干旱胁迫下胡杨实生幼苗氮素吸收分配与利用
2018-11-15马晓东钟小莉
马晓东,钟小莉,桑 钰
新疆师范大学生命科学学院,新疆特殊环境物种保护与调控生物学实验室,干旱区植物逆境生物学重点实验室, 乌鲁木齐 830054
土壤营养元素直接影响植物的生长发育和其在植物体内各部位间的分布特征,从而影响植物分布格局和种群演替[1]。氮素是土壤营养元素的重要组分,是构成植物体内蛋白质及核酸的重要元素,氮素水平直接或间接影响植物的光合生理和生长状况,尤其是土壤中的有效氮[2- 3]。植物对氮素的吸收利用可以带动整个生态系统的水分与氮素的应答效应[4]。塔里木河下游区域是典型的极端干旱气候区,该区土壤营养元素具有“缺氮少磷富钾”的特点[5]。在这样的干旱荒漠生态系统中,水和氮素成为影响植被组成、分布及多样性最主要的限制性因子[6]。因此,干旱胁迫和氮素匮乏极大限制了荒漠河岸林生态系统的更新演替。
胡杨(PopuluseuphraticaOliv)是塔里木河流域荒漠河岸林典型中生植被类型,作为重要的建群种和优势种能隔断沙漠,遏制沙化,对维护该区生态平衡有至关重要的作用[7]。研究表明,氮素是胡杨生态系统矿质营养的重要限制因素[8]。植物在受到干旱和养分等环境因子的限制时,通过调整其自身不同器官物质的分配等生长策略来维持正常的生存[9]。目前,针对塔里木河下游胡杨的研究多集中于胡杨的生态分布[10]、生理特性[11]及抗逆性等[12-13]方面,而有关胡杨实生苗应对干旱胁迫和氮素匮乏的生长策略研究较少。氮素的供应能有效调整作物不同器官的干物质分配[14],胡杨实生苗吸收有限的氮素后,各器官之间对养分的运移分配及养分利用效率的变化规律对幼苗生长影响很大。很多的研究都表明氮素能促进植株不同物候期的器官形态建成[11- 14],那么提出假设胡杨幼苗受干旱胁迫等环境因子限制时,利用吸收适量的氮素对干旱胁迫具有补偿效应,来调整自身物质分配,提高氮素吸收利用效率,促进其早期的形态建成。因此,我们采用添加标记同位素的方法,定量分析植物器官中同位素的空间分布,揭示塔里木河下游胡杨实生苗的生存机制或特征,可为胡杨林的保护和培育管理,以及维护荒漠河岸林系统稳定性提供理论依据。
1 材料与方法
1.1 实验材料
2016年5月中旬在塔里木河下游采集的胡杨成熟蒴果,挑选饱满、大小均匀的种子进行实验。播种至直径30 cm,高30 cm的花盆中。实验用土取自塔里木河下游胡杨群落分布区沙土,过2 mm筛去除杂质并进行高温灭菌。实验沙土的田间持水量为22.75%,全磷含量平均为0.467 g/kg,全钾含量平均为21 g/kg,全氮含量平均为0.33 g/kg,有效氮平均含量为12 mg/kg,有机碳含量约为2493.6 kg/hm2(0—60 cm土层),pH约为8.4。
1.2 实验设计
