桢楠种子的发芽与多胚特性及初生苗的生长特征
2018-11-09胡红玲廖玉华胡庭兴陈玉凤周光良
谭 飞,陈 洪,胡红玲,廖玉华,胡庭兴,陈玉凤,周光良,王 曦
(1.四川农业大学林学院,成都 611130;2.四川省凉山州林业调查规划设计院,四川 凉山 615000;3.四川省泸州市林业科学研究院,四川 泸州 646000)
桢楠(Phoebe zhennanS.Lee)为樟科(Lauraceae)楠属(Phoebe)常绿乔木,是我国特有的珍稀树种,喜温暖湿润气候,常生长于山麓肥厚腐殖质酸性至中性土壤[1],是国家二级重点保护树种[2],其树姿优美,具有很高的观赏价值,在城市建设和园林绿化中广泛栽培。材质优良,结构细致,纹理光滑、美观,容易加工,用途广泛,在建筑装饰以及家具生产中是贵重的高档用材[3]。桢楠种苗生产以种子为主,但其自身的繁殖能力和自然更新效率低下[4],由于种群稀少,种质资源有限,野外林下发芽率不高,成苗率更低,很难见到自然生长的桢楠幼树,加之长期以来的过度采伐利用,致使我国桢楠资源近于枯竭。而近年来国民经济建设对桢楠木材的需求又不断增长,导致其供求矛盾日益加剧[5]。为了保育桢楠种质资源,缓解其供求矛盾,有必要充分利用现有技术条件加速桢楠人工林的发展,促进桢楠资源的扩大再生产。近年来我们在桢楠苗木生产中,发现普遍存在多胚现象,虽然增加了苗木的总量但降低了幼苗的数量,增加了苗木规范化标准化培育的难度,因此有必要探讨桢楠种子的发芽及多胚苗特性及其对多胚苗生长的影响,以满足生态环境建设和社会经济发展对桢楠资源的需求。
研究表明,大量被子植物存在多胚现象[6-7],最早于1719年,(Citrus)[8-9]在柑橘属植物中发现了多胚现象。之后在玉米(Zea mays)[10-12]、苎麻(Boehmeria nivea)[13]、韭菜(Alluim tuberosum)[14-15]、咖啡(Coffea liberica)[16]等作物中都发现了多胚苗,随后在芒果(Mangifera indica)[17]、黄花风铃木(Tabebuia chrysantha)[18]等林木上也有过双胚苗的报道,2000年前后国外也曾有相关报道[19-20]。