基于高光谱的晚疫病胁迫下马铃薯叶片的过氧化物酶活性检测
2018-11-15胡耀华李清宇
胡耀华, 李清宇, 唐 翊
(1. 西北农林科技大学 机械与电子工程学院, 陕西 杨凌 712100; 2. 西北农林科技大学 农业部物联网重点综合实验室, 陕西 杨凌 712100)
在生产实践中,马铃薯晚疫病严重制约了我国马铃薯的产量和质量,是危害最为严重的几种病害之一.叶片染病后,先是形成不规则的黄褐色斑点;气候潮湿时,迅速扩展至主蔓或叶柄,导致叶片萎蔫蜷缩,全株焦黑呈尸腐状;由于其流行度大,防治难,经常造成无法预估的重大损失[1].因此,能够及时、准确地检测出马铃薯晚疫病是该病害防治中的重要内容.
过氧化物酶(peroxidase, POD)在植株体内普遍存在,与植株的各种重要生理活动都有着密切关系.因此,POD的酶活性可反映出植物的生长状况是否良好[2-3].遭受病原菌侵害或逆境的植株会打破内部原有平衡,所产生的大量活性氧会对植物的生理结构造成破坏.POD能把有毒的过氧化氢(H2O2) 转化为无害的H2O, 其活性值也会相应发生一系列改变,因此被广泛用于评价植物的抗逆性.目前实验室检测作物中POD活性的方法主要是化学比色法,但是化学比色法的预处理工作复杂繁琐、且要对样本进行破坏性处理,过程耗时费力,不能实现连续性监测.因此,研究一种能够快速无损测定作物中POD活性的方法十分必要.
高光谱成像技术已经在植物叶片中抗氧化酶系统的活性预测方面有所涉及[2-4].朱文静等[2]在建立番茄叶片中过氧化物酶(POD)活性的预测模型时,运用了高光谱技术与化学计量学结合的方法,实现的准确率高达93%以上;杨燕等[3]的研究表明,高光谱图像能够反演出水稻稻瘟病潜育期的抗氧化酶(SOD)活性,并可以由此推断出病害胁迫程度信息是可行的;谢传奇等[4]对茄子灰霉病叶片进行高光谱测定,结果表明基于PLSR模型推荐的隐含变量建立的BP神经网络模型对灰霉病胁迫下用过氧化氢酶(CAT)活性定量检测的准确率最高.但在马铃薯晚疫病研究中,还没有关于叶片中POD活性的研究成果.
笔者对感染马铃薯晚疫病的叶片在不同病害程度下的高光谱信息和POD活性值进行检测,利用高光谱信息和化学计量学相结合的方法,建立马铃薯晚疫病叶片中POD活性的预测模型,并通过POD值来判断叶片是否染病及染病程度,为晚疫病的早期防治简易检测装置的研发提供参考.
1 试验部分
1.1 仪器及设备
1.1.1 高光谱成像采集系统
试验采用了北京卓立汉光仪器有限公司研制的Zolix Hypersis农产品高光谱成像系统(装置示意图见图1).
由图1可见,该成像系统由 V10E-QE型可见/近红外波段光谱成像仪、光源、XEVA2616型面阵CCD相机、PSA300-X型电控平移台装置以及计算机等部件组成.所使用的光谱仪的波长范围为375~1 018 nm,光谱分辨率是2.8 nm;试验的采样间隔是0.65 nm,图像分辨率336 nm×256 nm.为了减小环境光线对测量的影响,将整个系统放置在暗箱内.该仪器的工作过程可简述如下:光源发射出光,照射在放置于电控平移台上的待测样本上,所形成的图像被相机捕捉到,成为样本高光谱图像.电控平移台带动样本连续运行以获取样本连续的图谱信息,这些信息被传送并存储到计算机中,以便进行后续分析.
采集数据前,设定如下参数:曝光时间为50 ms;电动平台移动速度为20 mm·s-1;样本到相机镜头的距离为65 cm.首先要让系统进行30 min的预热,再开始采集高光谱图像;同时,利用黑白校正来消除光照条件的变化对成像的影响,以及基线漂所导致的误差.具体操作方法如下:用标准反射板(聚四氟乙烯材料)采集的全白图像标定为W;盖上镜头盖采集的全黑图像标定为B;通过样本采集到的原始图像标定为I;则校正后图像R[5-6]的计算式为
(1)
1.1.2 POD酶活性测定设备
通过对比,最终决定使用紫外分光光度计法对POD的酶活性进行测量[7].测定所需设备如下:超低温冰箱(海尔BCD-226SDCZ,中国),冷冻温度-40~-20 ℃;冷冻离心机(安亭TGL-16G-A,中国),最大转速16 000 r·min-1,工作温度-5~30 ℃;紫外分光光度计(UV1102,中国),来测试190 nm到1 100 nm之间的波长.POD活性由紫外分光光度计在离心后加入试剂测得,具体测定方法见参考文献[7].
