静宁鸡肌肉生长抑制素基因单核苷酸多态性筛查及生物信息学分析
2018-11-10张丽刘丽霞戴洪伟陈红王瑞岳炳辉2
张丽,刘丽霞,戴洪伟,陈红,王瑞,岳炳辉2
(1.西北民族大学生命科学与工程学院,兰州730030;2.青海畜牧兽医职业技术学院,青海湟源812100)
肌肉生长抑制素基因(myostatin,MSTN)属于转化生长因子-β(transforming growth factor-β,TGF-β)超家族成员之一,不仅对肌肉生长有负调控作用,还对脂肪的沉积和骨骼的生长发育具有调控作用,甚至还能影响肌腱和韧带的强硬度。研究表明,该基因的突变与缺失可使家畜表现出肌肉超级发达的“双肌性状”[1]。比利时蓝牛和皮尔蒙特牛的“双肌”现象就是由MSTN基因第1外显子和第3外显子上的2个碱基突变造成蛋白活性区的移码突变,从而不能形成成熟的MSTN蛋白,导致体质量增加而脂肪质量不增加[2]。敲除MSTN基因的小鼠除肌肉质量增加外,脂肪含量也随年龄的增长而显著降低[3]。
静宁鸡又叫静原鸡、固原鸡,是适应黄土高原寒冷干旱生态条件的优良肉蛋兼用型的地方鸡种,因主产于甘肃省静宁县而得名。静宁鸡适应性强,体质结实,体形较大,觅食能力强,蛋质量较大,肉质好,具有独特的地方风味,深受广大人民的喜爱。但随着近年来主产区人口外迁和引进品种的冲击,静宁鸡饲养量日益下降,分布范围逐步缩小,加之较严重的近亲繁殖,其特色基因丢失,种质资源异化,优良的静宁鸡已处于濒危状态,也使其分子方面的研究受到极大限制。目前仅见安锋利[4]和李显耀等[5]关于静宁鸡分子遗传方面的研究,而有关静宁鸡MSTN基因多态性和蛋白结构的研究还未见报道。因此,如果能在静宁鸡群体中筛选出MSTN基因突变个体,则可通过育种培育出产肉率较高的优良家禽品种,拯救静宁鸡遗传资源濒危问题。
为深入探讨静宁鸡MSTN基因编码蛋白的性质和功能,避免人工选育和基因演化造成的差异,本研究利用生物信息学方法对静宁鸡MSTN基因序列结构及相关生物信息学特征进行预测和分析,为揭示其理化性质、功能信息、遗传特性、相关生理机制及与生长发育和肉质性状的关系提供理论基础。
1 材料与方法
1.1 血样采集与DNA提取
于宁夏彭阳县静原鸡繁育中心采集第5世代静宁鸡血样152份,用医用采血管保存。采用传统的苯酚-氯仿抽提法提取样品的基因组DNA,用去离子水溶解稀释。取DNA母液各5µL构建DNA混合池,于-20℃冰箱中冷冻保存,备用。
1.2 引物设计
参考Ensembl中原鸡MSTN基因序列(ENS GALG00000023961),利用Primer 5.0和Primer Select软件设计3对特异性引物序列(表1),由生工生物工程(上海)股份有限公司合成。
1.3 聚合酶链式反应扩增和测序
聚合酶链式反应(polymerase chain reaction,PCR)扩增体系(40µL)为:DNA模板2.0µL,0.25µmol/L 上 下 游 引 物 各 1.0 µL,2×Taq PCR MasterMix 22.0µL,双蒸水14µL。PCR扩增程序为:95℃预变性4 min;95℃变性30 s,60℃退火30 s,72 ℃延伸30 s,35个循环;72 ℃延伸10 min;4℃保存。然后用1%琼脂糖凝胶电泳检测扩增结果。PCR产物经凝胶回收试剂盒纯化回收后直接送至北京华大基因有限公司进行测序,测序结果用DNAStar、MegAlign和Editseq软件进行比对和拼接,获得静宁鸡MSTN基因编码区序列。

