老龄大鼠听功能下降与线粒体DNA4834bp缺失突变的关系
2018-11-22黄海林刘俊
黄海林 刘俊⋆
老年性耳聋是老年人中发生率较高的慢性疾病,是衰老在听觉器官的表现。研究表明,衰老与线粒体功能紊乱密切相关,而线粒体功能紊乱是线粒体DNA突变引起的。Bai等[1]报道mtDNA4977缺失与老年性耳聋存在可能相关性。Fischel-Ghodsian等[2]检测5例老年性耳聋患者膜迷路和螺旋神经节中mtDNA编码的细胞色素氧化酶Ⅱ基因,发现老年性耳聋患者mtDNA突变率高于老年对照组,但突变发生的部位和数量均存在较大个体差异。常见的线粒体DNA缺失突变片段有 13162bp,10422bp,7663bp,7436bp,4989bp,4977bp,其中人mtDNA4977bp(对应大鼠4834bp)是mtDNA中最为常见突变类型[3]。2015年10月至2016年3月作者通过实验研究,探讨老年性聋发生的可能机制。
1 材料与方法
1.1 实验动物 SPF级SD大鼠20只,其中24个月龄SD大鼠10只,平均(647.83±10.35)g;3个月龄SD大鼠10只,平均(210.78±5.62)g,所有动物均无噪声暴露史及其它药物使用史,且无中耳炎,动物由华中科技大学同济医学院提供。
1.2 方法 (1)分组:20只SD大鼠经听性脑干反应(ABR)检测,排除中耳炎后按年龄分2组,各10只,24个月龄为观察组;3个月龄为对照组。(2)检测大鼠听性脑干反应(ABR)反应阈:分别对对照组和观察组大鼠进行ABR反应阈测定(测定方法参照孔维佳建立方法[4])。测试前两组大鼠用10%水合氯醛按照0.4ml/kg腹腔内注射麻醉,使用GSI Audera脑干诱发电位仪测试。(3)蜗神经核及内耳组织mtDNA提取:两组动物ABR测试结束后立即麻醉断头处死,首先提取脑蜗神经核:于中线处剪开颅骨,将脑组织轻轻从颅骨中取出,根据蜗神经核[5]定位提取脑蜗神经核,置入匀浆器中,加入2ml匀浆液(pH7.4,0.01mol/L Tris-HCI,0.1mmol/L,0.01mol/L蔗糖)匀浆至无明显组织块存在(冰浴操作)。匀浆液4℃离心1000g l0min,取上清液0℃离心12000g 15min,弃上清液,沉淀物即为脑组织蜗神经核线粒体。然后提取内耳组织:提取双侧听泡,置于4℃预冷的充氧无钙镁细胞外液中(0.1mol/L PBS,pH 7.4),体视显微镜下剥去骨性蜗壳,解剖出耳蜗Corti器,血管纹,螺旋神经节等内耳组织。蜗神经核和内耳组织线粒体DNA提取方法:各组织置入匀浆管,加入RIPA裂解液,再加入蛋白酶 K,至终浓度10mg/ml,摇匀混合,55℃水浴过夜消化。最后用酚-氯仿-异戊醇抽提取线粒体DNA,所得 DNA 溶于 50μl TE 溶液,-20℃保存备用。取2μl DNA样本溶液稀释至50μl,用紫外线核酸检测仪器检测A260/A280纯度值并测定其浓度大小。(4)线粒体DNA缺失检测分析:根据大鼠线粒体基因组全序列(Genbank序列号X 14848),并遵循引物设计原则,引物由上海生工生物工程公司合成:P1:5'-GCGAAGCTTAGAGCGTTAAC-3',P2:5'-AGT GAG ATA AGG AAG CCT GC-3',引物合成后用灭菌超纯水稀释成10μmol/L 的工作液,-20℃冻存。配制25μl PCR反应体系:Taq DNA 聚合酶(2.5U)0.5μl,引物(10μM) 各 1μl,dNTPs 2μl,PCR buffer2.5μl。PCR扩增反应条件为:95℃预变性5min,94℃变性30s,56℃退火30s,72℃延伸45s,反应35个循环;末次72℃延伸7min。引物P1、P2如可以扩增597bp片段,则提示样本中有mtDNA4834bp缺失。如样本中无4834bp缺失突变,应在现实反应条件下无法合成近5000bp片段,故无法扩增出597bp PCR产物。(5)PCR产物鉴定与分析:各组PCR扩增产物经1.0%琼脂糖凝胶电泳,溴化乙啶染色,电泳缓冲液1 X TAE,电压100V,用凝胶成像仪扫描成像并保存结果。在紫外灯下确定597bp目的基因条带位置,将凝胶与产物一起切出,用DNA凝胶回收试剂盒(AXYGEN APGX-50G)分离纯化。加样后将离心管置37℃水浴1h,酶切产物经2. 5%琼脂糖凝胶电泳,溴化乙啶染色,凝胶成像仪扫描成像并保存结果。
1.5 统计学分析 采用SPSS18.0统计软件。计量资料以()表示,用t检验,P<0.05为差异有统计学意义。
2 结果
