黑河流域高寒草甸生态系统水分收支及蒸散发拆分研究
2018-11-15童雅琴佩1李小雁1张赐成
童雅琴,王 佩1,,李小雁1,,*,张赐成,白 岩
1 北京师范大学地表过程与资源生态国家重点实验室,北京 100875 2 北京师范大学地理科学学部自然资源学院,北京 100875
根据IPCC第五次评估报告,自工业革命以来,全球气候变化是当前人类面临的重要议题,其主要表现为气温升高和降水格局的变化[1],是改变水循环的重要驱动因素。水分收支是对水循环要素降水、蒸发蒸腾、径流以及土壤储水量变化等的定量刻画[2]。在水分收支过程中,降水是陆地生态系统水分的主要来源,直接影响植被生产力;蒸散发(ET)包括土壤蒸发(E)和植被蒸腾(T)[3],是陆地生态系统水分的主要耗散形式,土壤蒸发和植被蒸腾受不同的生物物理过程控制[4],对于其在生态系统中的相对贡献的定量研究对准确预测生态系统功能对气候变化的响应至关重要,当前关于生态系统蒸散发拆分的方法包括实地观测法[5- 6],同位素法[7- 10]以及模型估算法[11- 13];土壤水分是土壤-植被-大气连续体的关键因子[14- 15],土壤贮水量对维持“土壤水库”功能,调节降水分布不均等具有重要作用[16]。高寒草甸/草原占中国西部高寒区面积的64%[17],被认为是对气候变化最为敏感的区域[18- 19]。在气候变化背景下,准确评估其水分收支过程对理解气候、植被、水文交互作用至关重要。
黑河流域是典型的干旱区内陆河流域,是以水循环过程为纽带的冰川/冻土、绿洲和荒漠为主要特征的多元生态系统[20],上游高寒区被认为是干旱区水塔,水循环过程复杂。已有的研究多是从流域尺度运用模型进行水分收支估算,或者侧重水分收支的个别要素[21- 24],如Jia等[25]用分布式水文模型分析了黑河中上游水分收支和能量收支平衡,表明在上游地区年均蒸散发和年均径流分别占年均降水的63%和37%;Qin等[26]表明在黑河上游52.46%的出山径流量产生于裸地景观,上游森林景观几乎不产流;Yang等[27]运用分布式生态水文模型研究表明,黑河上游地下水径流和壤中流占主导,地表径流极少,且水分收支特征受植被类型的影响显著;以上模型虽然能很好的分析流域尺度水分收支,但是,一般流域尺度上空间异质性较强,缺少在生态系统尺度上对水分收支过程的验证,且缺少直接观测的生态系统实际蒸散发,可能导致对生态系统水分以及植被需水量的估算存在偏差。涡动相关技术实地观测的水汽通量数据,为准确的分析生态系统水分收支提供了可能,且当前运用双源模型对蒸散发进行拆分来评价生态系统水分收支状况的研究较少。因此,本文以黑河流域高寒草甸生态系统为研究对象,运用蒸散发拆分模型和涡动相关技术直接观测的水汽通量数据,估算2014和2015年黑河上游高寒草甸生态系统水分收支及其组分的季节变化和年际变化旨在定量评估黑河流域高寒草甸生态系统的水分收支状况,为流域水资源承载力评估及水资源管理提供科学依据。