目前对楠属植物多胚现象的研究也有报道,如舒金枝[21]在试验中首次发现桢楠存在多苗现象;余道平等[22-23]对细叶楠(Phoebe hui)种子的萌发特性和幼苗性状进行了观测,也发现其种子存在多胚现象,多胚率达11.1%,其中双胚苗为8.1%,其余为3胚苗和4胚苗,黄宇[24]研究发现闽楠(Phoebe bournei)种子的发芽率和多胚率与其贮藏时间和培养条件都有显著的相关性;虽然前人的研究已经涉及了楠属植物多胚苗的报道,但并未涉及桢楠种子多胚率、发芽率与母树个体和种粒大小是否存在一定的相关性以及桢楠多胚苗的生长特性。本研究旨在探讨其多胚率的变化规律、初生多胚苗的生长特性以及对苗木质量的影响,以期为优良桢楠母树的确认、种子质量控制、优良幼苗个体的选择和提高苗木出圃质量提供理论依据和技术参考。
1 材料和方法
1.1 试验区概况
育苗试验地设在雅安市雨城区四川农业大学科研园区大棚内,位于东经 102°59′16″,北纬 29°58′52″,海拔580 m。多年平均气温16.2℃,≥10℃的积温5 231℃,月平均最高温29.9℃(7月),月平均最低温3.7℃(1月),年均日照时数1 039.6 h,无霜期298 d,多年平均降雨量1 774.3 mm,空气平均湿度79%,属于亚热带湿润气候。
1.2 试验材料
试验于2014年和2016年分两次进行,第2次试验的目的主要是验证第1次试验的重复性并补充测定相关指标,供试的桢楠种子分别于2013年和2015年11月采收。根据当年母树开花结实情况,在雅安市雨城区小宝洞种群选择母树6株,在雨城区金凤寺风景区种群选择3株,在都江堰市灵岩山种群选择3株。
各母树按树龄组依次编号分别为1、2、3号(雅安市雨城区小宝洞,60 a生),4、5、6号(都江堰市灵岩山,80 a生),7、8、9号(雅安市雨城区小宝洞,110 a生),10、11、12号(雅安市雨城区金凤寺景区,150 a生)。其种子粒级划分为大粒、中粒、小粒,采用称重法得到不同母树种子千粒重后,分别将大粒和小粒种子选出达到千粒重1/3为区分大、中、小粒的标准[25],共36份材料,每份材料3次重复,每次重复为100粒种子。