1.2 样本制备与酶活性测定
试验选用中薯18号叶片,属晚疫病中感品种,广泛种植于西北地区.使用盆栽的方式进行培养.培养10盆,平均每盆栽种3株,共种植30株.2017年6月,盆栽种植在西北农林科技大学科研用玻璃温室中,经过日常管理,3个月后开始试验,选取质量为0.2~0.4 g、且大小基本一致的100片叶片进行离体接菌.试验所使用的马铃薯晚疫病孢子悬浮液由西北农林科技大学植物保护学院提供.试验接种于叶片背面二级叶脉之间,剂量为100 μL·片-1,覆盖面积约为20 mm2.在接种菌完成之后,将样本放入人工气候箱中,在温度为18 ℃,相对湿度为100%,每天16 h光照和8 h黑暗培养的条件下,进行为期6 d的连续培养.
将100片叶片分为5组,每组20片;采集完高光谱信息后,再测定POD酶活性.首先对第1组的20片马铃薯叶片进行了第1次样本采集;24 h后擦去剩余4组接种晚疫病菌样本的残留液滴,以防止交叉感染;同时对第2组的20片叶片进行第2次数据采集.之后的数据每隔24 h采集一次,共采集样本数据100组.
1.3 模型建立
1.3.1 光谱数据预处理
将获得的高光谱图像用软件ENVI4.8进行处理.选择病斑部位作为感兴趣区域(region of inte-rest, ROI),对未发病叶片也选取其相应的位置作为感兴趣区域.计算出所有感兴趣区域在全光谱下像素点的光谱反射率平均值,留待后续的处理,即将原始高光谱数据转化成为感兴趣部分的全光谱信息.
为了提高信噪比,以保证所建模型的有效和稳健性,应对提取出来的平均光谱曲线进行预处理.文中所采取的预处理方法有平滑(savitzky-golay smoo-thing, SG)、变量标准化(standard normal variate, SNV)、多元散射校正(multiplicative scatter correction, MSC)、二阶求导(2nd derivative, 2-Der).用于建模分析的光谱数据都是经过以上预处理获得的.
1.3.2 化学计量学建模方法
偏最小二乘回归法(partial least square, PLS)是一种建立线性回归模型的方法,可以反映出潜在变量.该方法常用来间接反映、解释变量,或者反映不同变量在一定条件下的关系.因此,此方法适用于处理多重共线性严重的数据,特别是变量多、样本量少的情况[8].但它无法将非线性特征纳入到预测模型中,为了提高模型精度,试验进一步采用最小二乘支持向量机(least squares-support vector machine,LS-SVM)法来建立模型.LS-SVM模型以线性方程代替二次规划问题解决线性和非线性多元回归问题,能够大幅度地简化运算,加快求解速度[9].
在光谱分析中,最基本、最常应用的处理方法就是多元线性回归(multiple linear regression, MLR)[10].这种方法用多个波长的光谱数据作为说明函数,将光谱中包含的有用信息集成起来,建立用来描述相关目的品质和光谱数据之间关系的模型.
校正集相关系数RC及其均方根误差RMSEC、预测集相关系数RP及其均方根误差RMSEP常用于评价模型的好坏,其中,RMSEP值小而RP值大的模型预测效果较好[11].
1.3.3 特征波长的优选
在使用全光谱进行建模时,有些波段数据之间存在的相关性小,所包含有效信息较少,除了会增大计算量,也严重影响到预测模型,造成模型精度低和稳定性差的后果.因此,采用连续投影算法(successive projection algorithm, SPA)提取出对被测成分的影响较大的几个或者十几个特征波长,可在信息最少化前提下保证模型准确度,提高建模效率.
用PLS进行建模分析时,可通过每个隐含变量下各个波长点获得所对应的载荷系数.载荷系数绝对值的大小能够反映出该波长对应下所建模型预测性能的好坏.因此,选取特征波长时,可以依据某一隐含变量下各个波长所对应的载荷系数绝对值大小(即载荷系数法x-loading weight, X-LW)来判断[12].利用提取的特征波长建立POD酶活性的预测模型,这样能够大幅度减少运算量,实现简化模型的目的.