1 研究区概况与数据来源
1.1 研究区概况
黑河是我国第二大内陆河,发源于青海省祁连山区的冰川积雪带,流域总面积14.3万km2,是集合高山冰川、森林草原、平原绿洲及戈壁荒漠的复合生态系统[28]。本研究站点位于黑河上游青海省祁连县中东地区的阿柔乡,黑河上游支流八宝河南侧的河谷高地上,经纬度为100.46°E,38.05°N,海拔3044 m,周围地势相对平坦开阔,自东南向西北略有倾斜下降,南北两侧约3km外是连绵的山丘和高山。研究区多年平均降水为400.3 mm[21],降水量年内分配不均,主要集中在生长季(5—9月)。研究区属于高寒草甸区,植被类型以小嵩草草甸为主。
1.2 数据来源
研究数据包括2014和2015年自动气象站数据,涡动相关数据及相关的遥感数据,本研究的气象数据和涡度相关数据来源于阿柔冻融观测站,黑河综合遥感联合试验:水文气象观测网数据集(http://hiwater.westgis.ac.cn/)[29]。其中,涡动相关数据用来计算实际蒸散发和检验模型的效果,该数据是经过Edire软件后处理的30 min通量数据,处理过程包括野点剔除、坐标旋转、频率响应修正、WPL修正以及初步质量控制等,本文对缺失的涡动数据运用查表法和平均昼夜法进行插补[30]。气象数据包括气温、降水、相对湿度、太阳辐射、风速、土壤温湿度(0—320 cm)等[31- 32]。其中,自动气象站的降水采用翻斗式雨量计观测,翻斗式雨量计安装在阿柔超级站40 m观测塔上。土壤温湿度探头埋设在地表0 cm和地下2、4、6、10、15、20、30、40、60、80、120、160、200、240、280、320 cm处,用于土壤水分和温度的观测。叶面积指数(LAI)数据来自2014和2015黑河生态水文遥感试验:黑河流域1km分辨率5d合成叶面积指数(LAI)数据集(http://www.heihedata.org/data)[33- 35],模型中需要的日叶面积指数(LAI)采用三次样条函数插值所得,植被高度数据参考黑河流域生态水文样带调查:2013年上游植被数据[36]。
2 研究方法
2.1 水分收支计算
陆地生态系统水分收支的基本方程[37]为:
P+Sd+I+R上入+R下入=ET+R上出+R下出+D+ΔW
(1)
式中,P为降水量,Sd为地下深层补给量,I为灌溉量,R上入、R下出分别为计算时段内经地面和地下流入生态系统内的径流量;ET(ET=E+T)为蒸散发,包含有土壤蒸发E和植物蒸腾T,R上出、R下入分别为计算时段内经地面、地下流出的径流量,ΔW为土壤贮水量的变化量,D为土壤水分的下渗。在祁连山高寒草甸没有灌溉且地下水埋藏较深,可将I和Sd忽略,本文用于计算土壤贮水量的是0—320 cm处的土壤水分,所以可将D忽略,R上入、R下入和R上出、R下出可分别表示为R入和R出,所以水分收支公式可以简化为:
P+R入=ET+R出+ΔW
(2)
