利用动力学模型探讨甲萘醌对脱氮假单胞杆菌发酵过程的影响
2018-11-05章帅文李昆太
杨 勇,刘 群,章帅文,李昆太
利用动力学模型探讨甲萘醌对脱氮假单胞杆菌发酵过程的影响
杨 勇,刘 群,章帅文,李昆太*
(江西农业大学 生物科学与工程学院,江西 南昌 330045)
以脱氮假单胞杆菌为供试菌,通过外源添加氧化应激诱导剂甲萘醌,测定了维生素B12发酵过程中代谢特征参数,应用Logistic、Luedeking-Piret 和Luedeking-Piret 修正模型建立了氧化应激状态下菌体生长、产物积累和基质消耗的动力学方程,采用Origin9.0软件对其进行非线性回归分析,获得动力学模型参数。研究结果发现:甲萘醌可以加快菌体生长、加速总糖消耗,但对维生素B12合成具有一定的抑制作用;拟合结果表明甲萘醌能够有效降低菌体的最大比生长速率m、提高总糖的比消耗速率、降低维生素B12合成的比生成速率,延缓稳定期出现的时间,且拟合相关系数2均达到90%,说明该模型能较好地反映本研究实验条件下脱氮假单胞杆菌的细胞生长、产物合成和底物消耗过程,为接下来深入研究不同氧化应激状态下的菌体代谢变化特征提供参考。
脱氮假单胞杆菌;甲萘醌;维生素B12;相关系数;氧化应激
维生素B12(Vitamin B12,VB12)系含有钴离子的类咕啉(corrinoid)类化合物,通常被称为钴胺素,其分子结构是以钴离子为中心的咕啉环和5,6-二甲基苯并咪唑(DMBI)为碱基组成的核苷酸[1]。作为一种重要的生物活性物质,VB12还是很多重要生物化学反应所必需的辅酶[2],比如分子内重排、分子间甲基转移反应以及由三磷酸核糖核苷酸生成2′-三磷酸脱氧核糖核苷酸的还原反应等[3]。目前,VB12主要用于治疗恶性贫血症以及末梢神经炎[4]。由于其分子构造高度复杂,全部的化学合成需要多达70余个反应步骤[5],而且成本极高,因此目前基本都是采用微生物发酵形式来生产合成VB12[6]。自然界中主要存在两种不同的VB12生物合成途径,即好氧途径与厌氧途径[7],这两种合成途径的本质区别在于钴离子螯合到咕啉环的时机不同以及中心咕啉环的缩合机制不同[8]。脱氮假单胞杆菌由于具有完整的VB12合成基因,能够在好氧条件下完成VB12的全合成,因而被广泛应用于VB12的工业发酵生产[9]。
发酵动力学模型和方程的建立是为了解在发酵过程当中菌体生长、产物合成和基质消耗的动态平衡及内在代谢规律,为后续深入研究提供理论基础[10]。近年来,国内外针对脱氮假单胞杆菌生物合成VB12发酵动力学的研究多集中在分批补料过程,如:刘东洪[11]、刘晓侠[12]等人通过建立发酵动力学模型很好地反映了VB12以及前体物质5-氨基乙酰丙酸(5-ALA)分批补料发酵过程,对VB12的发酵生产具有一定的理论指导意义。脱氮假单胞杆菌作为典型的具有ED代谢途径的微生物[13],该途径生成的NADPH,除了参与VB12合成的酶促反应[14],还是微生物胞内抗氧化防御系统的重要组成成分,其作为电子供体参与胞内的氧化还原反应以维持胞内正常的氧化还原平衡[15-16],然而目前国内外对外源氧化应激诱导剂存在下的VB12发酵代谢研究鲜有报道。甲萘醌作为一种超氧生成剂,可以进入细胞,经单电子还原成半醌自由基,从而进入呼吸链与O2作用形成·O2-[17-18]。因此,外源添加甲萘醌可以作为一种氧化应激诱导剂,改变菌体培养环境的氧化程度,起到添加活性氧的作用。
为此,本试验以脱氮假单胞杆菌为供试菌体,在外源添加甲萘醌对脱氮假单胞杆菌发酵过程的研究基础上,通过origin9.0非线性拟合,建立了菌体生长、VB12合成及基质消耗3个动力学模型,以期为接下来深入研究脱氮假单胞杆菌在维系菌体胞内氧化还原平衡和VB12合成中的关键调控作用,揭示限制性供氧促VB12高效合成的代谢机理提供试验依据和研究基础。
1 材料与方法
1.1 材料与试剂