表1 静宁鸡MSTN基因引物信息Table 1 Primer information of MSTN gene in Jingning chicken
1.4 等位基因频率的估算
利用Bioedit软件查看测序结果,运用MWSnap软件测量单核苷酸多态性(single nucleotide polymorphism,SNP)位点等位基因峰高,并依据公式对各等位基因频率进行估算[6]。其中,Fi表示SNP位点某等位基因频率,h1和h2分别表示测序图上该SNP位点等位基因1和2的峰高。
1.5 生物信息学分析工具
按照文献[7-8]中介绍的在线软件对静宁鸡MSTN基因编码区进行生物信息学分析。
2 结果与分析
2.1 静宁鸡MSTN基因PCR分段扩增结果
静宁鸡MSTN基因PCR分段扩增检测结果如图1所示。从中可知,扩增产物电泳条带清晰,特异性良好,片段大小与预期扩增片段长度相符。
2.2 静宁鸡MSTN基因序列测定
将拼接的静宁鸡MSTN基因与Ensembl中公布的原鸡MSTN基因CDS区序列进行比对,共发现4个SNPs(图2),其中在第1外显子发现3个SNPs,分别为c.60G>A、c.195C>G、c.234G>A,第3外显子仅存在一个c.966G>T突变。由于突变位点核苷酸的密码子简并现象,未引起编码氨基酸的改变,均属同义突变。

图1 静宁鸡MSTN基因PCR扩增产物检测结果Fig.1 PCR amplified products of MSTN gene in Jingning chicken
2.3 SNPs位点等位基因频率估算
由表2可知,静宁鸡MSTN基因突变位点的等位基因频率在突变前后存在差异,其中c.195C>G和c.966G>T突变前后差异较大,其余2个位点突变前后等位基因频率差异较小。
2.4 生物信息学分析
2.4.1 静宁鸡MSTN蛋白理化性质分析

图2 静宁鸡MSTN基因测序结果Fig.2 Results of MSTN gene sequencing in Jingning chicken
获得的静宁鸡MSTN基因编码区全长1 128 bp,含一个完整的开放阅读框(open reading frame,ORF),包含起始密码子和终止密码子,编码375个氨基酸。从图3中可以看出:亮氨酸(Leu)数目最多(30个),占整个氨基酸组成的8.0%;组氨酸(His)数目最少(5个),占整个氨基酸组成的1.3%。正电荷残基总数(Arg+Lys)和负电荷残基总数(Asp+Glu)均为46个。带正电荷氨基酸51个,酸性氨基酸81个,碱性氨基酸46个,极性氨基酸224个,疏水性氨基酸105个。对蛋白质空间结构起关键作用的半胱氨酸(Cys)为13个;α-螺旋终止者脯氨酸(Pro)为25个。蛋白质分子式为C1905H2988N510O561S22,分子质量约为42.7 kDa,理论等电点(pI)约为6.93,半衰期为30 h,不稳定指数为44.19。
2.4.2 静宁鸡MSTN蛋白亲水/疏水性预测