本研究采用温室内盆栽实验,设置4个水分梯度,土壤相对含水量分别为严重干旱20%—25%(D1)、中度干旱40%—45%(D2)、轻度干旱60%—65%(D3)、正常水分80%—85%(D4),于每日晚八点,采用整盆称重法控制土壤含水量,根据前一天晚上与当天晚上两次称重重量差及设定的土壤含水量来计算补水量,用塑料烧杯来补充自然蒸发的水分,同时结合WET- 2便携式水分快速测定仪测量土壤含水量以便更加准确的控制土壤含水量在各预定胁迫水平;并设置3个氮素梯度,氮肥采用NH4NO3为氮源,施氮处理设对照0 g/盆(N0)、3 g/盆(N1)、6 g/盆(N2)3个水平。同时,标记氮素选用以同位素标记的15NH415NO3为标记氮,标记量为20 mg15N /盆,采用微区注射法进行施肥,施加方式设定三层10孔(外层4孔,中层5孔,中心1孔),以保证其在土壤中分布均匀,把NH4NO3和15NH415NO3配置为硝酸铵溶液,使用注射器将氮肥溶液均匀的注射到土壤中,对照组添加相同量的蒸馏水作为对照。
2016年5月20日播种,待齐苗后间苗,每盆保留1株健壮幼苗,育苗期间进行常规水分管理,保证幼苗正常生长。40 d后进行氮素添加,每个处理4个重复,按实验要求各孔位添加NH4NO3水溶液8 mL,对照组添加等量的蒸馏水,氮素添加后3 d进行干旱胁迫实验。处理75 d后取样,按照根、茎、叶三部分进行取样,105℃杀青30 min,后置于80℃烘箱中烘干至恒重。
1.3 分析方法
(1)气象因素测定
采用2900ET小型自动气象站测定大气温度、相对湿度、太阳辐射等参数,并进行连续监测。
(2)生长量测定
采用称量法,将完整的植株挖出,按照根、茎、叶三部分进行取样,清水冲洗干净,再用去离子水润洗两遍,105℃杀青30 min,后置于80℃烘箱中烘干至恒重,然后用精度为千分之一的电子称测生物量。并应用WinRHZIO根系图像分析软件(Regent Instruments,Qué-bec,Canada)对根系长度、表面积等指标进行测量,按根系直径分为粗根(d>2.0 mm)、细根(0.5 mm≤d≤2.0 mm)、极细根(d<0.5 mm)分别记录根长、根表面积。
(3)叶绿素荧光参数
随机选取植株顶部完全展开的叶片(第4,5片),在晴朗天气下,于 2016年9月16日,采用便携式调制叶绿素荧光仪(Junior-Pam),从9:00—18:00,每隔3 h测定光适应下最大荧光值Fm′、初始荧光Fo′和实时荧光值F,每个处理重复6次。并于测量当日凌晨4:00点测量暗适应整夜的初始荧光Fo、最大荧光Fm。通过计算分析得到暗适应下的最大光量子产量(Fv/Fm)=(Fm-Fo)/Fm。
(4)光合色素含量
光合色素含量测定采用乙醇-丙酮混合液提取,在每个处理中随机摘取若干健康的胡杨幼苗叶片,将0.1 g新鲜叶片放入试管中,向试管中加入15 mL 80%丙酮和无水乙醇混合提取液(体积比为1∶1),每个处理共有6个重复,标好记号置于避光的条件下浸提48 h,待剪碎的同化枝变白,溶液呈绿色后摇匀,用紫外可见分光光度计分别在波长663、645、470 nm处测定吸光度(D)值,计算叶绿素a、叶绿素b含量及类胡萝卜素含量。