1.1.1 菌种 脱氮假单胞杆菌(),由本实验室保存。
1.1.2 培养基 斜面培养基:蔗糖30 g,蛋白胨10 g,玉米浆10 g,(NH4)2SO40.25 g,(NH4)2HPO41.5 g,MnSO4·H2O 0.1 g,ZnSO4·7H2O 0.1 g,琼脂20 g,去离子水定容至1 000 mL。灭菌前调pH至7.0~7.2。
种子培养基:蔗糖35 g,蛋白胨20 g,KH2PO45 g,(NH4)2SO42.0 g,(NH4)2HPO40.8 g,MgSO41.5 g,ZnSO4·7H2O 0.02 g,MnSO4·H2O 0.2 g,去离子水定容至1 000 mL。灭菌前调pH至7.2~7.4。
发酵培养基:蔗糖50 g,蛋白胨25 g,(NH4)2SO41 g,ZnSO4·7H2O 0.08 g,MgSO42 g,KH2PO40.8 g,甜菜碱10 g,5,6-二甲基苯并咪唑(DMBI) 0.08 g,CoCl2·6H2O 0.15 g,CaCO32 g,去离子水定容至1 000 mL。灭菌前调pH至7.2~7.4。
以上培养基均在121 ℃下灭菌20 min。
1.1.3 材料 氢氧化钠等常用试剂均为分析纯,西陇科学股份有限公司;蛋白胨(生化试剂),奥博星生物技术有限公司;甲萘醌,国药集团化学试剂有限公司;VB12标准品,阿拉丁生化科技股份有限公司。
1.2 仪器与设备
立式压力蒸汽灭菌锅,上海申安医疗器械厂;恒温培养箱,韶关泰宏医疗器械有限公司;摇床、超净工作台,博迅医疗生物仪器股份有限公司;JW-3022高速冷冻离心机,安徽嘉文仪器装备有限公司;UV759紫外分光光度计,上海佑科仪器仪表有限公司;KW-1000DC HH恒温水浴锅,金坛市中大仪器厂。
1.3 摇瓶发酵方法
种子液制备:每支斜面(18 mm×180 mm)以10 mL无菌水刮洗制成菌悬液。取2 mL至装液量为50 mL/250 mL三角瓶的种子培养基中,180 r/min,30 ℃下摇床培养至菌体光密度(700)为9~10。
摇瓶发酵:将培养好的种子液按照10%接种量接种至装液量为40 mL/250 mL三角瓶的发酵培养基中;甲萘醌溶液用0.25 μm有机相滤膜无菌过滤后按照相应的摩尔浓度梯度(浓度梯度分别为0,0.3 μmol/L)添加到发酵培养基中,30 ℃、180 r/min下摇瓶发酵培养。
1.4 分析方法
菌体生物量[19]、VB12含量[7]、残总糖[20]的测定按参考文献的方法进行。
1.5 试验数据获得与处理
在发酵培养过程中,每隔24 h取样,分别测定菌体生物量、总糖以及VB12含量。每个实验做3次平行实验,取其平均值。
采用Excel2003与DPS7.05进行数据处理,origin9.0软件进行作图与动力学拟合分析。
1.6 动力学模型
1.6.1 菌体生长动力学模型 Logistic[21]方程是由Verhulst-Peal提出,可以直观的显示菌体在发酵过程当中的生长规律,对拟合菌体的生长具有广泛的适用性。其方程形式如下:

式中:为菌体生物量浓度;为比生长速率,h-1;表示时间,h;Xmax表示菌体生物浓度最大值。
在发酵生产VB12的过程中,存在着许许多多影响产物合成的因素,譬如:高浓度甜菜碱分解物质的抑制作用,前体物质甘氨酸、琥珀酰CoA和谷氨酸合成速率的影响,供氧水平与菌体形态的变化,以及菌体生长所需的营养成分扩散限制而导致菌体生长迟缓等,因此,发酵过程并不是一个均衡生长的过程。从图1可以看出,菌体浓度呈S型增长,而Logistic模型恰好是一个非常典型的S型曲线,能够比较准确地反映出发酵过程中菌体浓度变化对自身生长的作用,以及拟合发酵过程中菌体的生长规律。
以时间等于0,以0为起始条件,对式(1)积分可得生物量与时间的函数:

利用origin9.0软件将()对时间进行非线性拟合,可获得最大比生长速率max,0和max。相关系数2可用来评价模型的拟合程度。