静宁鸡MSTN蛋白亲水性/疏水性预测如图4所示。第173位的异亮氨酸(Ile)疏水性最强(2.200),第263位的精氨酸(Arg)亲水性最强(-3.056)。在整条氨基酸肽链中,亲水氨基酸占63.47%,疏水氨基酸占36.53%,总平均亲水性为-0.399,总体表现为亲水性。由此可知,静宁鸡MSTN蛋白是一种亲水性蛋白[9]。

图3 静宁鸡MSTN基因编码蛋白氨基酸组成Fig.3 Amino acid composition of MSTN gene-encoded protein in Jingning chicken
2.4.3 SNPs对静宁鸡MSTN基因mRNA二级结构的影响
对外显子1和3处的突变位点c.60G>A、c.195C>G、c.234G>A、c.966G>T进行mRNA二级结构预测分析的结果表明,SNPs突变前后引起mRNA二级结构改变,并导致mRNA二级结构的最小自由能发生改变(图5)。其中:c.60G>A位点突变前后的自由能未发生变化;c.195C>G位点突变前后的自由能由-371.30 kJ/mol变为-374.23 kJ/mol;c.234G>A位点突变前后的自由能由-363.34 kJ/mol变为-367.95 kJ/mol;c.966G>T位点突变前后的自由能由-409.81 kJ/mol变为-398.93 kJ/mol。
2.4.4 静宁鸡MSTN蛋白的二级与三级结构预测
蛋白质二级结构主要指多肽链主链骨架依赖氢键排列成在一维方向上具有周期性结构的构象。使用SOPMA软件预测静宁鸡MSTN蛋白二级结构的结果表明:169个氨基酸构成无规则卷曲,占总氨基酸的45.07%,99个氨基酸构成延伸链,占总氨基酸的26.40%;另外81个氨基酸和26个氨基酸分别形成α-螺旋和β-转角,分别占总氨基酸的21.60%和6.93%。采用SWISS-MODEL对静宁鸡MSTN蛋白同源建模获得三级结构模型(图6),结果表明,静宁鸡MSTN蛋白主要由无规则卷曲和延伸链构成,这与二级结构预测结果一致。

图4 静宁鸡MSTN基因编码蛋白质的亲水疏水性预测结果Fig.4 Hydrophilic/hydrophobic predictions of MSTN geneencoded protein in Jingning chicken
3 讨论
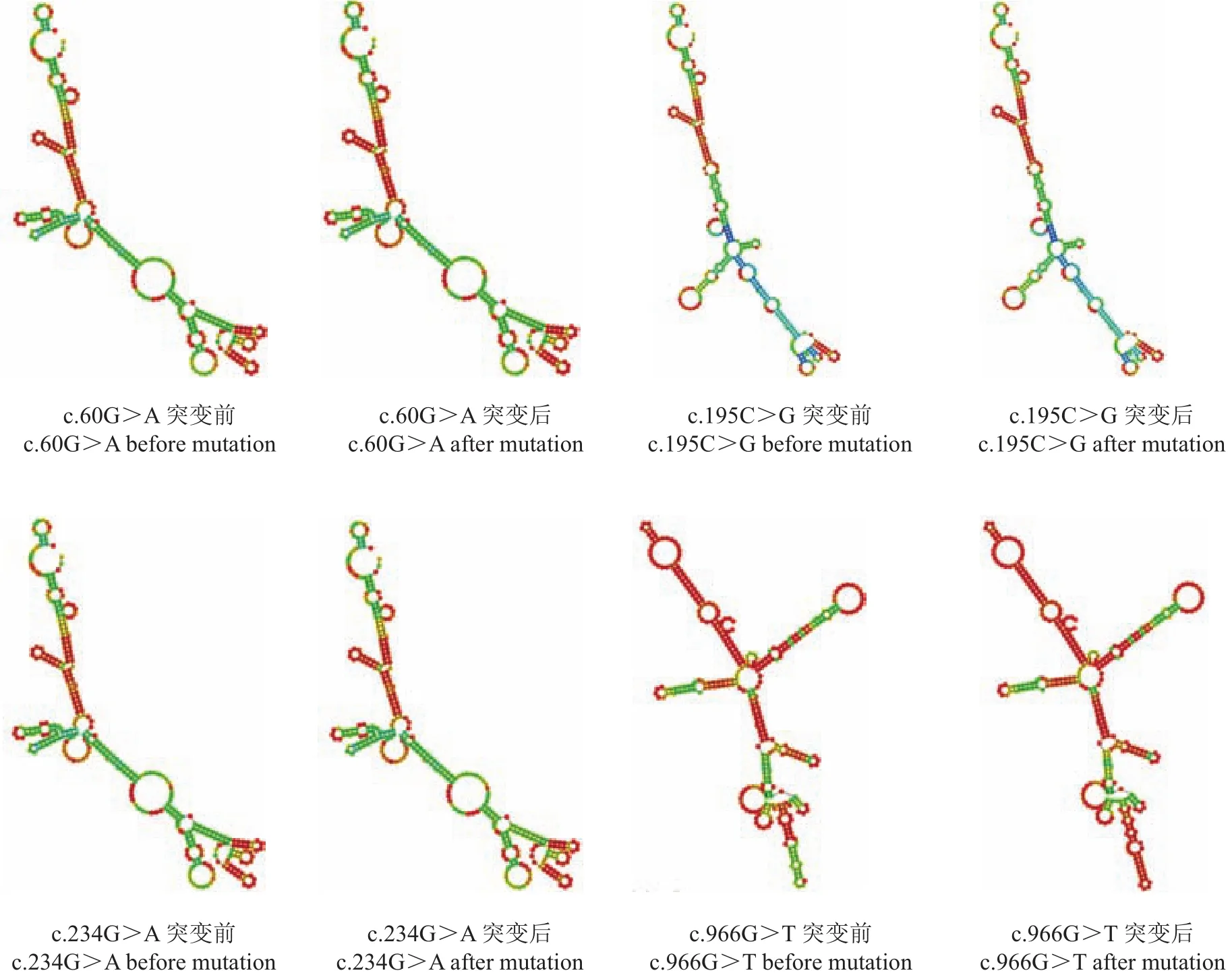
图5 静宁鸡MSTN基因mRNA二级结构Fig.5 Secondary structure of mRNAof MSTN gene in Jingning chicken

图6 静宁鸡MSTN蛋白三级结构预测Fig.6 Putative tertiary structure of MSTN protein in Jingning chicken