(5)叶片相对含水量
采用饱和称重法测定叶片相对含水量,取新鲜叶片,浸入蒸馏水中36 h,使叶片吸水成饱和状态,称取叶片饱和鲜重,再放入80℃干燥箱烘至恒重,称取干重,每个处理4个重复。叶片相对含水量=(叶片鲜重-叶片干重)/(叶片饱和鲜重-叶片干重)。
(6)15N丰度的测定
将用于测定的烘干样品器官分别进行粉碎,过0.15 mm筛,连续用四分法取出测定的样品量,并分装标记,用于测定植株含氮量和15N丰度。利用稳定同位素比例质谱仪,在中国植物研究所使用美国THERMO Finnigan 公司,型号为DELTAplusXP同位素比例质谱仪测定。计算公式如下:
Ndff(%)=[样品中15N的丰度%-自然丰度(0.365%)]/[肥料中15N的丰度%-自然丰度(0.365%)]×100;
分配率 = 幼苗各部分从15N吸收的氮量(mg)/15N吸收的总氮量(mg)×100;
利用率 = [Ndff×器官全氮量(g)]/施肥量(g) ×100;
总氮量(g) = 干物质量(g) × 全氮百分含量N%;
15N 吸收量(mg) = 总氮量(g)×Ndff%×1000。
1.4 数据处理
数据采用SPSS 19.0进行单因素方差分析(one-way ANOVA)、双因素方差分析(two-way ANOVA)及LSD多重比较检验相应数据的差异显著性(显著性水平设为0.05),使用Excel软件进行数据处理和制图各数据由平均值±标准误(SE)表示。
2 结果与分析
2.1 干旱胁迫下氮素对幼苗生理指标的影响
在D1条件下土壤严重缺水,幼苗的各项指标较低(图1)。在D2和D3时,叶片的相对含水量显著增加(P<0.05),此时氮素对其无显著影响;但在D1时,氮素显著提高叶片的相对含水量,且施氮量为N1时促进作用更大。说明适量的氮素能在较严重的干旱胁迫下显著提高叶片组织的含水量(图1)。干旱胁迫的加剧会使幼苗的最大光量子产量Fv/Fm明显降低。D1时,Fv/Fm明显低于其他处理,且差异显著(P<0.05)。但施氮能提高Fv/Fm,且在D3N1时达到显著水平(图1)。在D1、D4时光合色素含量较低,表现出其含量累积受抑制的现象,但D2、D3时氮素能显著增加两种色素含量(P<0.05),且D2N1时叶绿素含量最高达24.72 mg/L,D3N1时类胡萝卜素含量最高达到4.20 mg/L。同时,氮素N1较N2更有利于幼苗叶片的光合色素含量的增加(图1)。

图1 干旱胁迫下氮素对胡杨幼苗生长指标的影响Fig.1 The effect of nitrogen levels on growth parameter of P. euphratica seedlings under different drought stress①柱上不同字母表示处理间差异显著(P<0.05)。其中,D1:相对含水量为20%—25%,D2:相对含水量为40%—45%,D3:相对含水量为60%—65%,D4:相对含水量为80%—85%