育苗基质由珍珠岩和蛭石以1∶1比例混合组成。
1.3 种子贮藏与育苗
1.3.1 种子的采集与贮藏
将收集的成熟桢楠种子置于室内堆积数日,待果皮腐烂后,人工揉搓,用清水将种子冲洗干净,并采用水浮法剔除劣质桢楠种子。选出的种子再用洗衣粉水洗净种子上的油脂,以利其发芽[21]。洗净的种子在室内阴干后分级,测定各粒级种子的大小规格和重量,然后将其与干净河沙按1∶4比例混合均匀进行沙藏,种、沙混合物的湿度保持60%,贮藏于大棚内,保持贮藏体透气和排水良好。
1.3.2 种子育苗
育苗当年3月中旬对苗床进行整理和施肥。后将沙藏的种子取出。以母树为单元、按其大、中、小不同粒级种子分组播种,每组100粒,重复3次。采用条播法,在已经整理好的苗床上按行距10 cm、间距3 cm、深度为5 cm开沟,把种子平放在沟中,盖土厚度为l~2 cm,播种完后再浇少量水。每3 d观察种子发芽情况,并及时补水,用HH2土壤水分测定仪(ML2x,GBR)监测苗床含水量,将基质体积含水量控制在15%~20%左右(约为试验用土田间持水量的70%~80%),种子出苗后持续观察初生苗生长情况,待幼苗株高达10 cm,单株有3~5片真叶完全打开后(苗龄约90 d)起苗计数单胚、双胚至更多胚数量,检查多胚苗健康状况,并测定各类幼苗株高,地径,然后选择健康苗移栽至营养袋供进一步观察。
1.4 指标测定方法
1.4.1 发芽率测定
在初生幼苗起苗时测定。发芽率=发芽的种子数/播种的种子数(100粒)×100%
1.4.2 多胚率测定
在初生苗起苗时测定,仍以母树为单元,按不同粒级种子计算多胚率,并统计单胚率、双胚率、三胚率及其以上等多胚率,每粒级100粒种子出土的初生苗和未出土的坏芽全部纳入计算。同时按形态大小区分多胚苗类型,分别记为单胚苗(D)、双胚大苗(SD)、双胚小苗(SX)、三胚大苗(TD)、三胚中苗(TZ)、三胚小苗(TX)以及四胚苗各单株的株高、地径和生物量。多胚率计算如下:
双胚苗频率=出现双胚苗的种子数/播种的种子数×100%
三胚苗频率=出现三胚苗的种子数/播种的种子数×100%
四胚苗频率=出现四胚苗的种子数/播种的种子数×100%
多胚苗频率=双胚苗频率+三胚苗频率+四胚苗频率
观测中,未发现五胚及以上的多胚苗,而四胚苗种子数量很少且成苗率很低,达不到统计分析的数量要求,因此未纳入统计分析。
1.4.3 幼苗形态指标测定
幼苗形态采用直尺测量株高(0.1 cm),电子游标卡尺测量地径(0.01 mm),地径从两个相互垂直的方向测定,取其平均值作为实际地径,每类幼苗分3组重复,每重测定7株。
1.5 数据处理
采用SPSS 20.0统计分析软件(SPSS Inc.,USA)对试验数据进行单因素方差分析(One-way ANOVA),并用最小显著差数法(LSD法)进行多重比较。
2 结果与分析
2.1 不同树龄母树、不同粒级种子的千粒重
由表1可知,不同桢楠母树的树高、胸径与树龄呈现正相关的关系。12株桢楠母树种子千粒重均表现为随粒级的减小而减小,且粒级间呈显著差异(P<0.05),这种趋势也不因树龄的不同而改变。其中大粒种子千粒重比中粒种子增加了(11.00%~32.11%)、比小粒种子增加了(61.12%~97.81%)。生长环境一致的3株同龄母树种子重量之间差异不显著(P>0.05),不同树龄母树各粒级的种子千粒重量总体上差异显著(P<0.05)。由表1还可以看出,80 a生母树的种子千粒重均值分别超过60、110、150 a生母树种子30.15%、17.29%、36.26%,其差异达显著水平(P<0.05)。表明不同母树种子的千粒重可能存在随母树树龄变化呈先增后降的趋势;也有可能是都江堰与雅安两地的生境不同造成的,也有可能是不同种源的遗传学特性有差异,使得种子千粒重差异较大。对此还需进行深入的研究。
2.2 不同树龄母树及不同粒级种子的多胚率和发芽率