大量研究显示,MSTN基因突变已导致人[10]、牛[11]、羊[12]、猪[13]和狗[14]表现出“双肌性状”。MSTN基因突变的纯合体及杂合体与肉牛骨骼肌生长发育有关,使肉牛表现出肌肉超级发达、初生质量增加和生长速度快的优势[11]。在惠比特犬中存在1个可引起双肌表型自然发生的突变c.939-940delTG,其编码区第939~940 bp处缺失2个碱基T和G,从而引起第313位半胱氨酸转变为终止密码子;该纯合突变具有双肌表型,主要表现在宽阔的胸膛和发育超强的腿及颈部肌肉;杂合子的肌肉质量比野生型稍大,但表现出优异的竞速优势[14]。在绵羊中也发现了3个自然发生的突变可引起双肌效应,其中c.960delG是编码区第960位碱基G缺失,造成蛋白质翻译移码和终止子的提前生成,致使挪威白绵羊出现明显的双肌臀,且内脏脂肪和皮下脂肪含量偏低;c.120insA是编码区第120位插入了碱基A,造成古挪威绵羊蛋白质翻译移码和终止子的提前生成;c.2360G>A是MSTN基因3’UTR区发生的突变,虽没有改变MSTN蛋白的功能,但能引起骨骼肌MSTN蛋白的高度表达,并引起MSTN的转录抑制效应,造成血液中MSTN蛋白含量大幅下降,同时表现出双肌性状,这一突变常见于夏洛来羊、白萨福克羊和无角道赛特羊等[15-17]中。MSTN基因也可能是影响家禽骨骼肌生长的主效基因或与控制该性状的主效基因连锁,能够作为候选基因用于提高地方鸡产肉率的分子标记辅助选择。通过抑制MSTN活性来增加肌肉量,在畜禽分子育种上具有潜在的应用价值。
MSTN基因在脊椎动物间高度保守。MCPHGRRON等[18]研究表明,人、鸡、鼠、猫的MSTN蛋白在C-端同源性高达100%,而在牛、羊中只有1~3个氨基酸表现为不同,石斑鱼MSTN的保守性较差,与其他动物羧基端同源性为88%。MSTN基因含有3个外显子和2个内含子,不同物种间的MSTN基因长度有所不同。本研究获得的静宁鸡MSTN基因编码区全长1 128 bp,编码375个氨基酸,第1、2、3外显子大小分别为373、374、381 bp,与藏鸡[19]、三穗鸭[20]完全一致,但与猪[21]不同,符合同源基因的进化关系。
MSTN基因在动物进化过程中高度保守,仅有若干个单核苷酸的同义突变或错义突变。将静宁鸡和原鸡MSTN基因编码核苷酸序列进行比对发现:静宁鸡MSTN基因第1外显子变异最大,存在3处突变(c.60G>A、c.195C>G和c.234G>A);第3外显子变异最小,存在1处突变(c.966G>T);第2外显子未发现多态。这可能是因为MSTN蛋白功能区主要集中在第3外显子编码的氨基酸序列,其中9个保守性的半胱氨酸对该基因发挥着主要功能,其改变可能导致静宁鸡MSTN基因功能发生改变。与温彦涛等[22]和赵振华等[23]的研究结果相比,c.60G>A和c.966G>T是新发现的2个突变位点。4个SNPs位点均未引起编码氨基酸变异,属同义突变。这可能是由于MSTN基因序列和结构的高度保守性,以及密码子的简并性使相应的蛋白质未发生改变[24]。毛亮等[19]研究发现,藏鸡与原鸡氨基酸序列同源性高达99.7%,有6个同义突变和1个错义突变。左斌等[25]在泸宁鸡和米易鸡群体中发现第1外显子存在2个同义突变,其中g.2100C>T与鸡心脏质量、屠宰率和胫围显著相关;BARON等[26]和YE等[27]在肉鸡和蛋鸡群体中发现MSTN基因高度多态,其外显子1存在7个SNPs,外显子2存在7个SNPs和1个缺失,外显子3存在1个突变位点。朱智等[28]发现MSTN基因第1外显子的g.2100G>A和g.2109G>A突变与温岭草鸡的屠宰率、腹脂率和胸肌率显著相关。起源、进化和选育程度的不同,导致静宁鸡与其他家禽品种在MSTN基因核苷酸序列上的差异,但在氨基酸水平上无差异,说明其在核苷酸水平上的差异不影响编码蛋白的结构组成,这将为从DNA水平上研究品种间起源和进化关系提供一定的理论基础。MSTN基因在同一物种不同品种间表现出的碱基突变有可能影响蛋白质的空间结构,从而影响其功能,说明该基因具有作为遗传标记的潜在可能。