简化后公式的物理意义可以理解为到达生态系统的水分消耗于土壤和植被的蒸散发、径流以及土壤贮水量的变化,若P-ET+ΔW>R出-R入>0表示生态系统水分盈余;反之,表明生态系统水分亏缺。
2.2 土壤贮水量及其变化的估算
土壤贮水量(W)的计算采用各层土壤体积含水量的平均值与其代表的深度乘积累加求得[38],具体计算公式如下:
(3)
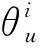
2.3 蒸散发组分估算
本文使用Wang和Yamanaka双源模型[39],该模型以植被冠层和土壤表面能量平衡为基本原理,包括太阳辐射传输过程,植被气孔导度的模拟,而且其在不同的生态系统对蒸散发的模拟与分割都取得了很好的结果[39- 41]。模型的主要公式表征如下:
(4)
(5)
式中,RnV是植被冠层的净辐射(W/m2),HV是植被冠层的显热通量(W/m2),T是蒸腾量 (kg m-2s-1),fV为植被冠层的电介常数,Sd是向下的短波辐射(W/m2),Ld是向下的长波辐射(W/m2),σ为 Stefan-Boltzmann常数(=5.67×10-8W m-2K-4),TG和TL分别为地表温度和叶片表面温度(℃),RnG为地表净辐射(W/m2),G是土壤热通量(W/m2),HG是地表显热通量(W/m2),E是蒸发通量(kg/ m2s),αG为地表反照率,αV是植冠层反照率,αG,αV可近似为常数0.1和0.2。
能量平衡中的显热通量可以通过以下等式表示:
HV=ρa(TL-Ta)/raV
(6)
HG=ρa(TG-Ta)/raG
(7)
式中,cp为干燥空气的热容量,raV为植被冠层的空气动力阻抗,raG是地表的空气动力阻抗(s/m),它们主要与风速,植被高度和风速测量仪的高度有关,详细描述可见王佩等[39]研究。
模型结果中的土壤蒸发(E)和植被蒸腾计算公式如下:
T=ρa([q(TL)-qa])/(raV+rc)
(8)
E=ρa([q(TG)-qa])/(raG+rSS)
(9)
式中,qsat(TL) 为叶片温度的饱和比湿 (kg/kg),qsat(TG)为地表温度的饱和比湿,qa为空气比湿,raV是植被冠层的空气动力阻抗,rc为气孔导度(s/m),以及rSS是土壤表面阻抗(s/m)。
rc=rst/LAI
(10)
式中,rst为叶片气孔阻抗。
(11)
式中,rst_min和rst_max分别为土壤完全湿润时叶片气孔阻力的最大值和最小值,公式中的常数CSW可表示为:
(12)
式中,θ为土壤体积含水量(m3/m3),θmax是土壤最大体积含水量。土壤的表面阻力与土壤含水量满足指数关系式:
rsoil=a(θs/θ)b+c
(13)
式中,a,b,c为经验系数,a=3.5,b=2.3 和c=433.5。
2.4 模型的敏感性分析
为了评价模型指定参数和输入变量可能存在的误差,对模型进行敏感性分析,敏感性系数si[39,42- 43]定义如下:
(14)
