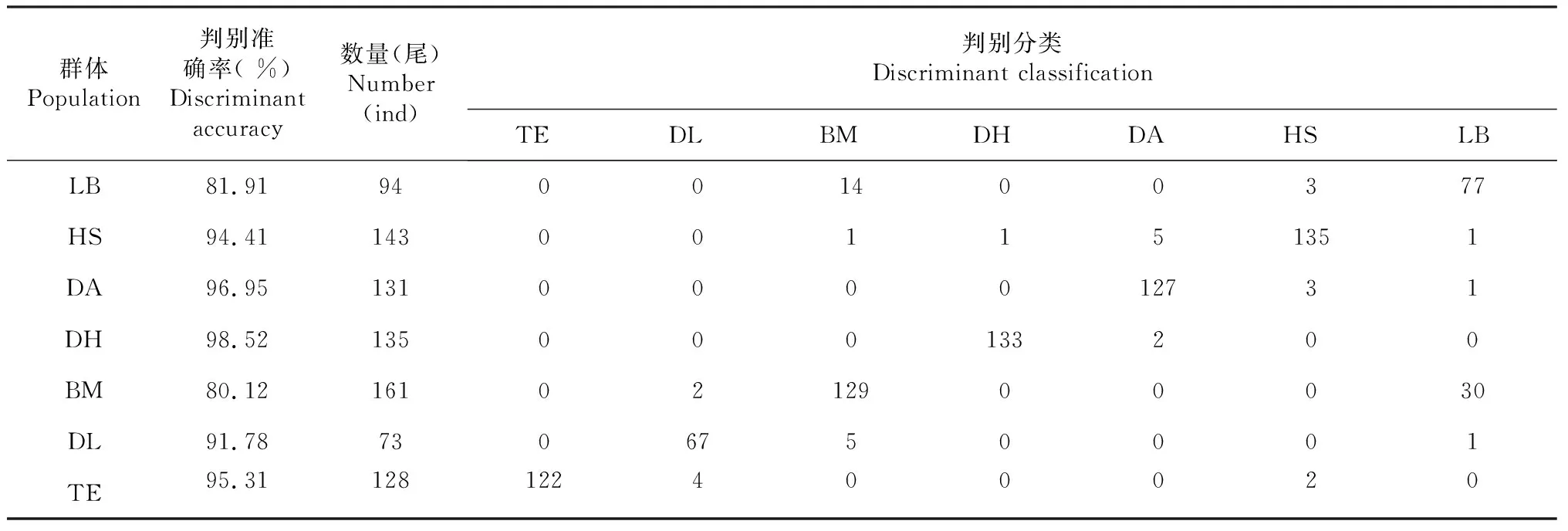
表3 红腹罗非鱼样本判别分类
离较近,先聚为一支,再与天峨群体和东兰群体聚类,最后与形态距离较远的来宾群体和巴马群体聚为一整支(图5)。
2.4 各地理群体红腹罗非鱼体长—体质量的相关性及肥满度
由红腹罗非鱼样本的体长—体质量回归分析及肥满度计算结果(表4)可知,各地理群体以及总体体长—体质量拟合方程的R2值为0.924~0.983,均能很好地拟合出样本体长与体质量的关系。此外,各拟合方程中幂指数a值均在3左右,表明红水河各地理群体红腹罗非鱼在野外环境下都近似等同于匀速生长。
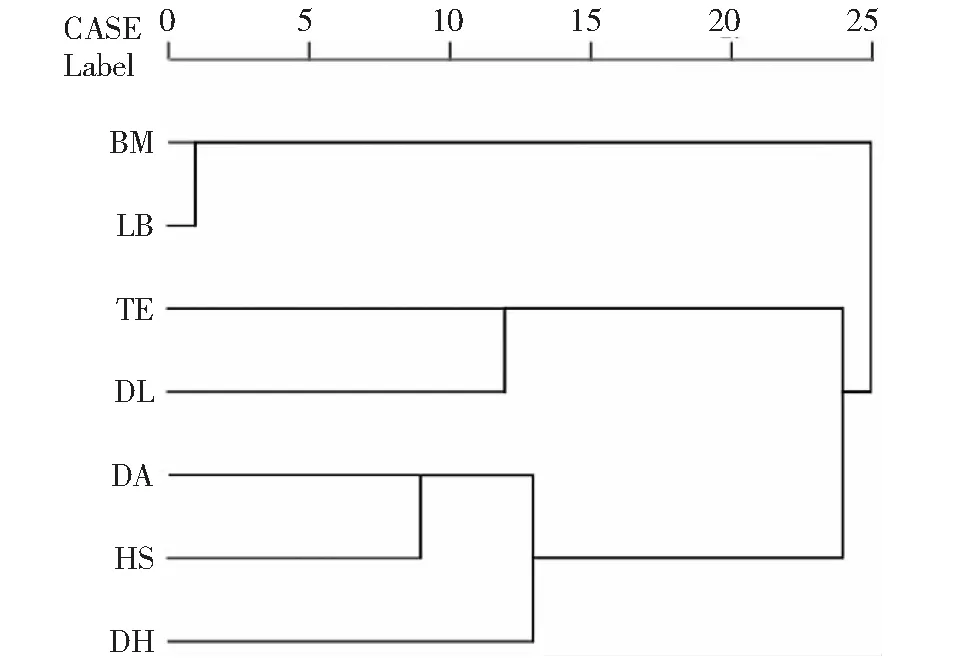
图5 红腹罗非鱼样本聚类分析结果Fig.5 Cluster analysis result of redbelly tilapia (Coptodon zillii)
表4中,肥满度基本在地理距离相近的2个群体间差异不显著(P>0.05),地理距离较远的群体间则往往差异显著(P<0.05)。7个地理群体中以东兰群体和天峨群体的平均肥满度最高,分别为4.58和4.52;其次是合山群体和来宾群体,二者均为4.39;巴马群体、大化群体和都安群体则相对较低,分别为4.17、4.10和4.04。