基因决定生物性状最终是通过蛋白质来体现的。蛋白质理化性质预测结果显示:静宁鸡MSTN蛋白由375个氨基酸折叠而成;总平均亲水性为-0.399,依据氨基酸分值越高疏水性越强,分值越低亲水性越强的规律,可见该蛋白为亲水性蛋白;不稳定指数值大于40,表明该编码蛋白是不稳定蛋白[29]。贾浩等[30]报道了蛋白质半衰期越长则稳定性越高,而本研究发现静宁鸡MSTN具有较长的半衰期(30 h)却是不稳定蛋白,这可能与MSTN蛋白不同生理功能的发挥存在某种联系。在20种氨基酸组成中,营养价值较高的亮氨酸(Leu)数目最多(30个),比鸭的少3个,比牛、羊少5~6个;弱碱性的组氨酸(His)数目最少(5个),比羊的多2个。生理条件下是否带电与周围内环境有关。对蛋白质空间结构起关键作用的半胱氨酸(Cys)为13个,与鸭、牛、羊的数目相同,说明各种动物MSTN蛋白中的半胱氨酸高度保守;已证实,第3外显子的9个保守性半胱氨酸残基就是MSTN基因发挥正常生物学功能并行使肌肉负调控作用的关键。终止α-螺旋的脯氨酸(Pro)为25个,比鸭、羊少1个。通过比较发现,各种动物MSTN基因编码区序列中性质相似的氨基酸数目和在序列中的位置都有不同程度的差异,表明该基因在动物进化过程中存在多态性。这与易恒洁等[20]关于三穗鸭的预测结果基本一致。
在外显子中构成编码区的核苷酸突变可能引起mRNA二级结构的改变,从而引起蛋白质结构改变而影响其生物学功能。对静宁鸡MSTN基因的mRNA二级结构预测的结果表明:突变前后mRNA二级结构和自由能的改变均会影响其结构的稳定性,c.195C>G和c.234G>A突变前后的自由能分别降低了2.93和4.61 kJ/mol,结构稳定性的提高使蛋白质二级结构也发生改变;c.966G>T突变前后的自由能增加了10.88 kJ/mol,稳定性降低,蛋白质二级结构改变;c.60G>A突变前后的自由能未发生变化。
蛋白质二级结构是多肽链依赖氢键排列在一维方向上具有周期性结构的构象,对其进行预测和分析有助于了解其三维构象和蛋白质的空间结构。本研究中静宁鸡MSTN二级结构包含169个氨基酸的无规则卷曲、99个氨基酸的延伸链、81个氨基酸的α-螺旋和26个氨基酸的β-转角,不存在β-折叠。按照α-螺旋比例大于45%,同时β-折叠比例小于5%的蛋白质二级结构分型标准,静宁鸡MSTN蛋白二级结构属于非混合型蛋白,这种结构是否影响该蛋白的能量代谢、运输和结合等过程还有待进一步探讨,但这一研究结果与鸭、牛、猪完全不同。易恒洁等[20]认为三穗鸭MSTN蛋白二级结构包含22个α-螺旋、26个β-折叠、26个β-转角和20个无规则卷曲;杜晓华等[31]的研究表明,西门塔尔牛MSTN蛋白质二级结构是以无规则卷曲和β-转角为主的混合型蛋白;王伟等[32]的研究表明,猪MSTN蛋白二级结构含有20.53%的α-螺旋、4%的β-转角、53.07%的无规则卷曲和22.4%伸展条的混合型蛋白。可见,静宁鸡MSTN蛋白二级结构最大元件是无规则卷曲,延伸链、α-螺旋和β-转角则散布于整个蛋白质结构中。