式中,Pi是指影响模型输出结果(如ET或者T/ET)的i个驱动参数或变量,其偏导∂O/(∂Pi)计算公式如下:
(15)
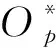
3 结果分析
3.1 模型的有效性和敏感性分析
图1为高寒草甸涡动相关系统观测的和模型模拟的蒸散发以及地表温度值的1∶1线性拟合关系,结果发现模型模拟值与实测值之间拟合度高,相关系数为分别0.90和0.89,表明该双源模型对高寒草甸生态系统蒸散发的模拟是有效的。
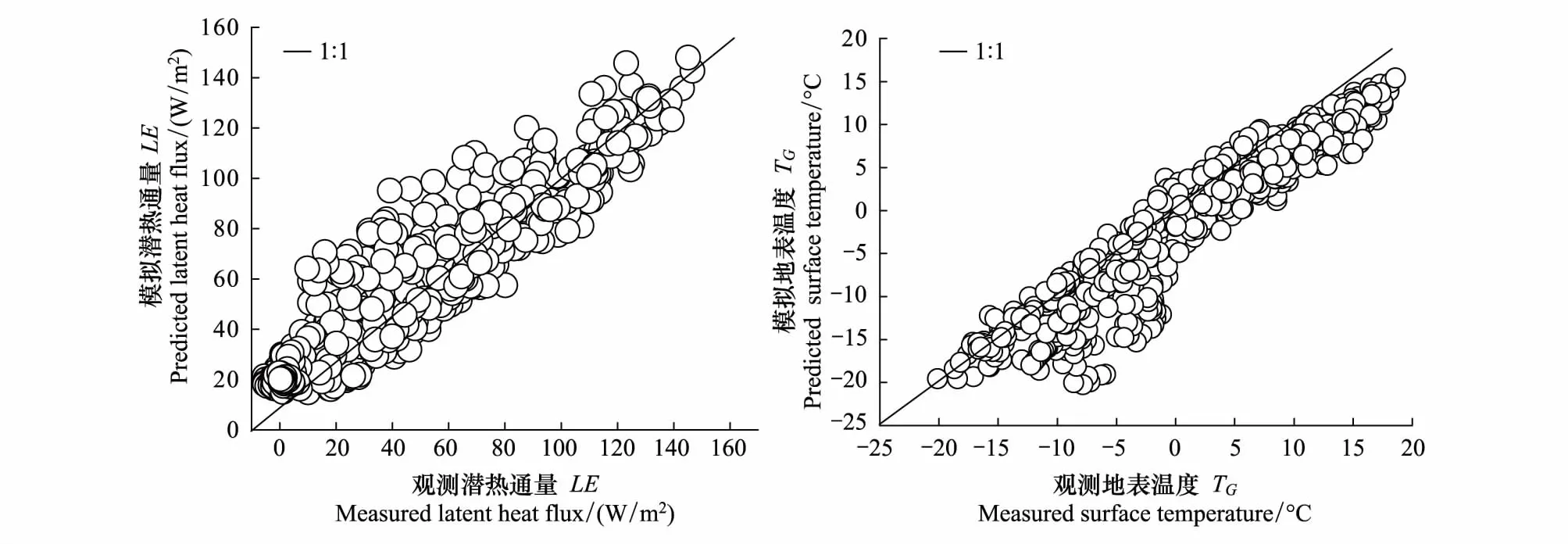
图1 实测值与模型模拟蒸散发(潜热)值与温度对比2014—2015Fig.1 Comparison of evapotranspiration (latent heat flux) and temperature between measured and the predicted by model during 2014—2015
表1总结了高寒草甸潜热通量(LE)和蒸散比(T/ET)对模型输入变量的敏感性。结果表明潜热通量(LE)对相对湿度(RH)最为敏感,为-0.74 ± 0.32,即相对湿度5%的误差会导致潜热通量平均3.7%的误差,对于蒸散比T/ET,叶面积指数(LAI)和土壤水分(θ)对其较为敏感,其5%的误差分别可导致蒸散比T/ET2.2%和1.7%的误差。因此可以看出模型估算的蒸散发(LE)和蒸散比(T/ET)存在的误差都在可以忽略的范围之内,表明模型模拟的结果较为可靠。
表1模型输入参数对蒸散发(ET)和蒸散比(T/ET)敏感性系数(Si)的平均值±标准偏差(SD)
Table1Meanandstandarddeviation(SD)ofthesensitivitycoefficients(Si)ofevapotranspiration(ET)andtranspirationfraction(T/ET)fortheinputvariablesinmode
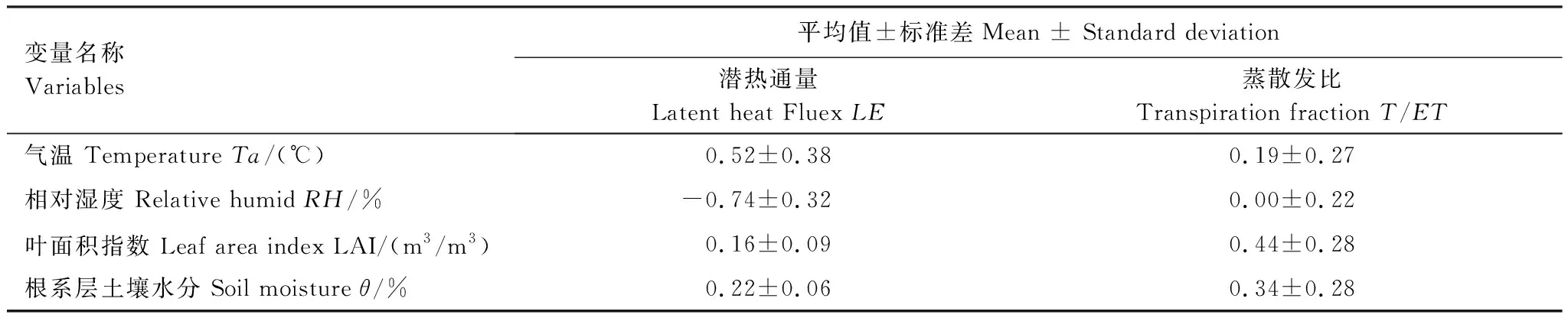
变量名称Variables 平均值±标准差Mean ± Standard deviation潜热通量 Latent heat Fluex LE蒸散发比 Transpiration fraction T/ET气温 Temperature Ta/(℃)0.52±0.380.19±0.27相对湿度 Relative humid RH/%-0.74±0.320.00±0.22叶面积指数 Leaf area index LAI/(m3/m3)0.16±0.090.44±0.28根系层土壤水分 Soil moisture θ/%0.22±0.060.34±0.28
3.2 高寒草甸生态系统水分收支要素特征
3.2.1 降水特征分析