1.6.2 产物生成动力学模型 从图1可以看出,在整个发酵过程当中,VB12合成与生长存在相关联系。VB12作为胞内产物,具备充足的菌体生物量是发酵高产的前提条件。根据发酵过程及试验数据来看,VB12的合成与生物量随时间的变化趋势相同,当菌体处于对数生长期,产物VB12同时产生,但只是部分合成;菌体生长停滞后,VB12继续以某种形式大量合成。根据文献记载,Gaden[22]将产物合成与菌体生长及基质消耗相关联,定性地将发酵过程归为3类:生长偶联型、非生长偶联型和部分生长偶联型。因此,发酵合成VB12的发酵过程为部分生长偶联型[11]。
部分生长偶联模型可用Leudeking-piret方程[10]表示:

式中:为产物浓度,μg/mL;,是经验常数(,均大于0),为与菌体量关联的产物合成常数[g/(g·h)-1],为与菌体生长率关联的产物合成常数(g/g)。
把式(2)代入式(3),并求积分得:

利用origin9.0软件()对进行非线性拟合,可获得,及初始值0,相关系数2可用来评价模型拟合程度。
1.6.3 基质消耗动力学模型 根据物料平衡法则,在VB12的发酵过程中,基质(蔗糖)的消耗主要可分为3个部分,即:构成菌体主要成分,维持菌体生长代谢的需要,合成产物VB12。因此,基质消耗速率可以采用Luedeking-piret修正模型表示:

式中:k为细胞维持系数;Y为菌体对基质的得率系数;为基质浓度,μg/mL;Y为产物对基质的得率系数。把式(3)代入式(5),得:
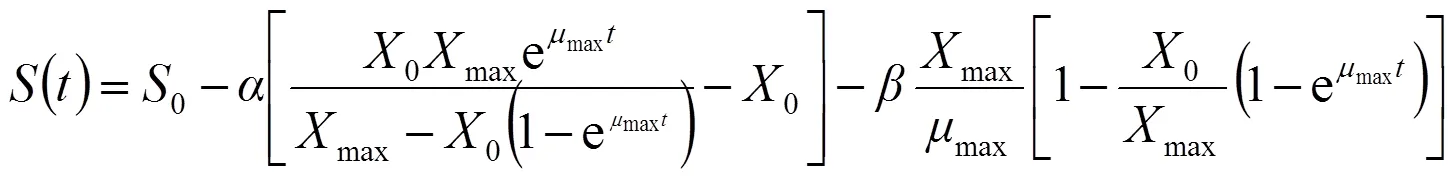
式(1)、(3)和(6)分别构成了发酵过程中的菌体生长、VB12合成以及总糖消耗的动力学模型,反映了发酵过程当中菌体生长、VB12合成及总糖消耗同发酵时间之间的关系;式(2)、式(4)和式(7)分别为3个模型的积分形式,便于模型的拟合求解。将()对进行非线性拟合,可获得模型参数、,初始值0。相关系数2可以用来评价模型的拟合程度。
2 结果与分析
2.1 甲萘醌对P. denitrificans发酵过程代谢变化特征影响
由图1可知,在0~24 h期间,菌体处于适应期,对照组与试验组菌体量都没有显著增加;随后,菌体进入对数生长期,在24~72 h期间,甲萘醌出现明显抑制细胞生长的作用。甲萘醌作为一种苯醌化合物,通过氧化还原体系不断地产生ROS,从而通过氧化压力导致细胞损伤[23],因此发酵初期对照组菌体量高于试验组,可能是由于甲萘醌对于细胞生长以及相关的酶系具有抑制作用。72 h后,试验组菌体量开始高于对照组,可能是菌体开始适应胞外胁迫环境并利用甲萘醌。随着发酵的进行,代谢不断加剧,菌体生物量急剧增加,残糖急剧下降;96 h之后,菌体进入稳定期及衰退期,残糖量变化基本不变了。与对照组相比,添加甲萘醌加速了菌体的糖耗,侧面反映了添加甲萘醌有利于菌体的生物量形成。另外,添加甲萘醌情况下,VB12产量也呈现出明显的降低趋势。尽管该甲萘醌浓度下的菌体生物量已达到最大值,但其VB12产量仍比对照组降低了7.5%,仅为56.82 μg/mL。
综上可知,添加甲萘醌可以加快菌体生长、加速总糖消耗,但是对VB12的合成具有一定的抑制作用。

□:细胞浓度(添加甲萘醌);■:细胞浓度(不添加甲萘醌);○:VB12添加甲萘醌;●:VB12不添加甲萘醌;△:总糖(添加甲萘醌);▲:总糖(不添加甲萘醌)
2.2 VB12发酵动力学模型拟合求解与检验
将图1中的实验数据应用于上述1.6中的各模型,采用Origin9.0软件对实验数据进行非线性拟合,得出各参数值及初始值,结果如表1所示。

表1 动力学模拟参数模拟值
将各个参数与初始值代入式(1)、式(3)和式(6),由此,我们可以得到本试验的发酵动力学模型:
(1)菌体生长动力学模型为:


(2)VB12合成动力学模型为:

(3)基质消耗动力学模型为:


将得到的各发酵动力学模型的拟合曲线与实际的实验数据相比较,结果见图2。
A1,B1,C1为对照组,A2,B2,C2为试验组
■实验数据(actual values);○拟合值(simulation values)
图2 动力学模型曲线与实验值的比较
从图2可以看出,所选择的模型基本上都能较好的反映发酵合成VB12的情况。对照组菌体生长和VB12合成的拟合度2分别为0.995 6、0.994 9,试验组二者的拟合度分别为0.990 5、0.997 5,拟合值与实验值吻合度好,相对误差均<1%。对照组和试验组总糖消耗的拟合度也分别达到了0.939 2、0.925 5,拟合值与实验值吻合度较好。此外,由图可知,无论是否添加甲萘醌,菌体的生长曲线均为典型的S型曲线。在0—24 h,菌体处于适应期,添加甲萘醌能明显降低该阶段菌体生长速率。随后菌体进入对数生长期,菌体浓度显著上升,残糖含量急剧下降。菌体发酵进入稳定期,菌体量和VB12含量峰值就出现在此时期。综合图2和实验数据可以看出,甲萘醌的添加能够有效降低菌体的最大比生长速率m、提高总糖的比消耗速率、降低VB12合成的比生成速率,延缓稳定期出现的时间。
3 结论与讨论
发酵过程的理论与方法已被广泛地应用于工业微生物产品生产及工艺优化,使得现代工业微生物发酵取得了高速发展[24]。Kang等[5]通过一系列发酵工艺的优化,使的VB12工业化发酵水平近年来得到了大幅度的提高,稳定在200 mg/L以上。但作为具有VB12好氧合成途径的菌株,是否会与嗜糖假单胞菌()[25]、恶臭假单胞菌()[26]等大多假单胞菌类一样,对胞内氧化环境较为敏感。为此,本文首先通过外源添加氧化应激诱导剂甲萘醌,利用进行摇瓶发酵实验,测定了VB12合成过程中的菌体量、VB12和残糖含量等代谢特征参数。根据结果显示,甲萘醌可以促进菌体生长、加速总糖消耗,但是对VB12合成具有一定的抑制作用。结合文献报道[27],可以分析得出,氧化应激诱导剂甲萘醌的加入虽然能延缓菌体稳定期出现的时间,显著增强NADP磷酸酶的酶活,增加胞内NADPH的供给,但同样诱发了胞内氧化环境的加剧,迫使菌体不得不将更多的NADPH用于抗氧化应激并维持胞内的氧化还原平衡,从而使得流向VB12合成途径的NADPH偏少,从而最终影响VB12的合成量。
发酵动力学模型是研究各种环境因素与微生物代谢活动之间的相互作用随时间变化规律的手段, 目的在于按照人们的需求来控制发酵过程[28],对发酵工艺的分析、优化、放大和控制具有非常重要的指导意义[29]。刘东洪等[11]以为供试菌株, 对VB12分批补料发酵动力学模型进行研究,建立了该菌株发酵合成VB12的菌体生长、产物合成和总糖消耗的动力学模型,较好地反映了VB12分批补料的发酵过程;Li等[30]通过研究发现,相比较于发酵过程中较高的DO控制浓度,限制性DO调控策略反而更有利于中的VB12合成。基于菌体在不同DO水平下的代谢调节机制,本文对氧化应激诱导剂甲萘醌状态下的菌体发酵产VB12动力学变化进行了研究,通过建立发酵动力学模型,可以实时监控发酵过程中菌体生长、代谢产物合成、基质消耗的变化规律。模型拟合结果表明,对照组菌体生长、VB12合成以及总糖消耗的拟合度2分别为0.995 6、0.994 9、0.939 2,试验组三者的拟合度分别为0.990 5、0.997 5、0.925 5,拟合值与实验值相近,模型能较好地反映发酵过程规律。
本文通过外源添加氧化应激诱导剂甲萘醌,测定了VB12发酵过程中代谢特征参数,建立了氧化应激状态下菌体发酵动力学模型,对深入探究在氧化应激状态下的代谢辅因子变化规律,明确其在维系菌体胞内氧化还原平衡和VB12合成中的关键调控作用,揭示限制性供氧促VB12高效合成的代谢机理具有重要指导意义。
[1] 李永毅. 谢氏丙酸菌维生素B12生物合成核糖开关敲除的功能研究[D]. 石家庄: 河北科技大学, 2013.