4 结论
由于肌肉生长抑制素基因能够调控动物肌肉生长,使人类获得更多的动物肉产品,所以其在畜牧生产中具有广泛的应用前景。本研究对静宁鸡MSTN基因的理化性质和蛋白结构功能进行了系统预测,以期为揭示静宁鸡的肉质性状提供理论基础,同时为分子遗传育种技术应用于生产实践提供依据。
参考文献(References):
[1] 杨永生,贺建华,邓惠中,等.肌肉生长抑制素对动物肌肉、脂肪和骨骼的影响.动物营养学报,2012,24(2):220-225.YANG Y S,HE J H,DENG H Z,et al.Myostain:Effects on muscle,fat and bone of animals.Chinese Journal of Animal Nutrition,2012,24(2):220-225.(in Chinese with English Abstract)
[2]KAMBADUR R,SHARMA M,SMITH T P L,et al.Mutationsin in myostatin(GDF8)in double-muscled Belgian Blue and Piedmontese cattle.Genome Research,1997,7:910-915.
[3] MCPHERRON A C,LEE S J.Suppression of body fat accumulation in myostatin-deficient mice.The Journal of Clinical Investigation,2002,109(5):595-601.
[4] 安锋利.静宁鸡肌肉发育调控机理.兰州:兰州大学,2013:57-81.AN F L.Mechanism of muscle developmental regulation in Jingning chicken.Lanzhou:Lanzhou University,2013:57-81.(in Chinese with English Abstract)
[5] 李显耀,张龙超,曲鲁江,等.静宁鸡和固原鸡品种划分的分子遗传基础.农业生物技术学报,2005,13(5):664-667.LI X Y,ZHANG L C,QU L J,et al.Molecular genetic basis of identifying Jingning and Guyuan chickens.Journal of Agricultural Biotechnology,2005,13(5):664-667.(in Chinese with English Abstract)
[6]SULIMAN H B,CARRAWAY M S,PIANTADOSI C A.Postlipopolysaccharide oxidative damage of mitochondrial DNA.American Journal of Respiratory and Critical Care Medicine,2003,167(4):570-579.