高寒草甸生态系统2014和2015年降水量为分别为520 mm和401 mm,降水量年内分配不均,降水主要集中在5—9月(图2),2014年和2015年生长季降水分别占全年总降水量的88.75%和90.21%,2014和2015年均温度分别-0.43℃和0.28℃,气温、饱和气压差(VPD)以及潜在蒸散发(PET)的季节变化趋势一致,呈现单峰变化规律,都是从5月开始上升,7月达到最大值,随后开始缓慢下降。
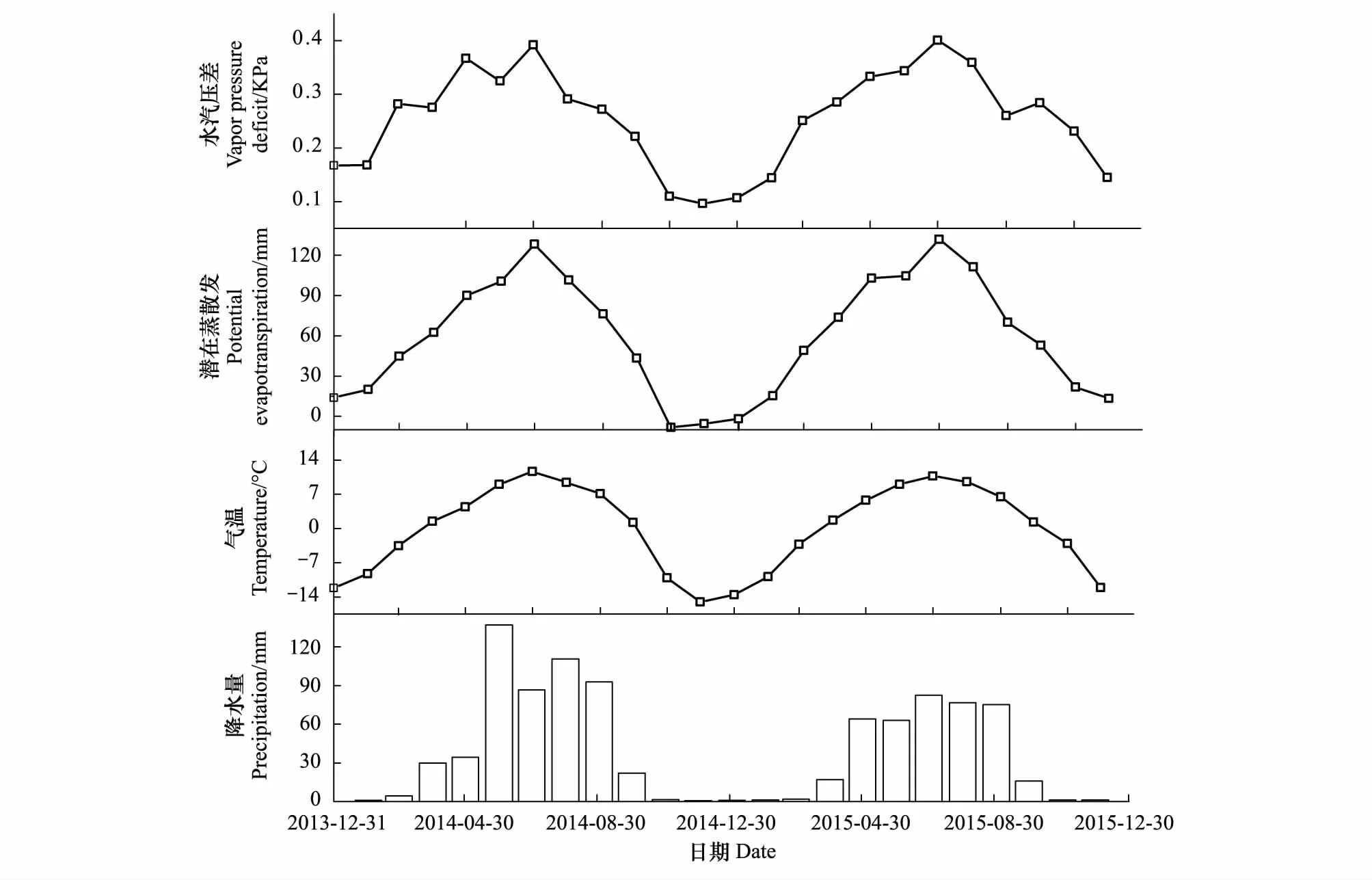
图2 2014—2015年研究区气象要素的季节变化Fig.2 Monthly variation of meteorological variables in study area from 2014 to 2015
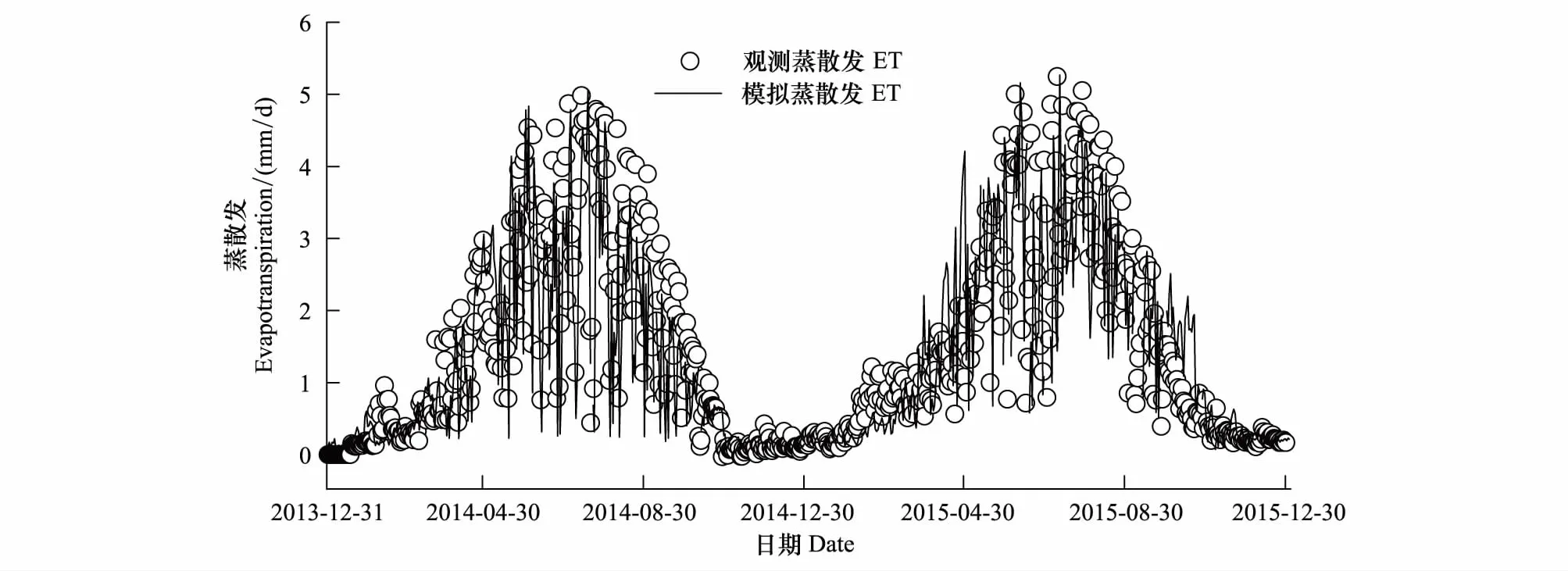
图3 2014和2105年模拟蒸散发与观测蒸散发对比Fig.3 Comparison of Predicted evapotranspiration and simulated evapotranspiration in 2014 and 2015
3.2.2 蒸散发及其组分变化特征分析
通过涡动相关观测的高寒草甸生态系统2014年和2015年的实际蒸散发量分别为520 mm和551 mm,生长季(5—9)蒸散发分别为408 mm和422 mm,分别占全年蒸散发总量的78%和76%,图3显示为高寒草甸蒸散发的时间序列,从中可以看出观测蒸散发、模型模拟蒸散发在时间变化上具有较好的一致性。日蒸散发量在全年尺度上呈现明显的单峰变化规律,4月份蒸散发量开始增加,7月达到全年最大值,2014年和2015年月最大蒸散发量ET分别为85 mm和89 mm,日最大蒸散发量分别为5.30 mm/d和5.24 mm/d,随后开始下降,这与生长季植被物候变化特征相一致。
已有研究表明植被蒸腾(T)是陆地生态系统水分收支的重要组分,蒸散比(T/ET)可以很好的指示生态系统的生产力。通过模型对蒸散发的拆分发现,在高寒草甸生态系统2014年和2015年蒸散比(T/ET)的日变化也呈现明显的单峰变化规律(图4),在非生长季,缺少植被覆盖,蒸散发(ET)主要是土壤蒸发,蒸散比(T/ET)很小0.3左右;在生长季(5—9月),随着植被的生长,植被盖度增加,植被蒸腾在蒸散发中占主导,T/ET迅速增加,2014和2015年生长季平均蒸散比(T/ET)分别为0.74和0.79,日最大值可达0.91。
3.2.3 土壤水分变化特征
2014年和2015年高寒草甸生态系统0—320 cm土壤贮水量季节及年际变化特征如图5所示,从图中可以看出该高寒草甸生态系统土壤贮水量存在年内波动,呈现双峰变化规律。每年3月开始,土层解冻再加上积雪融化,土壤贮水量快速增加,到5—6月份达到每年的最大值,7—8月草甸生态系统具有较强的植被蒸腾,植被的生长消耗了土壤中储存的水分,使得土壤贮水量略有下降,9—10月蒸腾减弱,土壤贮水量又有所上升,每年11月至次年3月份,受土层冻结的影响,实测土壤含水量仅为土壤中的液态含水量,固态水无法探测,该时段内的土壤贮水量很小,即土壤贮水量受冻融作用影响显著。