由表2可知,从60~150 a 4个不同树龄组的12株桢楠母树种子都存在多胚现象,均表现为多胚率随胚数的增加呈下降趋势,其中150 a树龄各母树间单胚率和双胚率差异不显著(P>0.05),而与三胚率和四胚率以及三胚与四胚率之间差异显著(P<0.05)。其他3个树龄组各胚数之间多胚率差异均显著(P<0.05);而不同母树间无论是单胚率还是双胚率,总体上都表现出显著的差异,但三胚率和四胚率各母树间差不显著。由表2还可看出,多胚率最高达 60.00%(12号母树,150 a),最低为 6.33%(4号母树,80 a),而且不同树龄母树间多胚率差异显著,尤其是150 a生母树与其他树龄母树种子单胚率、双胚率、三胚率、四胚率和多胚率尤为不同,但并没表现出与树龄的相关性,这可能是种源的生境不同或遗传学特性不同,使得种子多胚率差异较大。
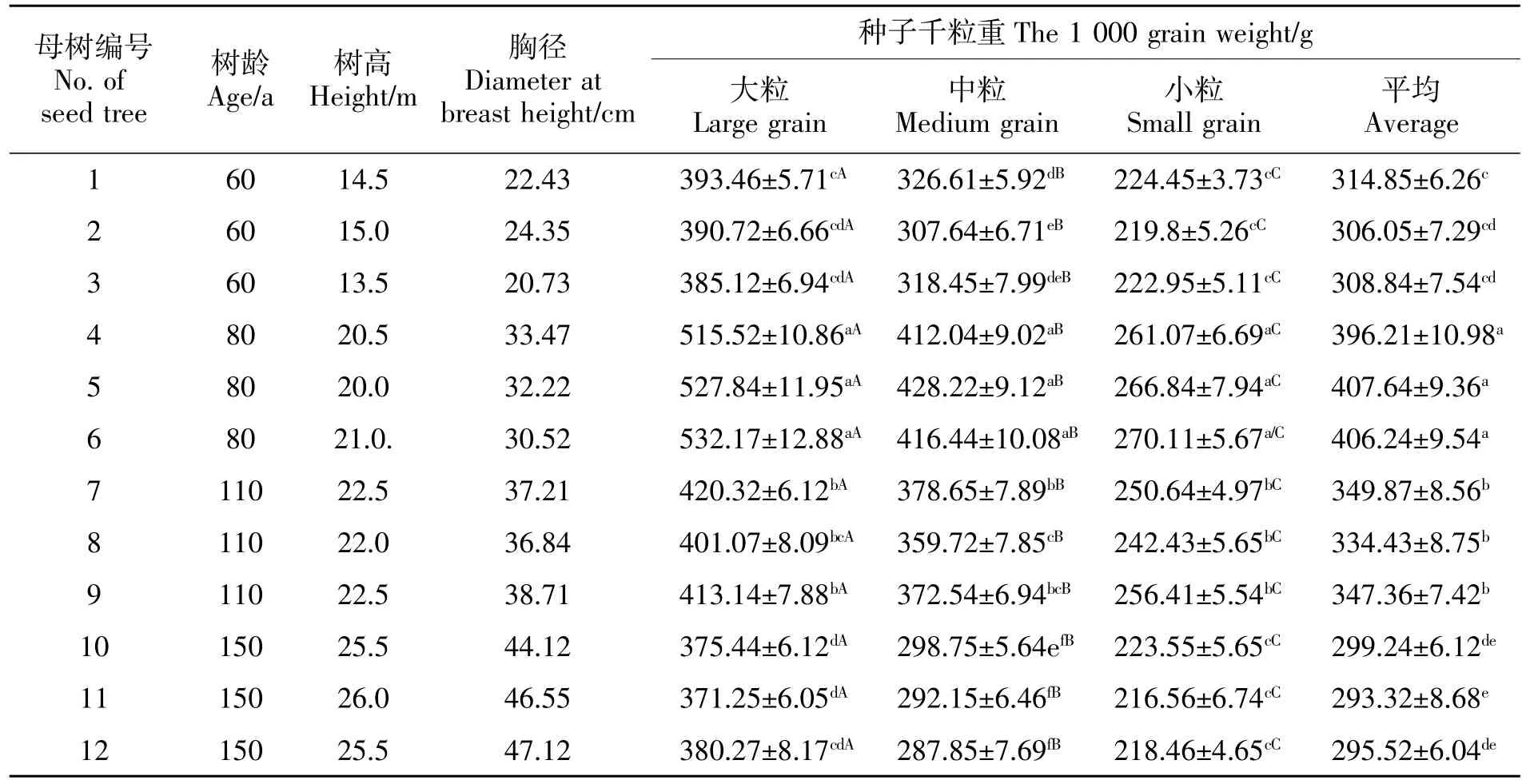
表1 不同树龄桢楠母树各粒级种子千粒重Table 1 1000 seed weight of P.zhennan seed trees with different age
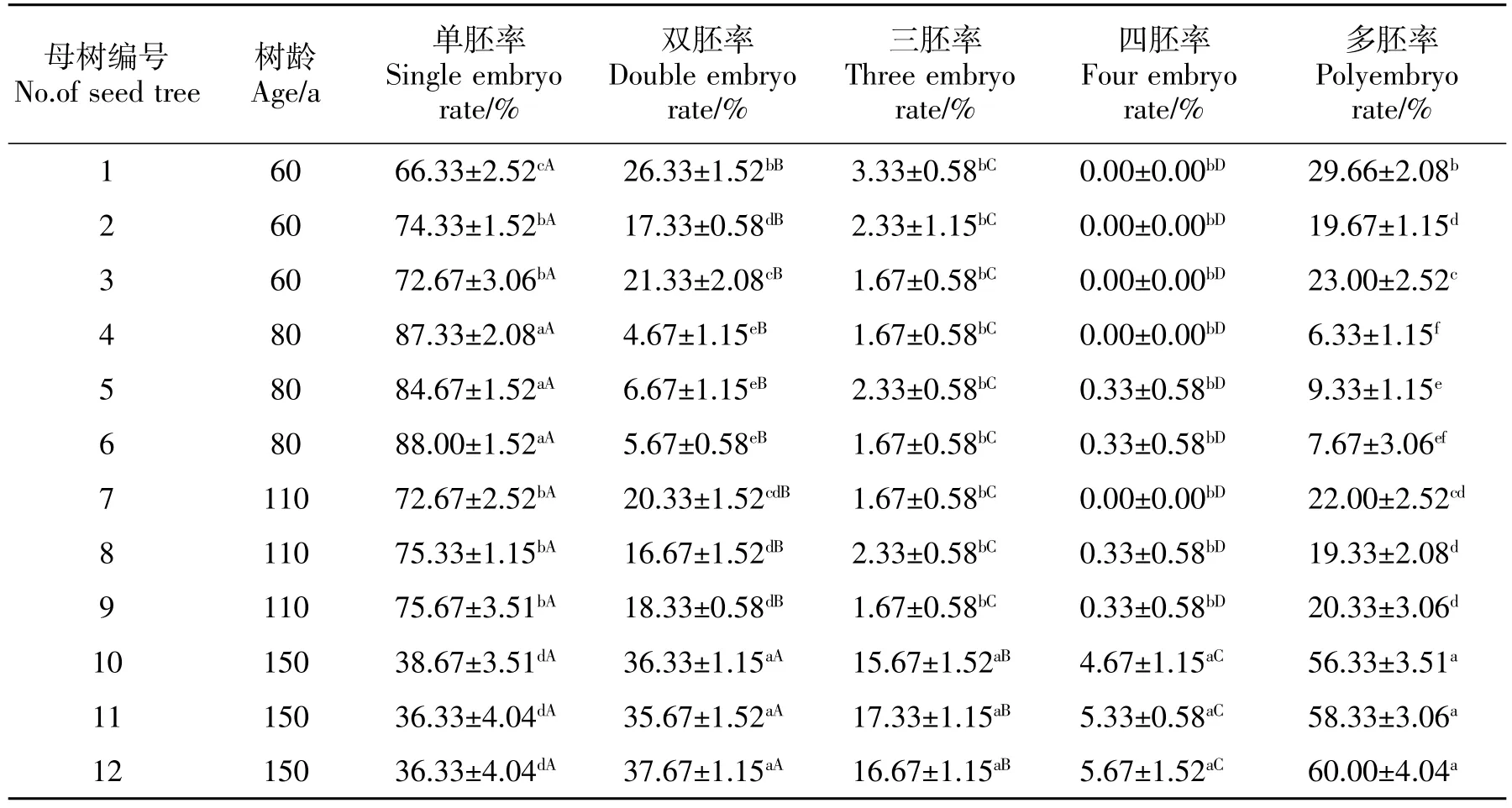
表2 不同树龄桢楠母树种子的多胚率和发芽率Table 2 The polyembryo rate and germination rate of P.zhennan seed trees with different age
由表3可知,12株桢楠母树中除了3号、8号和9号母树种子多胚率与种子粒级呈现显著下降的趋势外,其他母树种子多胚率与种子粒级无明显的相关性;不同母树不同粒级种子间的发芽率在92.33%~97.33%之间,且相互之间差异不显著(P>0.05),不同树龄母树各粒级种子间多胚率也不显著。由表3还可看出,就多胚率而言,不同母树之间都表现为双胚率最大,三胚率次之、四胚率最低。同时多胚率也没有表现出随母树树龄增加而呈现出规律性变化的趋势,同一母树不同种子粒级间多胚率差异不显著,表明种子粒级对多胚率影响不显著。
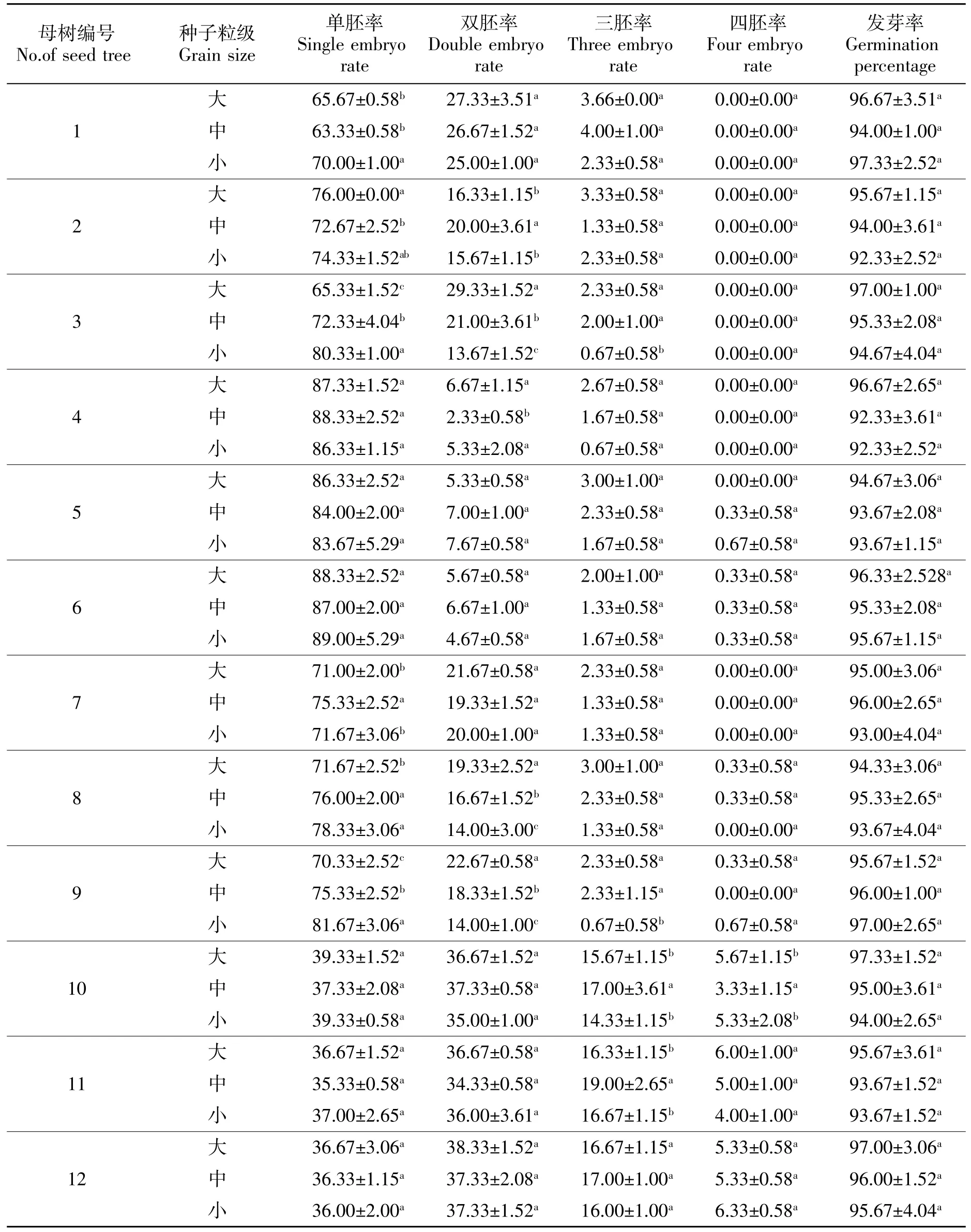