3 讨 论
3.1 红腹罗非鱼各地理群体的形态差异
在统计学中,主成分分析是一种可以将多种参数综合成少数因子从而能够直观体现群体变异来源的分析方法[12]。本研究通过主成分分析发现红腹罗非鱼各地理群体间的形态差异主要集中在头尾轴、躯干部和尾部性状上,与多数鱼类不同地理群体间的主要差异性状一致[14, 17-18]。一般情况下,鱼体头尾轴和躯干部性状决定了鱼的体型,有助于鱼类适应不同水文环境下的游泳阻力,而尾部是鱼类前进的推动器官,尾部性状的差异直接影响其游动速度。这些形态参数是区分红腹罗非鱼不同地理群体形态差异的关键要素。此外,逐步判别分析被认为是比主成分分析量化指标效果更佳的一种分析方法,通过构建判别函数不仅能对不同群体进行判别,还能据此推断群体间差异程度[14, 17]。本研究分别建立了广西红水河7个红腹罗非鱼地理群体的判别函数,综合判别率达91.29 %,其中来宾群体和巴马群体的部分个体存在误判,判别准确率均低于82.00 %,其它5个地理群体的判别准确率则较高(91.78 %~98.52 %),表明通过28项形态性状建立的判别函数能有效地区分不同地区样本,除来宾群体和巴马群体具有一定的相似性外,其余各地理群体间形态分化明显。进一步对不同地理群体的34项标准化性状进行聚类分析,也发现来宾群体和巴马群体虽地理间隔较远,形态相似性却较高,而天峨群体、东兰群体、大化群体、都安群体和合山群体表现出地理分布和形态距离间的一致性,与Poulet[19]对法兰西雅罗鱼(Leuciscusburdigalensis)的研究结果相似。可见,红腹罗非鱼虽同在红水河广西段的不同河段中,但不同地理群体间在形态上仍存在一定的差异。
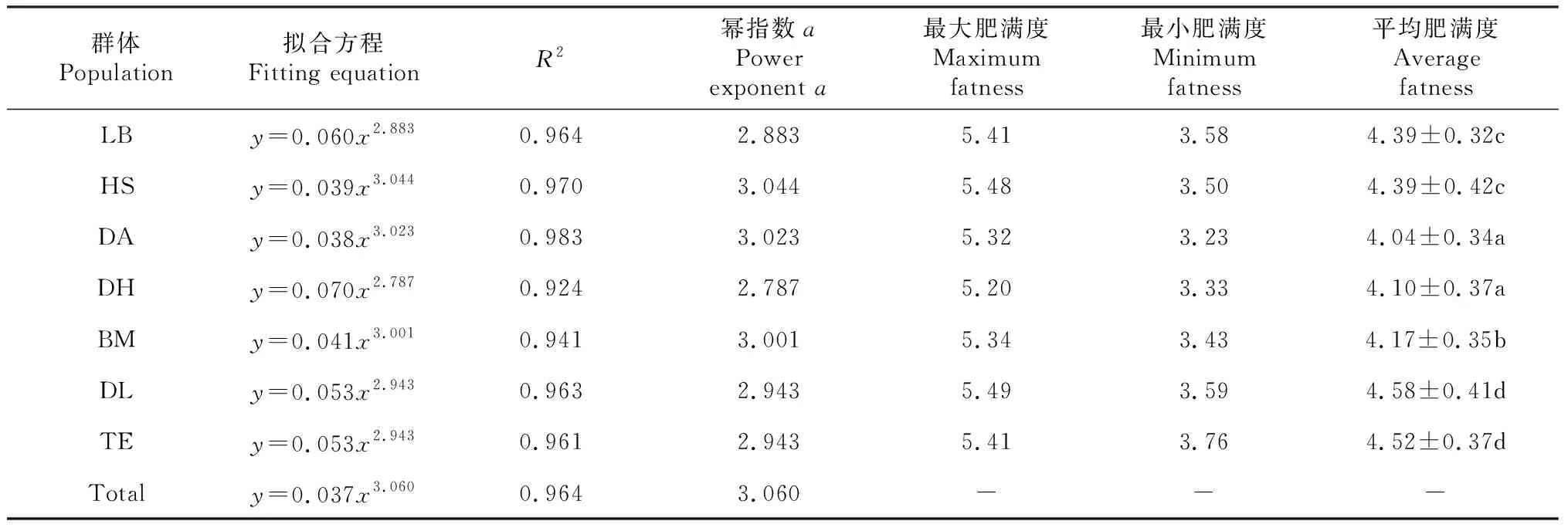
表4 各地理群体红腹罗非鱼体长—体质量的拟合方程及肥满度
注:最后一列数据后不同小写字母表示群体间差异显著(P<0.05)。
Note:The different lowercase letters after the data in last column indicate significant difference among populations (P<0.05).