2.2 干旱胁迫下氮素对幼苗根系形态的影响
幼苗根系的形态建成是实现长期生存和生长的关键。干旱胁迫减轻时,施氮能显著增加幼苗生物量,D3N1较D1N1幼苗生物量增加了89.5%;水分充足时D4,幼苗生物量反而有所降低。不同的水分和养分条件直接影响胡杨幼苗根系的可塑性。土壤水分对根冠比影响极显著(P<0.01),随土壤水分增加呈先升后降的趋势,D2时根冠比最大,D1时根冠比最小,表明严重干旱胁迫极大的限制根系扩展。相同干旱条件下,氮素对根冠比影响不明显,但对根长和根表面积影响较大。D1严重干旱胁迫时,幼苗根长和根表面积值很低,添加氮素使之进一步降低。随土壤水分状况的增加,根系形态明显改变。在D3时,根系长度和表面积达到最高,同时添加氮素为N1时,根系长度增加,加速了根系向纵深生长,根表面积也增大,有助于增强根的吸收能力。D3N1条件下,细根(0.5 mm

图2 旱胁迫下氮素对胡杨幼苗生物量及根系形态的影响Fig.2 The effect of nitrogen levels on biomass and root morphology of P. euphratica seedlings under different drought stress①柱上不同字母表示处理间差异显著(P <0.05); ②图B、图C、图D中不同小写字母表示相同干旱胁迫条件下,不同氮素间的差异显著(P<0.05);不同大写字母表示相同氮素条件下,不同干旱胁迫间差异显著(P <0.05)
此外,方差分析表明(表1),土壤含水量对胡杨幼苗生长的各指标均有极显著影响(P<0.01),氮素显著影响植株生长(P<0.05),同时两者的交互效应对幼苗光合参数和总生物量具有极显著的影响(P<0.01),说明土壤水分和氮素是影响胡杨幼苗早期生长的两个重要因素。

表1 数据显著性水平的方差分析
*显著P<0.05; **极显著P<0.01; NS显著性P>0.05
2.3 干旱胁迫下胡杨幼苗各部分的Ndff

图3 干旱胁迫下氮素对胡杨幼苗Ndff的影响 Fig.3 The effect of nitrogen on the Ndff of P. euphratica seedlings under different drought stress
植株各部分的Ndff指各部分从肥料中吸收分配到的15N含量对该部分全氮含量的贡献率,反映植株各部分对15N的吸收征调能力。胡杨幼苗各部分对15N的吸收征调能力(Ndff)对土壤含水量的变化十分敏感。随干旱胁迫减弱,幼苗各部分Ndff呈增加趋势,表明干旱胁迫会阻碍氮素的吸收。同时,相比于N2,N1条件下幼苗各部分Ndff显著较高,说明幼苗在较低或适量的氮素供给下有更好的吸收征调能力(图3)。