表3 各桢楠母树不同粒级种子的多胚率和发芽率Table 3 The polyembryony rate and germination rate of P.zhennan seeds with different sizes
2.3 种子粒级对多胚初生苗的株高、地径和生物量的影响
从图1可知,不同粒级的多胚幼苗的株高和地径都分别呈现单胚苗>双胚大苗≈三胚大苗>双胚小苗≈三胚中苗>三胚小苗的变化规律,且单胚苗各类指标均显著高于其他各类幼苗相同指标。其株高分别比其他各类幼苗高出的幅度分别为24.28%~31.84%、28.44%~37.86%、100.54%~117.73%、106.78%~138.91%和137.66%~187.74%;其地径比其他各类幼苗高出的幅度分别为7.86%~16.52%、10.71%~20.72%、37.17%~65.43%和86.11%~86.74%。幼苗的生长指标分别随着种子粒级由大到小呈现出相同的变化趋势,但大粒和中粒种子的生长指标差异不显著(P>0.05),而小粒种子的生长指标则显著低于大粒和中粒种子。
由图2可知,随着多胚苗的胚数的增加,其根、叶、茎、以及总全株干重呈现显著(P<0.05)下降的趋势。不同粒级的种子幼苗的生长指标随着多胚数的增加而降低,表明在多胚幼苗形成阶段,其种子自身营养有限,多胚苗越多分配的营养物质越少,因此多胚初生苗长势越弱。不同粒级的种子都是随着种子千粒重的增加其生长指标呈上升的趋势,表明种子规格越大,养分含量越充足,幼苗生长越好。
3 讨论
被子植物的多胚苗都起源于相应的多胚种子,且多胚的类型多样,因而其发生的机理比较复杂。目前,关于被子植物多胚苗的发生机理还没有形成统一的定论,自1998年黄群策综合分析被子植物多胚苗发生机理后,对多胚现象的机制已有多种解释[26]。冯辉等研究表明,咖啡、韭菜种子的多胚率为4.5%[14,16],谭志军等发现水稻种子多胚率在9.5%~14.4%之间,而用激素处理后可提高到29.4%~38.1%[27]。同时张斯淇等观察到柑橘育种亲本的多胚苗率为3.5%~7.2%,杂交后代可提高到28.0%~44.2%[8-9]。