图6为不同深度冻融前后土壤体积含水量的变化,表层至10 cm处土壤体积含水量冻融前后变化显著,消融后土壤体积含水量明显高于冻结前,20 cm以下至120 cm处则相反,冻融前后土壤体积含水量变化不大,至160 cm处土壤无冻融,表明冬季冻结对高寒草甸生态系统表层土壤水分保持具有重要作用。
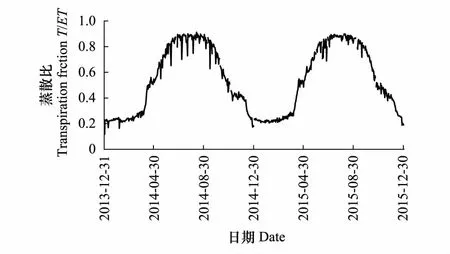
图4 2014和2015年高寒草甸蒸散比(T/ET)的日变化 Fig.4 The diurnal variation of transpiration fraction T/ET of Alpine meadow ecosystem in 2014 and 2015
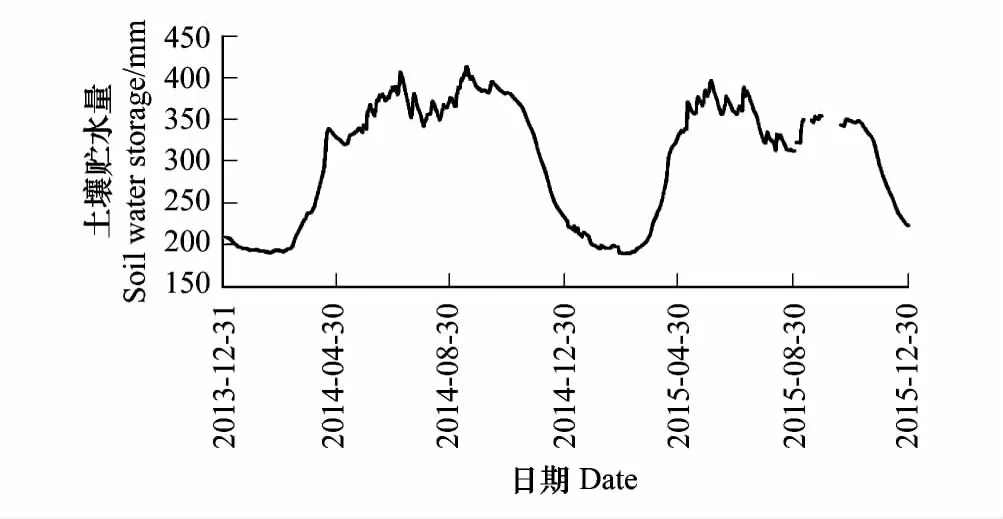
图5 2014和2015年高寒草甸0—320 cm土壤贮水量的日变化 Fig.5 Diurnal variation of soil water storage (0—320 cm) of Alpine meadow in 2014 and 2015
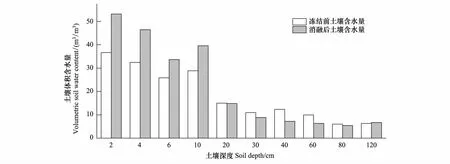
图6 高寒草甸生态系统不同土层深度冻结前消融后土壤体积含水量Fig.6 Compare to the soil water content between the before freezing and after thawing at different soil depth
在垂直变化上,高寒草甸生态系统土壤体积含水量在0—40 cm处波动较大(图7),越往表层,土壤含水量越高,40 cm以下土壤体积含水量小(10%以下)且变化不大,说明0—40 cm为高寒草甸生态系统主要的贮水层。从年际变化上看,2014年该高寒草甸生态系0—320 cm处的土壤贮水量大于2015年,表明土壤贮水量的变化深受当年降水量的影响。
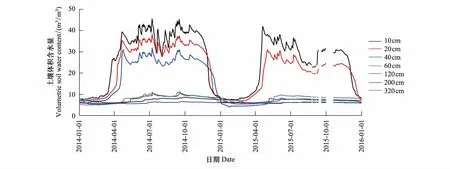
图7 不同深度处的土壤体积含水量的变化Fig.7 Variation of volumetric soil water content at different depths
3.3 高寒草甸生态系统水分收支特征
图8为2014和2015年高寒草甸生态系统水分收支要素降水(P),蒸散发(ET)和土壤贮水量变化(ΔW)的月变化及水分收支状况,每年10月至次年2月,观测到的降水量与蒸散发量很小,土壤贮水量的变化由于冬季土层冻结,使(P-ET-ΔW)为正值;在每年3—5月,春季气温回升,再加上降水少,该时段冻结的土层及冬季降雪相继融化,补充土壤水分,土壤储水量急剧增加,导致(P-ET-ΔW)为负值,可见,水分收支的季节变化受冻融过程影响显著;在完全不受冻融影响的月份6—9月(表2),2014年降水较多,生态系统有地表径流形成,6—9月地表径流量分别为6、15、0.45和22 