幼苗在生长的不同时期(0—75 d)各部分的Ndff表现出一定的差异,D1和D2处理下,幼苗的Ndff呈降低趋势,在75 d时值最低。氮素为N1时,D3和D4处理下的幼苗Ndff在50—75 d时呈增加趋势,75 d时均达到最高;而N2处理时,D3处理下幼苗Ndff在生长后期反而降低(图4)。

图4 不同时期干旱胁迫处理下氮素对胡杨幼苗Ndff的影响Fig.4 The effect of nitrogen on the Ndff of P. euphratica seedlings in different growth phase under different drought stress
幼苗地上和地下部分比较,25 d时,地上部分的Ndff显著高于地下部分(P<0.05),表明此时地上部分对15N吸收征调能力强(图5);50 d时,地下部分与地上部分的Ndff差异不显著,仅在D1时地下部分的Ndff显著高于地上部分(图5);75 d时,地下部分的Ndff显著高于地上部分,表明此时吸收的氮素主要用于根系的形态建成(图5)。
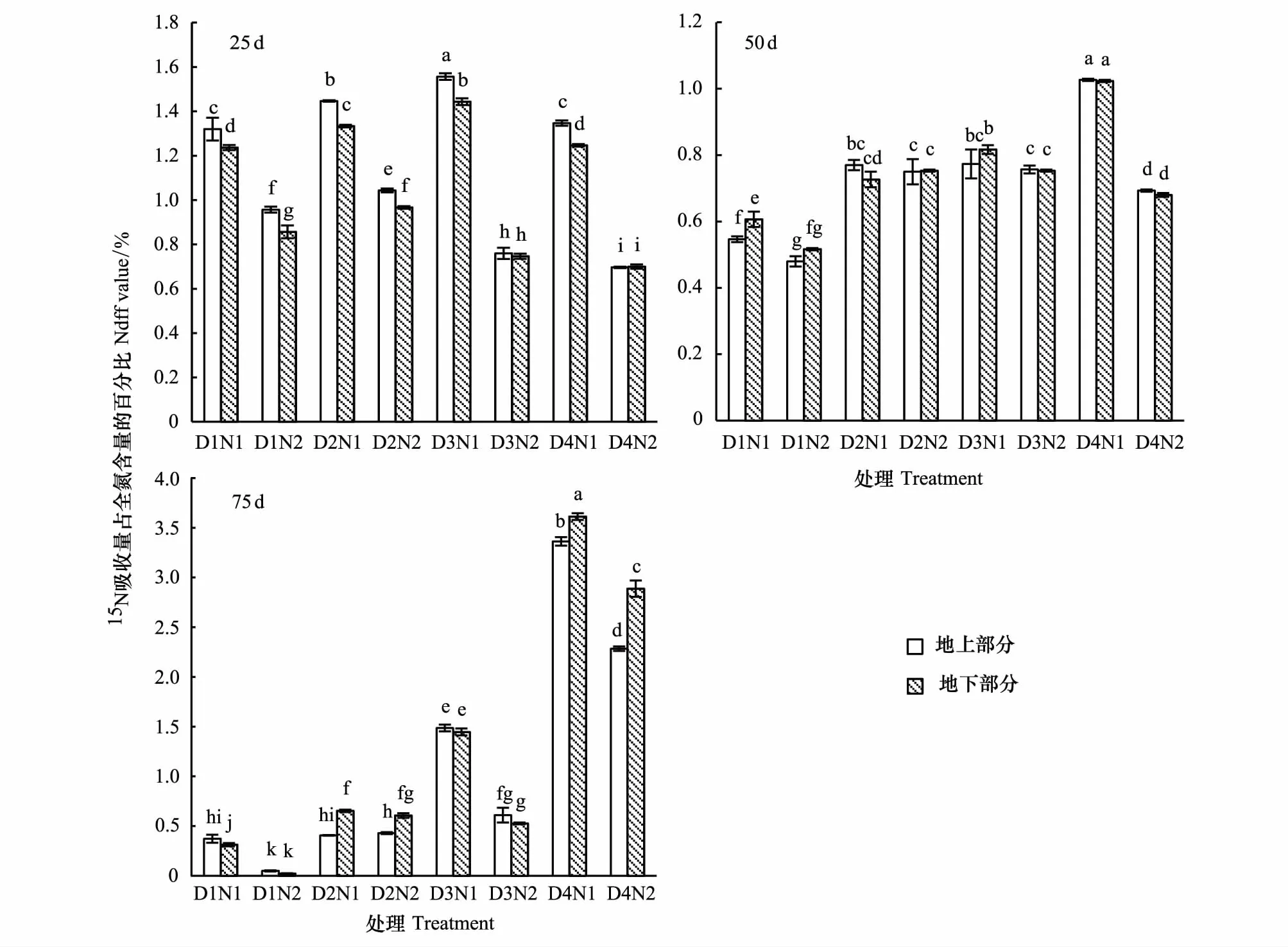
图5 不同时期各处理对胡杨幼苗地上和地下部分Ndff的影响Fig.5 The effect of Ndff on aboveground and underground part of P. euphratica seedlings in different growth phase under different treatments柱上不同字母表示处理间差异显著(P <0.05)
2.4 干旱胁迫下胡杨幼苗各部分的氮素分配率
不同处理条件下胡杨幼苗生长的不同时期,其15N的分配率差异显著(P<0.05)。幼苗处理25d后,地上部分15N的分配率高于地下部分,且在严重干旱胁迫D1时,幼苗地上部分的分配率最高;幼苗处理50 d后,幼苗根系15N的分配率随干旱胁迫加剧呈上升趋势,而地上部分15N的分配率则呈降低趋势。幼苗处理75 d后,根系15N的分配率在不同水分条件下地下部分均显著高于地上部分,这表明幼苗吸收的氮素在生长后期大部分分配到根系中贮藏。整个实验处理阶段,幼苗地上部分的分配率总体呈降低趋势,而地下部分的氮素分配率则有小幅的增加(图6)。
氮素在胡杨幼苗各器官之间的迁移规律及在植株内的分布情况差异显著(表2,P<0.05)。不同处理下,15N分配率总体表现为粗根(d>2.0 mm)最高,茎最低。粗根与细根(0.5 mm

图6 不同时期各处理对胡杨幼苗氮素分配率的影响/%Fig.6 The effect of distribution rate of P. euphratica seedlings in different growth phase under different treatment