范志忠等发现甘蓝型油菜(Brassica napus)多胚苗高达 85.2%[28],而莫饶等发现芒果的多胚苗发生率更高,其多胚率高达98.9%[17]。楠属植物种子的多胚现象也很常见,本试验发现,参试的所有桢楠母树都存在多胚现象,其单株多胚率最高达到了60.0%,最低为6.33%,其中包括两胚、三胚和四胚,但不同母树都以双胚率最高。
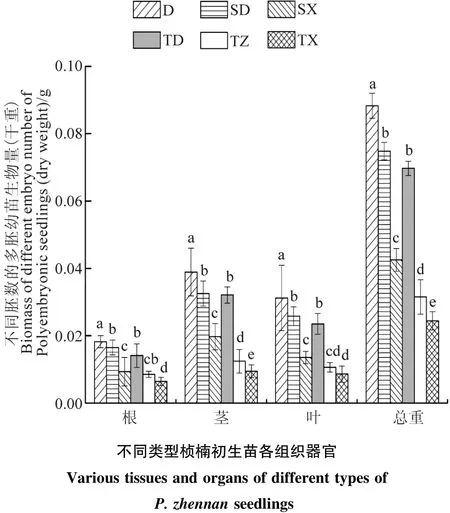
图2 桢楠不同类型多胚初生苗的生物量(干重)Figure 2 Biomass of different type of embryo polyembryonic seedling of P.zhennan(dry weight)/g
3.1 不同树龄桢楠母树、不同粒级种子对发芽率和多胚率的影响
本研究发现,同一种群同龄母树之间种子千粒重差异不显著,而不同树龄母树千粒重差异显著,表明种子千粒重可能随母树树龄的增加存在着先增后减的趋势;不同粒级桢楠种子千粒重差异显著,但对其发芽率和多胚率的影响都不显著;不同母树之间种子发芽率差异不显著,说明种子发芽率与母树个体无关,而同一种群同龄母树之间种子多胚率差异显著,但是不同树龄(组)母树之间种子多胚率没有呈现出规律性变化,表明桢楠种子多胚率与树龄没有相关性。许方等[29]研究发现,植株的年龄、年结实量、植株的营养状态和枝条的着生部位等因素对柑橘属植物的不定胚的发生数目有明显影响。此外,黎垣庆等[30]对水稻API-APIV多胚苗进行遗传学研究发现其双胚苗性状的表现主要受两对隐性基因控制,而双胚苗发生频率的高低则受某些修饰因子的影响。因此,我们猜测桢楠种子的多胚率可能与母树种子形成与发育过程中受到某些环境因素影响有关(如授粉过程、土壤营养条件以及环境温度等),也可能与母树个体的遗传特性有关,因而有必要从基因遗传学的角度来分析其多胚特性的控制与表达,这需要开展进一步的研究。
3.2 不同粒级种子对桢楠多胚幼苗生长指标的影响