mm;2015年降水量少,6—8月份,蒸散发不仅消耗全部的降水,还消耗储存在土壤中的水分,生态系统呈现水分亏缺,6—8月亏缺量分别为22、12、13 mm,9月份到植被生长的末期,植被蒸腾减弱,该时段生态系统产流量为7 mm。
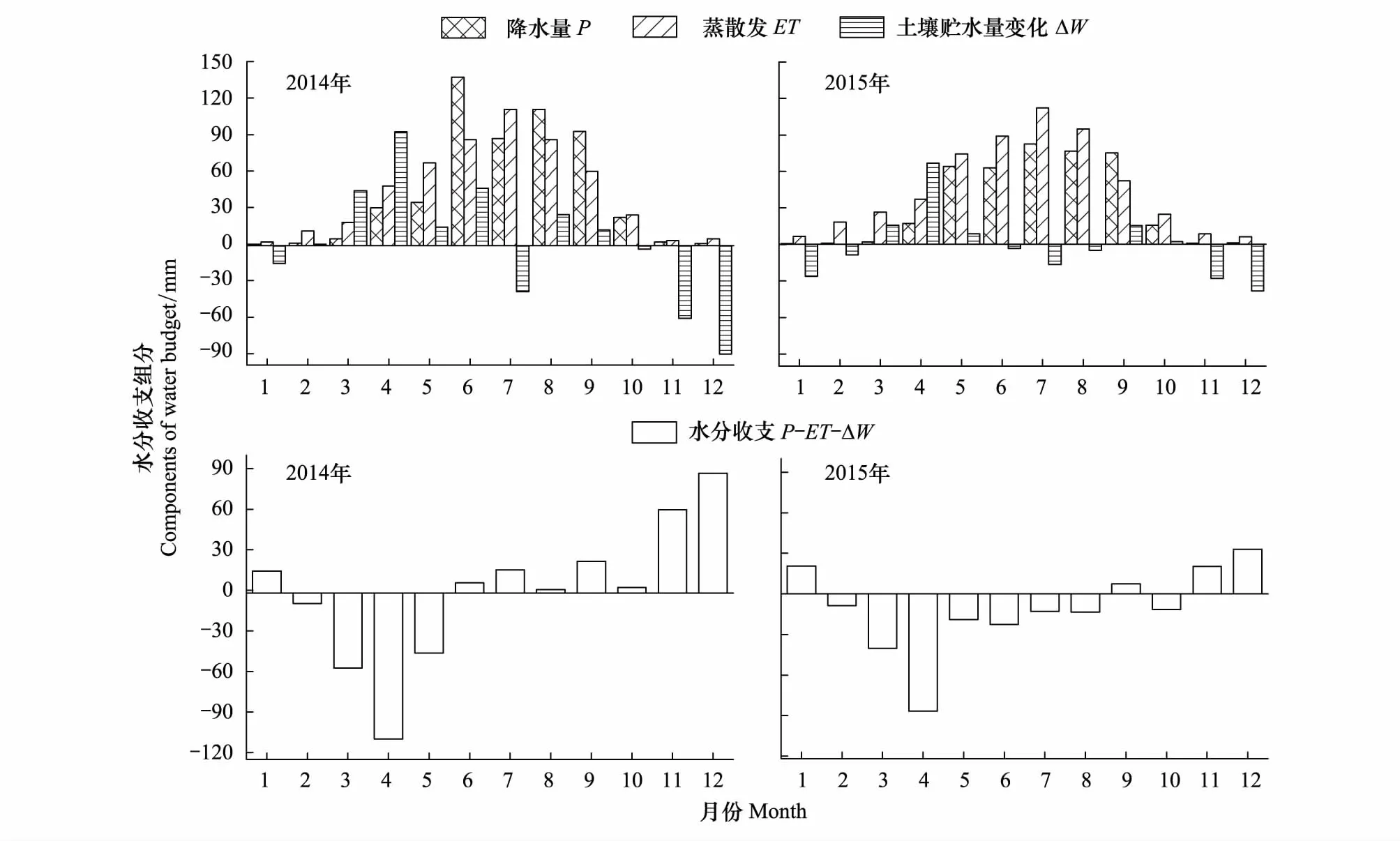
图8 2014和2015高寒草甸生态系统水分收支月变化Fig.8 Monthly variation of water budget in Alpine meadow ecosystem in 2014 and 2015
从表3可以看到,在全年尺度上,2014年和2015年高寒草甸呈现不同程度的水分亏缺,2014年降水多,亏缺量为18 mm,2015年亏缺量为134 mm;在完全不受冻融影响的月份,2014年该高寒草甸生态系统产生地表径流量为42 mm;2015年降水量少,生态系统呈现为水分亏缺,总的亏缺量为26 mm。
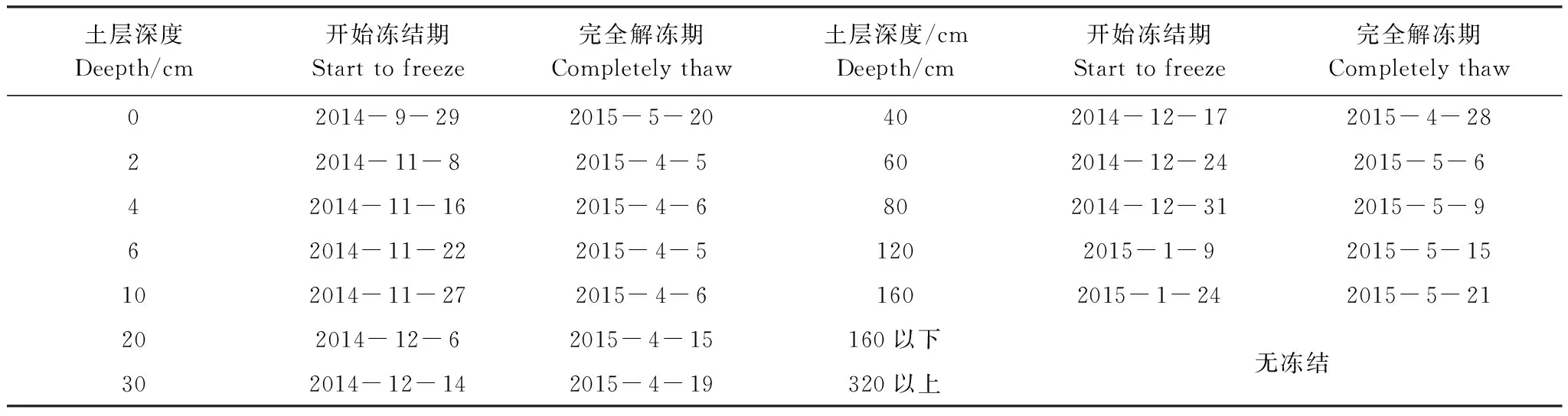
表2 阿柔高寒草甸2014至2015年冻融起止时间
根据不同深度土壤最高温度(Tmax)和最低温度(Tmin),秋季始冻期(Tmax>0℃且Tmin<0℃)、完全冻结期(Tmax<0 ℃且Tmin<0℃)和春季解冻期(Tmax>0℃且Tmin<0℃),完全解冻(Tmax>0℃且Tmin>0℃)。
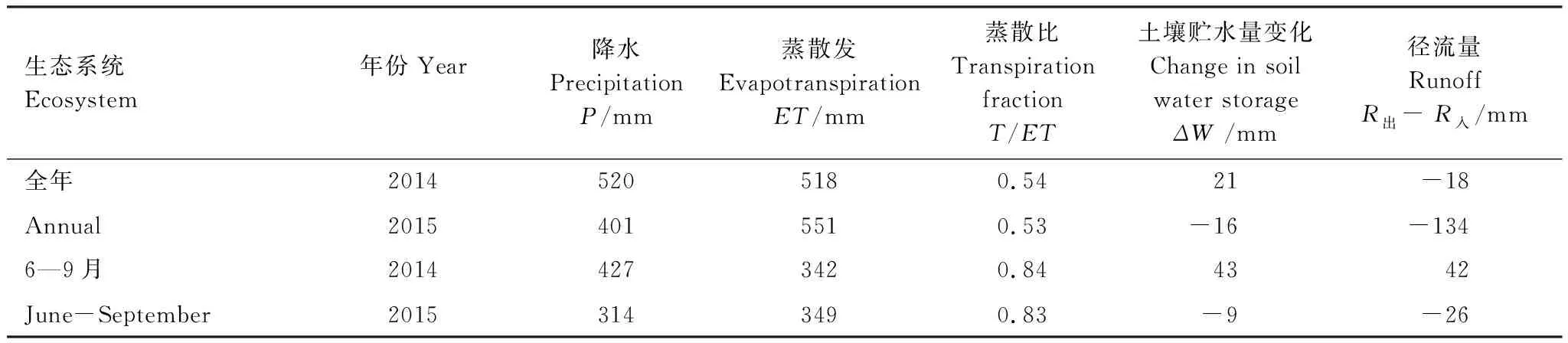
表3 2014和2015高寒草甸生态系统水分收支要素年变化
4 讨论
4.1 蒸散比T/ET及影响因素分析
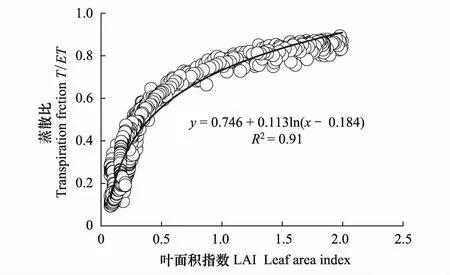
图9 叶面积指数(LAI)与蒸散比(T/ET)的关系 Fig.9 Relationship of LAI with daily mean transpiration fraction (T/ET)
通过对蒸散发ET的拆分发现在高寒草甸生态系统植被蒸腾,尤其生长季植被蒸腾是生态系统水分消耗的主要形式,2014和2015年高寒草甸生态系统年均蒸散比(T/ET)分别为0.54和0.53。Wei等[44]研究表明在全球尺度上T/ET为0.57 ± 0.06。Hu等[4]基于S-W双源模型研究的2004年高寒草甸的T/ET为0.44,王海波等[45]基于Penman-Monteith模型研究发现2008—2009年高寒草甸T/ET约为0.6。本研究估算T/ET与其他相关研究基本接近,但也存在一定的差异,可能与LAI数据集有关,已有研究表明T/ET在季节尺度上主要受植被叶面积指数(LAI)的影响[4,39,44,46- 48],这与我们的研究结果相一致(图9),叶面积指数LAI与T/ET呈现明显的对数关系。Wei等[44]表明,不同的来源的LAI数据集,可能会导致T/ET存在一定的误差,因此,高精度的LAI数据对蒸散发ET拆分影响重大。