[2] 杨勇, 龙悦, 吴志明, 等. 柠檬酸钠对脱氮假单胞杆菌发酵产维生素B12代谢过程的影响[J]. 江西农业大学学报, 2018, 40(1): 174-181.
[3] Randaccio L, Geremia S, Demitri N, et al. Vitamin B12: Unique metalorganic compounds and the most complex vitamins[J]. Molecules, 2010, 15(5): 3228-3259.
[4] Stabler S P. Vitamin B12deficiency reply[J].New England Journal of Medicine, 2013, 368(21): 2041-2042.
[5] Kang Z, Zhang J, Zhou J, et al. Recent advances in microbial production of δ-aminolevulinic acid and vitamin B12[J]. Biotechnology Advances, 2012, 30(6): 1533-1542.
[6] 张翠所, 李昆太. 维生素B12的研究进展[J]. 中国酿造, 2008, 27(12): 17-20.
[7] Martens H, Barg H, Warren M J, et al. Microbial production of vitamin B12[J]. Applied Microbiology & Biotechnology, 2002, 58(3): 275-285.
[8] Moore S J, Biedendieck R, Lawrence A D, et al. Characterization of the enzyme CbiH60involved in anaerobic ring contraction of the cobalamin (Vitamin B12) biosynthetic pathway[J]. Journal of Biological Chemistry, 2013, 288(1): 297-305.
[9] Ainala S K, Somasundar A, Park S. Complete genome sequence ofATCC 13867[J]. Genome Announcements, 2013, 1(3): 63-96.
[10] 薛文娇, 马赛箭, 常帆, 等. 利用动力学模型探讨碳源浓度对普鲁兰分批发酵过程的影响[J]. 化学工程, 2015, 43(11): 5-10.
[11] 刘东洪, 李昆太, 王永红, 等. 脱氮假单胞杆菌产维生素B12分批补料发酵动力学研究[J]. 化学与生物工程, 2008, 25(5): 43-45.
[12] 刘晓侠. 5-氨基乙酰丙酸合成酶基因的克隆表达及生物合成5-氨基乙酰丙酸的研究[D]. 杭州: 浙江大学, 2006.
[13] Wang Z J, Wang P, Liu Y W, et al. Metabolic flux analysis of the central carbon metabolism of the industrial vitamin B12producing strainusing13C-labeled glucose[J]. Journal of the Taiwan Institute of Chemical Engineers, 2012, 43(2): 181-187.
[14] Warren M J, Raux E,Schubert H L, et al. The biosynthesis of adenosylcobalamin (vitamin B12)[J].Natural Product Reports, 2002, 19(4): 390-412.
[15] Flamholz A, Noor E, Bar-Even A, et al.Glycolytic strategy as a tradeoff between energy yield and proteincost[J].Proceedings of the National Academy of Sciences of the United States of America, 2013, 110(24): 10039-10044.
[16] Ying W. NAD+/NADH and NADP+/NADPH in cellular functions and cell death: regulation and biological consequences[J]. Antioxidants & Redox Signaling, 2008, 10(2): 179-206.
[17] Thor H, Smith M T, Hartzell P, et al. The metabolism of Menadione (2-Methyl-1,4-naphthaquinone) by isolated hepatocytes [J].Journal of Biological Chemistry, 1982, 257(20): 12419-12425.
[18] Michaela K, Linda M H, Brian M. Oxidative stress response of a recombinant aspergillus niger to exogenous menadione and H2O2addition [J]. Enzyme & Microbial Technology, 2002, 30(3): 346-353.