有研究表明,在苗期阶段苗木的质量和出苗的整齐度受种子分级的影响较大,例如,姜景民等[31]对湿地松种子分级育苗后,发现在生长初期苗木的质量随种粒大小呈正相关的趋势;梁机等[32]在研究种粒大小对擎天树幼苗生长的试验中也发现大粒和中粒种子幼苗的苗高、地径和叶片数均显著高于小粒种子相应指标,而大粒和中粒种子幼苗之间的各项生长指标差异不显著;王志贤等[25]对文冠果种子分级后育苗,发现分级能显著提高出苗的整齐度和苗木质量,并可经济有效的利用种子。本试验中,不同粒级的桢楠种子的各项生长指标和生长情况都与上述相关研究结果相似,不同的母树种子千粒重越大其幼苗的各项生长指标越好,且随着种子粒级的增加,其幼苗生长指标也显著增加。表明桢楠种子重量越大,贮存的营养物质含量越高、活力更强,幼苗具有更佳的生长优势,因此优质种子是优良苗木的基础,在生产中应首先选择千粒重更大桢楠母树种子,或在批量种子选择的大、中粒种子进行育苗,从而培育出高质量的苗木。
本研究还发现,桢楠多胚苗植株各项生长指标有较大的差异,随着胚数增多,其长势越弱,且多胚苗的株高、地径、根、茎、叶、总生物重等指标都显著低于单胚苗,在生产上不能达到优质合格苗的要求,表明由于种子养分含量有限,发芽过程中形成的多胚苗健康状况相对较差,加之在前期育苗的过程中多胚苗之间仍然存在着竞争关系,虽然幼苗数量有所增加,但对苗木出圃质量有一定的影响,降低合格苗出圃率。谭飞等[33]研究发现,育苗过程中加强养分管理,每株施复合肥0.2 g可以明显促进桢楠多胚苗生长,缩小与单胚苗的生长指标的差距,对提高苗木出圃率、降低育苗成本有重要的作用。
综上所述,在本试验中,60~150 a树龄段的桢楠母树的种子多胚率和发芽率与母树的树龄和种子的粒级的大小没有显著的相关性,不同母树个体多胚率差异显著,可能与母树本身的遗传特性和环境条件有关,但各母树都呈现出双胚率最高,三胚率次之,四胚率最低的变化规律;单胚苗与多胚苗的株高、地径、根、茎、叶、总重差异显著,说明多胚率对幼苗的出圃质量有较大影响;在生产上应选用千粒重更大桢楠种子,或在批量种子中的选择大、中粒种子进行育苗,同时,在初生苗移栽过程,只保留单胚苗、双胚大苗和三胚大苗,可提高出圃苗木的数量和质量。