根据以往的研究结果,鱼类形态易受地理隔离、局域生境变化等环境因素的影响,鱼类不同群体间可能在长期隔离和适应各自环境后出现显著的形态差异,并且和群体的地理分布有关[13, 17, 19]。红水河广西段属典型的喀斯特地形,地势高低起伏,水文环境复杂多变,水资源极为丰富且水流落差大,该河段已建立了龙滩、岩滩、大化、百龙滩、乐滩和桥巩等多个梯级水电站[20]。水电工程建设往往会导致水域的片段化和破碎化,影响不同水域鱼类群体间的基因交流,改变各鱼类群体的栖息环境[21-22]。2017年,非洲学者也发现尼日利亚境内3个水库中红腹罗非鱼的形态差异与处于地理阻隔条件下各水体的温度、盐度和水流速度等环境因子存在一定的关联性[23]。综合前人和本研究结果,广西红水河外来红腹罗非鱼的表型分化可能与红水河中水电工程大坝形成的地理阻隔以及各河段复杂各异的水文条件、营养状况等局域生境有关,但此推测仍需更多的相关性研究资料予以证实。
总体而言,上述统计分析方法从不同角度分析了广西红水河外来红腹罗非鱼的形态特征,它们之间不能相互替代,但却可以相互印证。3种分析方法所得结果基本一致,即天峨、东兰、大化、都安和合山5个地理群体的红腹罗非鱼形态差异明显,而来宾群体和巴马群体则具有特殊性,这两个地理位置较远的群体间具有较高的形态相似性而与其它群体间形态距离较远,推测可能与这两个群体引种来源相同而与其它群体存在差异有关。郑文佳等[24]根据云南伊洛瓦底江外来种麦穗鱼的形态学分析结果,认为不同地理群体形态差异较小可能与引种来源相同以及引入时间较短有关,这对分析本研究结果有参考意义。生物的表型是特定基因在外部环境中表达的结果[17],不同地理群体红腹罗非鱼的形态特征受遗传和环境因素的双重影响。本研究仅从形态学上对7个不同地理群体红腹罗非鱼的形态差异特点进行了初步探讨,后续仍需基于线粒体和核基因标记进行遗传学分析,以获得更多有价值的研究资料。
3.2 广西红水河红腹罗非鱼野外生存现状
由于水生生态系统本身的复杂性和不易观察性,使得鱼类的生态入侵尤为难以控制[25]。近期研究发现红腹罗非鱼已成功入侵伊朗部分水域和我国福建九龙江华安段及广东东江[2, 5, 26-27],表现出极强的环境适应能力和竞争力。广西水系发达,红水河拥有布柳河、盘阳河、刁江和清水河等众多支流,为红腹罗非鱼的成功入侵提供了良好条件和广阔的生存空间,本研究在样品采集过程中也观察到红腹罗非鱼已扩散至广西红水河各自然水域,且数量众多。
体长和体质量是鱼类重要的生物学指标,通过体长—体质量回归分析和肥满度计算能够客观了解鱼类的生存状态。Omotayo等[28]对尼日利亚水库内的红腹罗非鱼和尼罗罗非鱼进行了体长—体质量回归分析,发现2种鱼的幂指数a值均显著小于3.00,鱼体呈现负异速生长,并认为这可能与2种鱼相互竞争食物及环境因子有关。而本研究结果显示,红腹罗非鱼各地理群体体长—体质量幂指数a值均在3.00左右,说明鱼体在广西红水河中生长速度均匀,生存状况良好。Nehemia等[29]探究了淡水和海水饲养条件下红腹罗非鱼肥满度的变化,结果表明当鱼体在淡水条件下肥满度大于1.00时,可视为生长状况良好。本研究所测各地区样本平均肥满度为4.04~4.58,与Anene[30]计算的尼日利亚人工湖中红腹罗非鱼平均肥满度(4.30)相类似,二者都远大于1.00,表明各地理群体的鱼体均较肥满。
以上分析说明,广西红水河外来红腹罗非鱼野外生存状态良好,具有较强的适应性和竞争力,结合潘勇等[25]的调查研究,红腹罗非鱼在成功入侵新环境后可能会通过与土著鱼类竞争食物和生存空间,甚至吞食鱼卵和幼苗而对其造成危害。此外,已有研究发现红腹罗非鱼能够携带线虫、吸虫、蠕虫类寄生虫以及多种病原微生物[31-32]。因此,对于已经入侵到广西红水河的红腹罗非鱼,既要及时开展危害风险评估,建立相关监测和预警机制,还应根据其生理特性(如食性、环境因子耐受性、繁殖习性、耐寄生虫和病菌感染等)进行综合治理,防止进一步扩散。
4 结 论
广西红水河外来红腹罗非鱼在不同地理条件下已产生了较明显的适应性形态分化,野外生存状态良好,竞争力强,对其应及时开展危害风险评估及相关监测。