处理Treatment细根Fine root 粗根Coarse root 茎 Stem叶LeafD1N118.87±3.65abB35.74±4.34cdA11.46±1.25bcB33.93±2.36abAN228.20±4.57aA28.95±2.18dA5.30±1.70dB37.55±5.32aAD2N114.55±1.33bC54.48±1.45aA10.02±0.19cD20.95±0.81cBN210.09±2.58bC57.67±3.64aA11.73±0.46bcC20.51±0.76cBD3N113.51±0.94bC42.48±2.95bcA8.90±1.81cdC35.11±2.04aBN214.85±1.04bC50.29±1.64abA9.10±0.92cdC25.76±1.68cBD4N116.99±4.60bC41.46±1.47bcA14.83±0.90abC26.72±2.58bcBN213.54±4.76bC56.02±6.07aA18.12±2.65aC12.33±0.76dC
同一列不同小写字母表示同一植株器官不同处理下差异显著(P<0.05);同一行不同大写字母表示不同植株器官同一处理下差异显著(P<0.05)
2.5 干旱胁迫下胡杨幼苗各部分的氮素利用率
胡杨幼苗对氮素的利用率在不同生长期差异显著(P<0.05)。生长前期(0—50 d),幼苗氮素利用率随土壤水分增加呈先升后降趋势,在D3N1时显著增高,D1N2时最低,说明土壤水分严重匮乏时,其中的养分向植物中的转移受限,土壤中氮素有效性对土壤水分存在一个阈值。75 d时,随干旱胁迫的加剧幼苗氮素利用率显著降低(P<0.05),D1N2时幼苗的15N利用率最低,而D4N1处理下利用率最高。严重干旱胁迫下(D1N1),3个不同生长期的幼苗地下部分的利用率均高于地上部分,分别高出46.63%、52.59%、14.06%。相同干旱胁迫时,氮素量显著影响幼苗氮素利用率,始终呈现为N1显著大于N2,表明随施氮量增加利用率反而显著降低(P<0.05)(图7)。
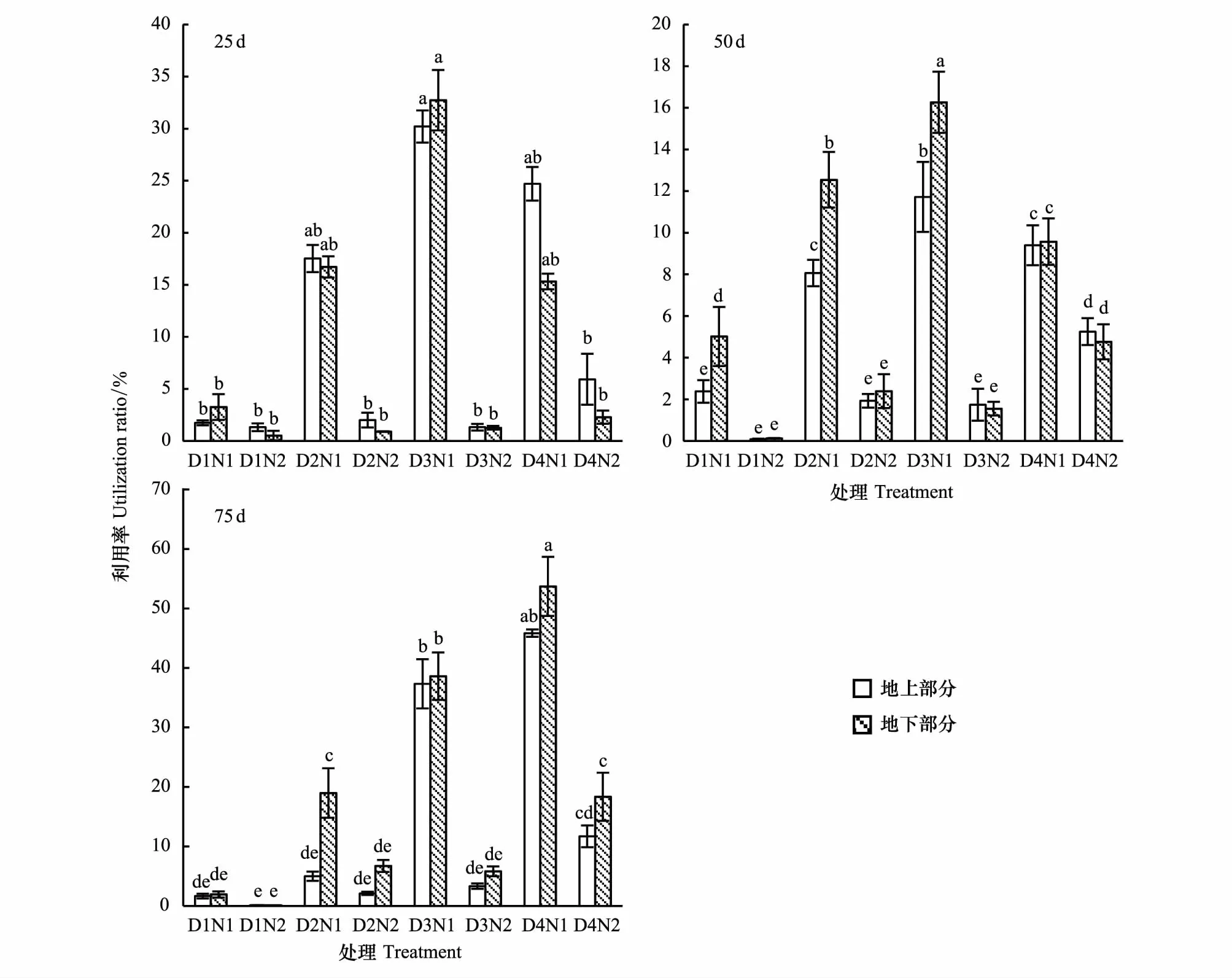
图7 不同时期各处理对胡杨幼苗氮素利用率的影响Fig.7 The effect of utilization rate of P. euphratica seedlings in differen growth phase under different treatments
幼苗各部分氮素利用率随干旱胁迫的缓解呈显著增加(P<0.05)。叶在D3N1时,氮素利用率最高,较D1N1增加了92.98%,而细根、粗根、茎在D4N1时最高,较D1N1处理时利用率分别增加了95.05%、96.47%、96.45%。说明土壤含水量的增加促进幼苗的氮素吸收,迅速提高利用率。同一处理时,各器官利用率总体上呈现出根>叶>茎的规律,粗根利用率均较高,茎的利用率最低(表3)。