2.1 ABR反应阈 对照组大鼠ABR平均听阈(19.50±3.69)dB, 观 察 组 大 鼠 ABR平 均 听 阈(46.00±3.94)dB,两组差异有统计学意义(P<0.05)。
2.2 mtDNA4834bp缺失突变电泳结果 对照组大鼠脑组织蜗神经核样本(10例)及内耳组织样本(10只)均未检测出597bp(提示4834bp缺失)扩增条带(见图1),观察组大鼠蜗神经核样本检出率为100%;内耳组织样本检出率90.0%。见图2。
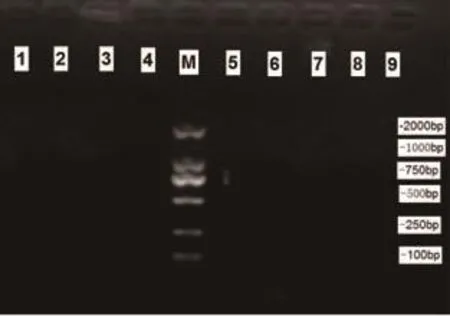
图1 对照组各组织标本PCR结果
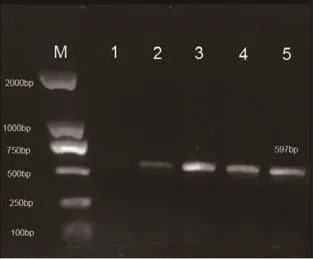
图2-1 观察组蜗神经核组织mtDNA PCR结果
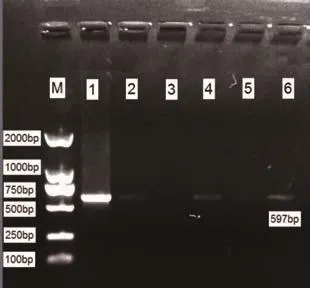
图2-2 观察组内耳组织mtDNA PCR结果
2.3 PCR酶切电泳 根据大鼠线粒体基因组全序列(Genbank序列号X 14848),根据DNASTAR软件选择限制性内切酶(HphⅠ)酶切,酶切位点见图3,结果证实酶切产物来自mtDNA4834bp特异型缺失扩增片段,见图4。

图3 HphⅠ限制性内切酶酶切位点
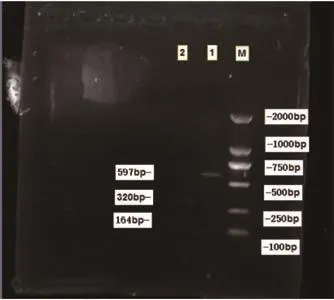
图4 PCR产物酶切电泳图
3 讨论
老年性耳聋发生机制目前尚不明确,大多学者认为其由多种危险因子的长期累积共同作用形成,其中最主要因素为衰老。根据Harman[6]提出衰老的自由基学说,认为衰老是由自由基引起的组织随机毒害的结果,自由基损害作用的机制主要涉及DNA的损伤,尤其是mtDNA的氧化损伤,其持续产生于线粒体电子传递链的活性氧簇(ROS)是衰老过程中线粒体DNA氧化损伤产生的主要原因。由于mtDNA裸露于产生高氧自由基的线粒体内膜下,本身无组蛋白的保护又缺乏损伤修复系统,其突变率较核DNA高10~20倍[7],极易受氧自由基的侵袭发生氧化修饰和mtDNA突变,由缺陷的mtDNA编码的呼吸酶影响电子转运功能,增加电子泄露,不断产生的ROS又反过来继续氧化损害线粒体功能,导致不同组织细胞程度不一的mtDNA累积突变的蓄积,即“老化的线粒体理论[8]”。衰老过程中产生的mtDNA缺失突变型mtDNA比完整的野生型mtDNA长度小[9],复制速度快,其增殖优势导致缺失突变型mtDNA分子比例随年龄增长不断累积增加。当细胞内突变mtDNA积累达到氧化磷酸化(OXPHOS)所生成的能量低于维持正常细胞功能阈值时,组织器官开始退化,出现临床症状,引起老年退化性疾病,在听觉器官表现为听力逐渐下降。本实验中对照组大鼠ABR平均听阈为(19.50±3.69)dB peSPL,观察组大鼠ABR平均听阈(46.00±3.94)dB peSPL,两组间差异有统计学意义(P<0.05)。
老年性聋是机体衰老过程在听觉器官的表现。研究发现线粒体DNA突变与老年性耳聋的发生关系密切。人类mt DNA 在8483~13459bp之间存在着13bp的重复序列,在氧化应激刺激下,易发生断裂,在修复时由于两端重复序列,产生DNA两条链下游环,该碱基环富含鸟嘌呤碱基,对自由基及其敏感,一旦受到ROS攻击,该环降解即容易导致“误认”发生了4977bp片段缺失[10],此机制被称为线粒体重组滑行错配学说。此缺失在大鼠,mtDNA8103-12937bp之间存在16bp直接重复序列,故大鼠也存在类似缺失突变(mtDNA4834bp)的常见缺失(CD)。突变不仅存在衰老组织中(心肌、肝、脑、肺、肾等),还可存在于线粒体脑肌病、进行性眼外肌麻痹和Kearns-Sayre综合征等疾病中,且随着年龄增加在组织中逐渐积累[11]。本实验PCR扩增出597bp片段(提示mtDNA4834bp缺失),该缺失突变编码包括ND3,ND4,NDS,COIII,ATPaseB,ATPase6 等[12]基 因,影响呼吸链有关亚单位蛋白的合成,从而形成有缺陷的呼吸链,ATP产生不足导致蜗内离子失平衡,内环境紊乱,最终导致内耳毛细胞非特异性损害,引起听力下降[13]。本实验在大鼠蜗神经核和内耳组织中均发现了mt4834bp缺失,表明该常见缺失普遍存在于衰老组织。因此以后可以通过减少和预防mtDNA缺失突变的发生,以缓解和预防老年性耳聋发生发展,从而为治疗老年性耳聋提供新突破。但老年性聋不一定均有mtDNA4834bp片段缺失,本实验中观察组有1只未检测到该缺失突变,表明除了基因背景以外,还有其他环境易感因素(如药物,噪音等)在老年性聋的发病过程中起重要作用,具体机制有待于进一步的研究。