[7] 张丽,刘丽霞,李强子,等.天祝白牦牛MSTN基因编码区克隆及生物信息学分析.浙江农业学报,2017,29(4):618-624.ZHANG L,LIU L X,LI Q Z,et al.Cloning and bioinformatics analysis of MSTN gene of Tianzhu white yak.Acta Agriculturae Zhejiangensis,2017,29(4):618-624.(in Chinese with English Abstract)
[8] 盘道兴,王振,杨茂林,等.江口萝卜猪FABPs主要家族基因SNPs筛选及生物信息学分析.基因组学与应用生物学,2015,34(1):47-52.PAN D X,WANG Z,YANG M L,et al.SNPs screening and bioinformatics analysis of the main family genes of FABPs in Jiangkou Luobo pig.Genomics and Applied Biology,2015,34(1):47-52.(in Chinese with English Abstract)
[9] KYTE J,DOOLITTLE R F.A simple method for displaying the hydropathic character of a protein.Journal of Molecular Biology,1982,157:105-132.
[10]SCHUELKE M,WAGNER K R,STOLZ L E,et al.Myostatin mutation associated with gross muscle hypertrophy in a child.The New England Journal of Medicine,2004,351(26):2682-2688.
[11]GROBET L,MARTIN L J R,PONCELET D,et al.A deletion in the bovine myostatin gene causes the doublemuscled phenotype in cattle.Nature Genetics,1997,17(1):71-74.
[12]CLOPA,MARCP F,TAKEDA H,et al.A mutation creating a potentialillegitimate microRNA targetsite in the myostatin gene affectsmuscularity in sheep.Nature Genetics,2006,38(7):813-818.
[13]STINCKENS A,LUYTEN T,BIJTTEBIER J,et al.Characterization of the complete porcine MSTN gene and expression levels in pig breeds differing in muscularity.Animal Genetics,2008,39(6):586-596.
[14]MOSHER D S,QUIGNON P,BUSTAMANTE C D,et al.A mutation in the myostatin gene increases muscle mass and enhances racing performance in heterozygote dogs.PLoS Genetics,2007,3(5):245-263.
[15]BOMAN I A,KLEMETSDAL G,BICHFELDT T,et al.A frameshift mutation in the coding region of the myostatin gene(MSTN)affects carcass conformation and fatness in Norwegian White Sheep(Ovis aries).Animal Genetics,2009,40(4):418-422.
[16]BOMAN I A,VAGE D I.An insertion in the coding region of the myostatin(MSTN)gene affects carcass conformation and fatness in the Norwegian Spaelsau(Ovis aries).BMC Research Notes,2009,2:98.
[17]BOMAN I A,KLEMETSDAL G,NAFSTAD O,et al.Impact of two myostatin(MSTN)mutations on weight gain and lamb carcass classification in Norwegian White Sheep(Ovis aries).Genetics Selection Evolution,2010,42(1):4.
[18]MCPHERRON A C,LEE S J.Double muscling in cattle due to mutations in the myostatin gene.Proceedings of the National Academy of Sciences of the USA,1997,94(23):12457-12461.