2 结果与讨论
2.1 叶片染病后的POD活性变化
将100个样本数据按照3 ∶1的比例划分为建模集75个,预测集25个(每天20个样本中取15个数据作为建模集,其余5个作为预测集,共5 d的数据).POD活性的统计信息见表1,其中A为POD的酶活性值.图2为不同感染时间下POD酶活性的平均变化趋势,其中t为累计染病时间.

表1 POD酶活性值的统计信息
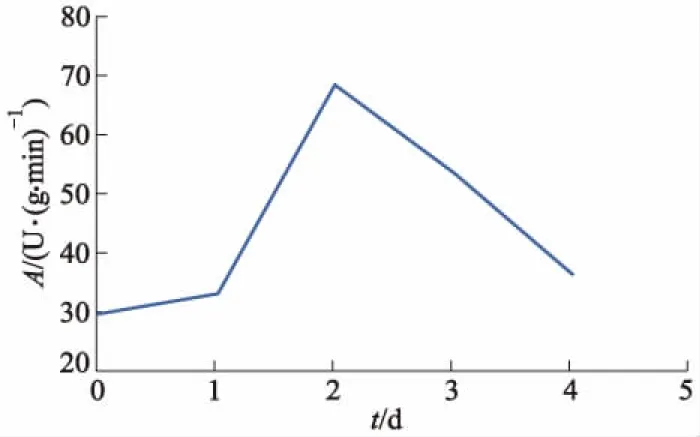
图2 不同感染时间下POD酶活性的平均变化趋势
由图2可见,马铃薯叶片在初期感染晚疫病的12 h内酶活性上升缓慢,这意味着该马铃薯品种植株的抗病性不强.染病24~48 h,酶活性上升迅速,并达到最大,此时叶片处于被晚疫病强烈侵害的应激状态,POD活性增强;之后,由于叶片组织细胞壁不断遭到破坏,酶活性随之降低.
2.2 高光谱反射曲线分析
图3显示了在染病的不同阶段中,马铃薯病叶的高光谱反射率曲线随时间的变化情况.图3中,λ为波长,Rr为光谱反射率.由于提取到的光谱数据在<400 nm和>1 000 nm的波长存在严重的噪声干扰, 因此仅采用400~1 000 nm的波长范围进行了分析.
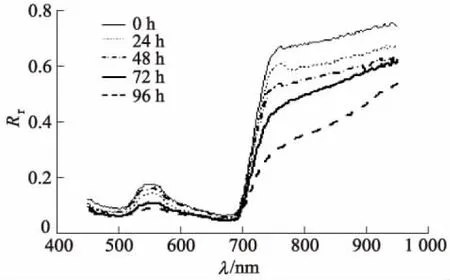
图3 不同患病时间的平均光谱曲线
由图3可知,发病叶片的光谱反射率都会随着染病时间的增加而呈现出减小的趋势,即健康叶片往往呈现出最高的光谱反射率,染病程度较轻的叶片较弱,染病程度最高的叶片光谱反射率最低.具体表现在:马铃薯叶片从健康状态发展到感染病害且病情不断加深时,叶片中叶绿素含量逐渐减少,对蓝光(λ=490~500 nm)、绿光(λ=500~560 nm) 的吸收增强,而反射减弱.在近红外区域(λ>780 nm),晚疫病破坏马铃薯叶片组织细胞,光合作用无法合成产物,原有物质不断分解,使得叶片在病害严重时的光谱反射率进一步下降.这表明马铃薯晚疫病叶片在不同病害程度时的光谱反射曲线变化有着一定的规律性,并且遵循病叶的生理变化规律.
2.3 高光谱数据预处理和基于全光谱的建模分析
表2给出了不同预处理方法对PLS模型的预测结果.