4.2 水分收支估算
本研究利用水量平衡方程对高寒草甸生态系统2014和2015年水分收支进行估算。在每年的非生长季,各月蒸散发量略大于降水量,生态系统水分亏缺,这与Yao等[49]运用2004—2011实测数据对海北高寒草甸生态系统水分收支研究结果相一致。在2014年不受冻融影响的生长季,生态系统水分盈余。在正常年份的生长季,生态系统水分呈现亏缺,蒸发和蒸腾不仅消耗全部的降水,还消耗储存在土壤中的水分,储存在土壤中的水分对生态系统水分收支起重要调节作用。Yan等[50]通过对森林生态系统的研究发现,土壤水分是旱季蒸散发的重要水源,这与本研究结果较为一致。在年尺度上,阿柔站高寒草甸生态系统并无地表径流产生,只有在降水量大的湿润年份的生长季,有地表径流形成。Yang等[51]运用分布式水文模型对黑河上游水量平衡研究表明2001—2012年,地下水径流占总径流量的80%左右,壤中流占20%左右,地表径流量极小,占径流量的2%—3%。陈仁升等[17]在黑上游高寒地区通过长时间小流域观测实验表明,高山寒漠带为山区流域的主要产流区,而高寒草甸/草原区径流贡献很少,其水源涵养功能大于水文功能;通常平缓地形下的高寒草甸/草原具有较多的水源涵养功能,对径流的贡献少[17],这与本研究结果相当。
在高寒草甸生态系统,土壤水分的剧烈变化主要发生在0—40 cm处,与郭淑海[45]在天山高寒草甸的试验结果一致,0—40 cm土壤贮水,是蒸散发消耗的重要水源,对维持植被生长、发育具有重要作用。此外,在生态系统水分收支中,外界径流补给是生态系统水分消耗的重要来源,而且已有研究[52- 53]表明高寒草甸生态系统地下水埋藏较深,且壤中流较为发育,已有相关研究[26,54-55]表明黑河上游,受冻土的影响,流域产流以壤中流为主,因此,高寒草甸生态系统亏缺的水分可能是壤中流补给。
5 结论
本文运用双源模型对黑河上游高寒草甸生态系统蒸散发组分进行拆分及以生态系统水量平衡方程为原理,估算了黑河流域高寒草甸生态系统2014年和2015年水分收支状况,得到以下结论:(1)生长季植被蒸腾是生态系统水分主要的消耗形式,在季节尺度上,蒸散比(T/ET)主要受叶面积指数(LAI)的控制;(2)0—40 cm土壤储水对维持生长季旺期植被蒸腾具有重要的意义;(3)在湿润年(2014年)高寒草甸生态系统水分收支基本平衡,且生长季有产流;在正常年份(2015年)生态系统呈现水分亏缺,亏缺的水分主要来自外界壤中流的补给。总体而言,实测数据可以准确的刻画生态系统水分收支过程,但也存在一定的不确定性,如仪器的观测精度和观测年限等,因此,在未来的研究中应加强对生态系统的连续观测,为遥感数据和模型模拟研究提供更精确地验证数据。