3 讨论
在干旱区水资源是植株生存和生长的关键因子,尤其是幼苗生长初期对水分的需求量较大[15]。氮素是胡杨生态系统矿物质营养的主要限制因素,胡杨生长发育过程中受到氮素的影响[16]。植株根系对水土条件胁迫十分敏感,增加细根的根长及其表面积是大多数植物应对不利环境的基本策略[17]。本研究中,严重干旱胁迫极大地限制幼苗根系的生长,从而影响了根系对水分和养分的吸收利用,不利于后期的“水分自维持”,造成幼苗光合功能减弱,生物量积累锐减,降低了它在极端干旱时期的生存竞争力。反之,干旱胁迫缓解时,适量氮素(N1)增加了胡杨幼苗两级细根的长度和表面积,提高了根系的固着和吸收功能,水分和养分的高效利用促进植株生长,增加物质量积累。植物组织含水量直接影响植株生长、气体交换、光合功能等,维持较高的叶片组织含水量能增强植株抗旱性[18- 19]。我们发现,土壤水分亏缺较轻时叶片相对含水量维持较高水平,氮素作用不显著;但较重(D1)时,适量氮素(N1)能显著提高了叶片组织含水量,同时也增加叶片含氮量,促进类胡萝卜素合成,这有利于调控光合性能,防止强光和氧化胁迫损坏光合机构,增强植株抗性。另一方面,最大光量子产量Fv/Fm在添加氮素后显著提高,且D3N1时最高。这些结果共同说明水分和氮素的交互效应,能够缓解干旱胁迫对幼苗生存的威胁,塑造根系形态,增强自身的光竞争力,改善叶片的水分状况,提高幼苗耐受性,加强当年贮藏营养,赢得次年存活优势。
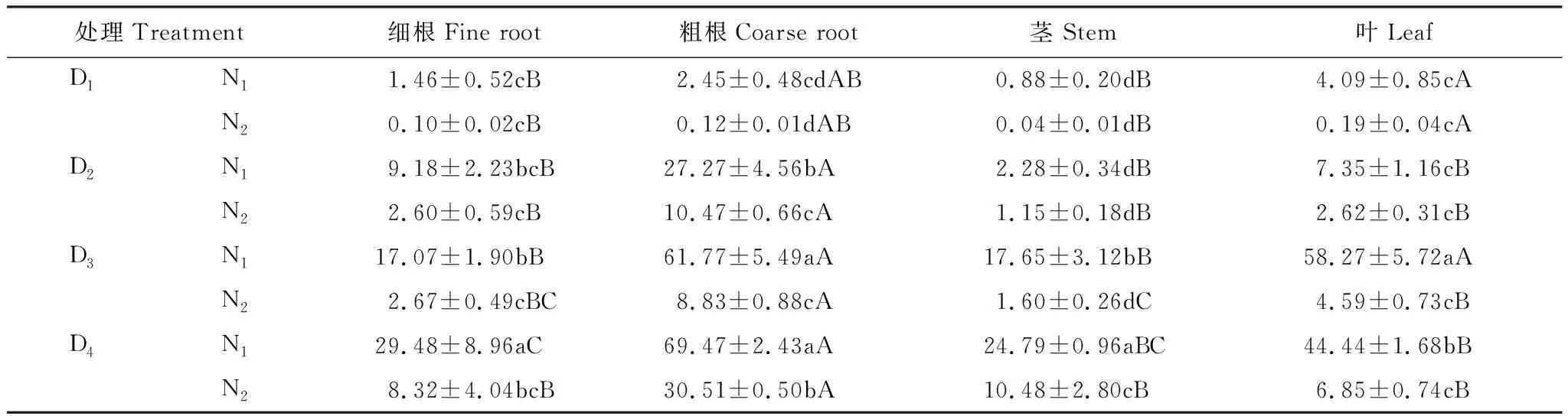
表3 不同处理下胡杨幼苗各部分的氮素利用率/%
运用稳定同位素示踪技术研究植物体内的氮素吸收和积累等特征已取得较大进展[20- 21]。植株受到干旱等环境限制因子时,能够通过调整自身物质分配,权衡地上和地下部分生长策略,以优化在胁迫环境中对有限资源的获取,维持必要的生存[22]。同时,前人研究认为植株的氮素吸收分配因氮素水平、器官部位和发育时期的不同而有很大差异[23- 24]。本研究中,胡杨幼苗各部分Ndff随干旱胁迫减轻呈增加趋势,随幼苗的不断生长,D3和D4处理能不断增加幼苗各部分Ndff,并且适量氮素(N1)使之增加显著,说明土壤水分能促进氮素的运移,增加根系形态塑造,增强幼苗的养分吸收能力,且氮素水平也影响各器官对养分的竞争吸收能力[25]。严重干旱胁迫时(D1),幼苗Ndff值较低,表明严重的干旱胁迫直接影响根系的发生和生长,根系活力降低,丧失对土壤中有效性氮素的吸收能力,导致地上部分新生器官生长缓慢甚至停滞。此外,幼苗生长25 d时,地上部分Ndff大于根系,而75 d时,根系的Ndff高于地上部分。说明幼苗生长早期叶片等地上新生器官作为生长中心对养分的竞争力较强,而生长后期碳水化合物向地下部分转运,促进根系的拓展,根系对氮素的征调能力增强。幼苗改变生长分配模式来适应环境以提高存活概率。