[19] 彭卫福, 陈未, 周佳, 等. 无机氮源对脱氮假单胞杆菌产维生素B12代谢过程的影响[J]. 食品工业科技, 2013, 34(18): 178-181.
[20] 张文平, 李昆太, 黄林, 等. 产胞外多糖菌株的筛选及其对土壤团聚体的影响[J]. 江西农业大学学报, 2017, 39(4): 772-779.
[21] Phisalaphong M, Srirattana N, Tanthapanichakoon W. Mathematical modeling to investigate temperature effect on kinetic parameters of ethanol fermentation[J]. Biochemical Engineering Journal, 2006, 28(1): 36-43.
[22] Gaden E L. Fermentation kinetics and productivity[J]. J Biochem Microhiol Technol,1959,1(4): 413-429.
[23] 麻攀, 刘洪涛, 许青松, 等. 壳寡糖缓解甲萘醌诱导巨噬细胞损伤机制初探[J]. 中国生物工程杂志, 2011, 31(6):18-21.
[24] 夏永, 王斌, 王公孝, 等. C群流行性脑膜炎球菌多糖分批发酵动力学模型的建立[J]. 中国生物制品学杂志, 2014, 27(11): 1473-1476.
[25] Stettner A I, Segrè D. The cost of efficiency in energy metabolism[J]. Proceedings of the National Academy of Sciences of the United States of America, 2013, 110(24): 9629-9630.
[26] Nikel P I, Kim J, de Lorenzo V. Metabolic and regulatory rearrangements underlying glycerol metabolism inKT2240[J]. Environmental Microbiology, 2014, 16(1): 239-254.
[27] Singh R, Mailloux R D S, Appanna V. Oxidative stress evokes a metabolic adaptation that favors increased NADPH synthesis and decreased NADH production in[J]. Journal of Bacteriology, 2007, 189(18): 6665-6675.
[28] 程新, 夏海林, 黄林, 等. 红曲霉JR发酵动力学模型的建立[J].江西农业大学学报, 2010, 32(3): 590-594.
[29] 姜言晖, 张京良, 江晓路. 球形节杆菌A152产透明质酸酶的发酵优化及动力学模型的建立[J]. 生物工程学报, 2017, 33(11): 1883-1888.
[30] Kun-Tai L, Jia Z, Xin C, et al. Study on the dissolved oxygen control strategy in the large-scale vitamin B12fermentation by[J]. Journal of Chemical Technology & Biotechnology, 2012, 87(12): 1648-1653.
Study on Effects of Menadione onFermentation by Kinetics Model
YANG Yong, LIU Qun, ZHANG Shuai-wen, LI Kun-tai*
(College of Biological Science and Engineering, Jiangxi Agricultural University, Nanchang 330045, China)
In this paper, usingas the producing strain, the metabolic parameters of vitamin B12fermentation were studied by adding the exogenous oxidative stress inducer menadione. And the kinetic equations of biomass growth, production formation and substrate consumptionwere established by applying the Logistic, the Luedeking-Piret and the modified Luedeking-Piret equations. The non-linear curves were fitted and the kinetic parameters were calculated with Origin9.0. The results showed that menadione could promote the growth of bacteria and accelerate the substrate consumption, but significantly inhibited the synthesis of vitamin B12. By fitting, it was found that menadione could effectively reduce the maximum specific growth rate of bacteria, increase the specific consumption rate of total sugar, reduce the specific production rate of vitamin synthesis, and delay the emergence time of stable phase. And all the fitting correlation coefficients2were up to 90%, which showed the model could well reflect the cell growth, product synthesis and substrate consumption of. This study provided a reference for the further study of metabolic characteristics of bacteria under different oxidative stress condition.
; menadione; vitamin B12; correlation coefficient; oxidative stress
Q924
A
2095-3704(2018)03-0184-08
2018-06-19
国家自然科学基金项目(209005220)、江西省(青年)自然科学基金重大项目(20143ACB2100)和江西省青年科学家培养计划项目(20142BCB23025)
杨勇(1993—),男,硕士生,主要从事微生物代谢调控方面的研究,wy931221@sina.com;
李昆太,教授,博士,atai78@sina.com。
杨勇, 刘群, 章帅文, 等. 利用动力学模型探讨甲萘醌对脱氮假单胞杆菌发酵过程的影响[J]. 生物灾害科学, 2018, 41(3): 184-191.
10.3969/j.issn.2095-3704.2018.03.41