[19]毛亮,徐亚欧,麻丽霞.藏鸡肌肉生长抑制素基因克隆及序列分析.西南民族大学学报(自然科学版),2009,35(4):772-775.MAO L,XU Y O,MA L X.Study on cloning of the MSTN gene in the Tibetan gallus.Journal of Southwest University for Nationalities(Natural Science Edition),2009,35(4):772-775.(in Chinese with English Abstract)
[20]易恒洁,李辉,杨胜林,等.三穗鸭肌肉生长抑制素基因的生物信息学分析.中国畜牧兽医,2014,41(4):1-6.YI H J,LI H,YANG S L,et al.Bioinformatics analysis of MSTN gene in Sansui duck.China Animal Husbandry&Veterinary Medicine,2014,41(4):1-6.(in Chinese with English Abstract)
[21]乔宪凤,刘西梅,华文君,等.猪肌生成抑制素(MSTN)基因cDNA克隆及序列分析.江西农业学报,2010,22(10):130-132.QIAO X F,LIU X M,HUAW J,et al.Cloning and sequencing of Hubei white swine myostatin gene cDNA.Acta Agriculturae Jiangxi,2010,22(10):130-132.(in Chinese with English Abstract)
[22]温彦涛,武子寅,赵振华,等.鸡MSTN基因多态性及其与屠体性状的关联分析.中国家禽,2012,34(16):29-32.WEN YT,WU Z Y,ZHAO Z H,et al.Polymorphisms in exon 1 of myostatin gene and its relationship with carcass traits in chicken.China Poultry,2012,34(16):29-32.(in Chinese with English Abstract)
[23]赵振华,黎寿丰,黄华云,等.MSTN基因突变对优质鸡肌肉生长的影响.安徽农业大学学报,2015,42(5):733-737.ZHAO Z H,LI S F,HUANG H Y,et al.The myostatin gene(MSTN)and its relationship with muscle fiber traits in chickens.Journal of Anhui Agricultural University,2015,42(5):733-737.(in Chinese with English Abstract)
[24]孙涛.基因突变对蛋白质影响的几种不同情况.中学生物教学,2011(7):41-42.SUN T.Several different circumstances of gene mutations influencing the protein on the different circumstances.Middle School Biology Teaching,2011(7):41-42.(in Chinese)
[25]左斌,任冰冰,蒋小松,等.泸宁鸡和米易鸡MSTN基因多态性及其与生长性状的关联性.贵州农业科学,2015,43(7):26-30.ZUO B,REN B B,JIANG X S,et al.Polymorphism of myostatin(MSTN)gene and correlation between MSTN polymorphism and growth traits in Luning and Miyi chicken.Guizhou Agricultural Sciences,2015,43(7):26-30.(in Chinese with English Abstract)
[26]BARON E E,WENCESLAU A A,ALVARES L E,et al.High level of polymorphism in the myostatin chicken gene:Proceedings of the 7th World Congress on Genetics Applied to Livestock Production,2002.[S.l.]:[s.n.],2002:19-23.
[27]YE X H,BROWN S R,NONES K,et al.Associations of myostatin genepolymorphismswithperformanceand mortality traits in broiler chickens.Genetics Selection Evolution,2007,39:73-89.
[28]朱智,吴登俊,徐宁迎.鸡Myostatin基因单核苷酸多态性及其对屠体性状的遗传效应分析.遗传,2007,29(5):593-598.ZHU Z,WU D J,XU N Y.SNPs of myostatin gene and its genetic effects on carcass traits in chicken.Hereditas,2007,29(5):593-598.(in Chinese with English Abstract)
[29]GURUPRASADK,REDDYBV,PANDITMW.Correlation between stability of a protein and its dipeptide composition:A novel approach for predicting in vivo stability of a protein from its primary sequence.Protein Engineering,1990,4(2):155-161.
[30]贾浩,张小白,宋晓峰.人类胞内蛋白半衰期与其亚细胞定位的相关性研究.计算机与应用化学,2011,28(4):411-414.JIA H,ZHANG X B,SONG X F.Relationship between intracellular protein half-life and subcellular localization in human cells.Computers and Applied Chemistry,2011,28(4):411-414.(in Chinese with English Abstract)
[31]杜晓华,李积友,杨阳,等.甘肃西门塔尔牛高台类群MSTN基因多态性与生物信息学研究.畜牧兽医杂志,2015,34(1):13-17.DU X H,LI J Y,YANG Y,et al.Polymorphism and bioinformatics analysis of MSTN gene in Gaotai Simmental Cattle of Gansu.Journal of Animal Science and Veterinary Medicine,2015,34(1):13-17.(in Chinese with English Abstract)
[32]王伟,连林生,李继中.猪MSTN基因生物信息学分析.安徽农业科学,2012,40(10):5943-5945.WANG W,LIAN L S,LI J Z,et al.Bioinformatics analysis on MSTN gene of swine.Journal of Anhui Agricultural Science,2012,40(10):5943-5945.(in Chinese with English Abstract)