植株体内氮素分配和含量因器官部位和发育时期的不同差异显著[26]。25 d时,地上部分的分配率显著高于根系,此时叶片作为幼苗生长代谢最旺盛的部位,将更大比例的氮分配到营养器官,促进植株快速生长,这是幼苗期胡杨自身调节物质分配,保证植株的器官建成和生长。50 d时,分配率随干旱胁迫处理的不同在不同部位间存在显著的动态变化,这与幼苗地上和地下部分的权衡作用有关。在生长的不同阶段,幼苗生长中心的转移且各器官间氮素不断发生再分配,因此75 d时,根系的氮素分配显著增加,实现高的氮素利用率。并且,在相同处理下,粗根的分配率最高,这是由于在生长后期细根的养分吸收能力下降,使得各器官间的养分竞争更加激烈,大量内源性氮从叶中重新分配到粗根组织中作为贮藏养分[27]。这类贮藏养分在植株当年落叶休眠期贮藏于多年生枝干和粗根,次年调运到新生器官,用于枝叶等营养生长,提高存活概率。茎仅作为氮素贮藏的“临时库”,用于养分的运输通道,其分配率较低。因此氮素分布主要集中于生长活跃部位,直接影响着植株的氮素营养及生长发育状况。氮素水平对氮素在幼苗各部分的迁移分配影响较小,而对其利用率影响显著(P<0.05)。过量的氮素供应会引起植株内部各种营养元素不平衡,进而影响正常的生长发育。随氮素水平的增加,胡杨幼苗各部分的利用率显著降低,这与许多研究结相似[26- 28],幼苗在同一处理下利用率呈现根>叶>茎的规律,这充分说明胡杨实生苗的早期生长过程,通过增加养分分配及利用集中根系的发育,使幼龄胡杨能避开表层土壤的干旱胁迫。生长前期(0—50 d),叶片在D3N1时对氮素有较强的吸收能力并且呈现高分配高利用的状态;生长后期,粗根呈现高分配高利用的状态。总的来说,在干旱和半干旱区,水分和氮素普遍匮乏,二者也是胡杨幼苗渡过生长脆弱期的关键因素[21]。因此,在胡杨幼苗的不同生长阶段,通过权衡地上及地下的生长抵御不利环境的威胁而维持生存。生长早期增加地上部分的养分分配,促进新生器官的形态建成;生长后期增加根系的养分分配利用比例,促进植株根系的拓展发育,为越冬及次年的萌发做准备。“厚积薄发”的生长分配模式是胡杨实生苗存活的必要条件。干旱抑制幼苗吸收利用氮素,但氮素的吸收利用通过促进根系发育也能增强植株抗旱性。
4 结论
干旱胁迫和氮素水平共同影响着胡杨幼苗的各部分的养分吸收利用。胡杨幼苗在土壤相对含水量60%—65%(D3)、氮素添加量3 g/盆(N1)时,其生长表现为最佳状态;氮素水平N1较N2能显著提高幼苗各部分的Ndff和利用率;生长前期(25—50 d),叶片在D3N1时对氮素有较强的吸收能力并且呈现高分配高利用的状态,生长后期,粗根呈现高分配高利用的状态。因此,适量的氮素供给对干旱胁迫下的胡杨实生苗生长具有补偿效应,植株在不同生长阶段能通过调整自身物质分配的可塑性,优化限制性资源获取,实现早期形态建成,有效地帮助幼苗渡过脆弱期。