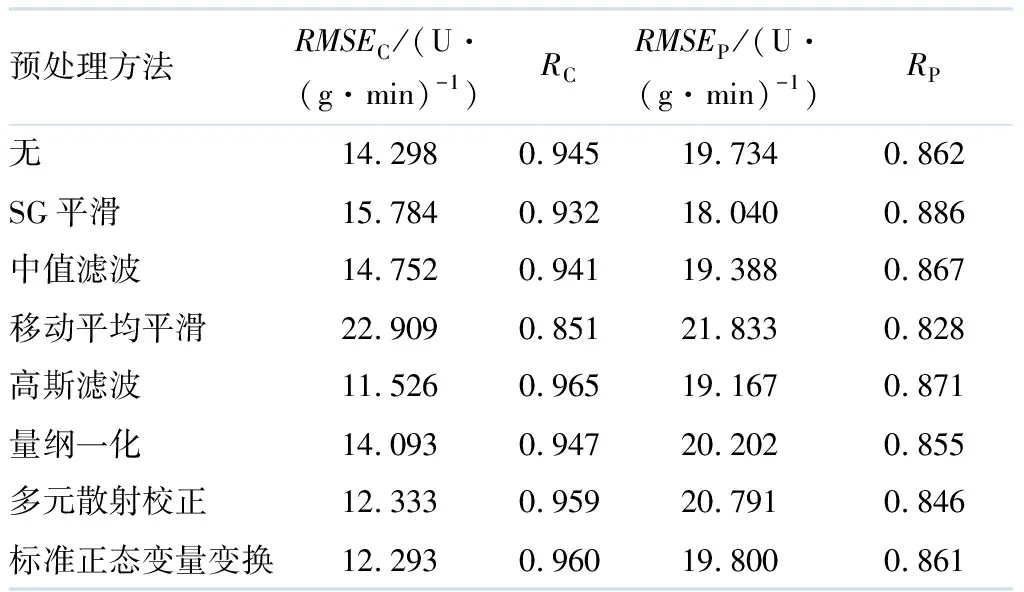
表2 不同预处理方法对PLS模型的预测结果
由表2可见:经过高斯滤波预处理后,建模集的效果最好;SG平滑与中值滤波预处理后,对预测集的效果相当;经过中值滤波预处理后,建模集和预测集的均方根误差相差最小,模型稳定度最高,预测效果最好.综上所述,选择中值滤波作为最优预处理方法,文中所有光谱数据都使用该方法进行预处理.
同时,经过中值滤波法预处理后的光谱数据作为输入值,POD的活性作为输出值,建立基于全光谱的LS-SVM模型.该模型的预测结果见表3.

表3 模型预测结果的对比
由表3可见,与使用PLS法所建立的模型相比,使用LS-SVM法建立的模型建模集RC值较小,而其预测集RP值大、RMSEP值较小,所以使用LS-SVM法建立的模型预测能力强.综上,在全光谱范围之内,使用LS-SVM法建的模型更适用对POD活性的预测.由表3可知,LS-SVM的RC值和RP值都在0.9以上,说明拟合效果较好.
2.4 特征波长建模分析
2.4.1 基于SPA算法提取特征波长的预测模型
试验中设定最小和最大选定波段数分别为1和30,计算过程通过Matlab软件实现.计算结果见图4,5,其中N为特征波长的个数,X为x载荷系数.最佳特征波长个数的确定以建模集的RMSE趋于稳定且达到最小时所对应的波长数为依据.
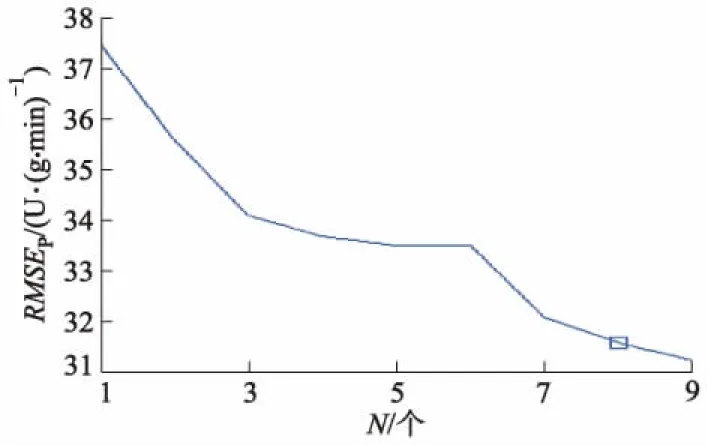
图4 RMSEP值随有效波长数目的变化趋势
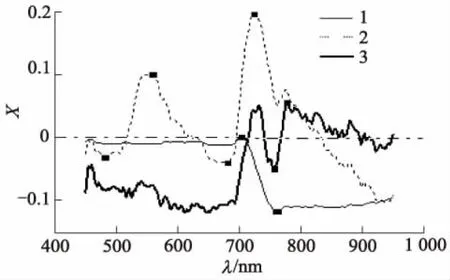
图5 前3个LV的x载荷系数
图4展示了均方根误差随特征波长增加的变化趋势,由图4可确定出特征波长数目为8个.图5显示了特征波长所在的波段序号,其编号分别为1,6,20,39,93,101,128,199,所对应的波长分别为450,462,496,543,677,697,766,949 nm.
把以上8个特征波长对应的光谱数据作为输入,分别建立基于PLS,MLR法的线性模型和基于LS-SVM法的非线性模型,结果见表4.
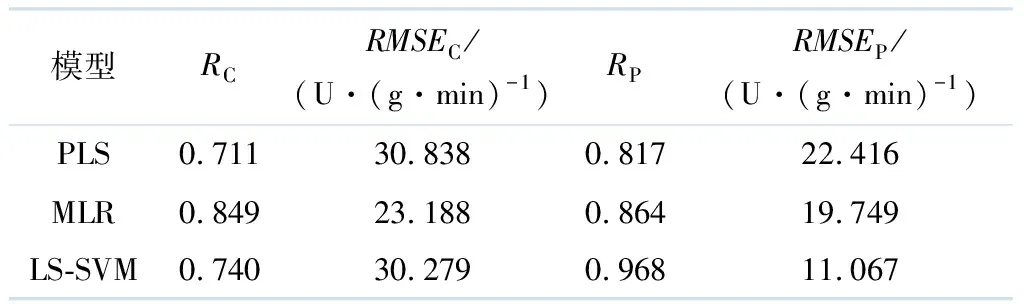
表4 不同预测方法的SPA模型的结果
由表4可见,使用SPA算法提取特征波长后,基于非线性LS-SVM模型预测效果较基于线性PLS,MLR模型的好.在2种线性模型中,显然SPA-MLR的模型预测效果更好,预测的表达式如下:
A=67.322-3 191X1-3 083X2+
3 841X3-535.571X4-7 451X5+
3 238X6+35.224X7-137.391X8,
(2)
式中X1-X8依次为450,462,496,543,677,697,766,949 nm这8个特征波长下的光谱反射率.
2.4.2 基于X-LW法提取特征波长的预测模型
在Unscrambler环境中建立全光谱的PLS模型时,把推荐的前3个隐含变量(LVs)作为已知参数.图5显示了每个隐含变量在各个波长点处的载荷系数.特征波长就是局部载荷系数在绝对值最大处对应的波长点.由图5可得到10个特征波长,分别为481,557,560,680,702,722,755,758,761,776 nm.把使用X-LW法提取到的10个特征波长所对应的光谱数据作为输入值,分别建立基于PLS,MLR的线性模型以及基于LS-SVM的非线性模型.表5给出了X-LW方法对不同模型的预测结果.
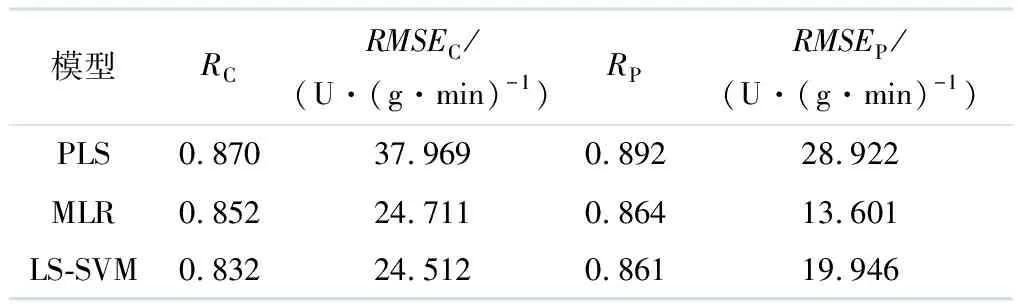
表5 X-LW方法对不同模型的预测结果
由表5可见,基于X-LW线性模型法提取特征波长后,使用线性模型PLS,MLR模型预测效果比使用非线性LS-SVM法所建的模型好.其中,X-LW-PLS模型的RC和RP均为3者中最高,预测效果最佳.但3种模型的相关系数都低于0.9,可以进一步优化模型以达到更好的预测效果.
综上所述,在使用特征波长所建立的预测模型中,SPA-LS-SVM模型的预测集RP值最大,但与建模集RC相差较大,不利于建模的稳定性.因此,选择X-LW-PLS作为最优预测模型.
3 结 论
1) 基于全波长的预测模型中,LS-SVM模型预测效果最优,预测集RP值为0.932,RMSEP值为14.966 U·(g·min)-1.利用SPA与X-LW这2种方法进行择特征波长的选择,得到的X-LW-PLS模型,预测效果最优,预测集RP值是0.892,RMSEP值是28.922 U·(g·min)-1.
2) POD酶活性的变化并不是随着染病时间的增加而单调递增或是递减,因此仅仅凭借POD酶活性是无法对马铃薯的染病时间进行判断的,还需结合病害的其他症状.
3) 根据POD酶活性的变化可以判断叶片是否处于染病状态,为及时发现病害并进行早期防治提供了参考.
4) 高光谱成像技术能够用来实现马铃薯晚疫病叶片中过氧化物酶POD酶活性的快速无损测定,并证实了POD酶活性实现晚疫病的早期实时检测的可行性.
