秋季降温期水温和饵料量对沙蜇无性繁殖的影响
2018-11-02邢勤旺王佳晗
孙 明,邢勤旺,2,王佳晗,段 妍,李 楠,3,柴 雨,董 婧,*
1 辽宁省海洋水产科学研究院,辽宁省海洋生物资源与生态学重点实验室,大连 116023 2 中国海洋大学水产学院,青岛 266003 3 上海海洋大学海洋科学学院,上海 201306
全球变化背景下,继赤潮、绿潮等由植物快速繁殖引起的大型海洋生态灾害后,一种由海洋动物——水母的数量增多而导致的海洋生态灾害也受到全球的关注[1]。沙蜇(Nemopilemanomurai)是东亚水域大型灾害水母优势种之一,20世纪末以来,沙蜇频繁暴发给中国、日本和韩国沿海的渔业生产、海洋旅游业及人民健康安全带来极大地负面影响[2- 5]。近年来,随着水母暴发机理研究的逐渐深入,人们越发意识到环境条件对底栖螅状体阶段无性繁殖的影响对水母暴发形成的重要作用[6-10]。众多研究结果表明,在各环境因子对大型水母生长发育的影响研究中,温度和饵料应为研究的重点,水温与饵料的综合变化情况对水母生活史的发育方向与数量增长起着非常重要的调控作用[6,11-12]。
在室内对沙蜇螅状体进行了不同恒定温度条件下不同饵料量对沙蜇螅状体生长与足囊繁殖的影响研究[13],试验结果表明,沙蜇螅状体可在0—27.5℃范围内良好存活,7.5℃以下几乎没有足囊繁殖行为,螅状体可在低温不摄食的状态下成功越冬,在8—20℃范围内,螅状体生长、足囊形成及萌发均随水温和饵料量的增加而显著增加。由此可以分析出,沙蜇水母体在每年夏秋季经有性繁殖出的当龄螅状体,从出生到越冬前的无性繁殖量对于翌年春季横裂释放的碟状体数量有决定性影响,那么这一时期沙蜇螅状体足囊繁殖量受水温与饵料量的影响是否显著?是否会有横裂生殖发生?本文在实验室模拟条件研究了秋季降温期不同水温和饵料量条件对沙蜇无性繁殖的影响,探讨了不同降温速度和饵料环境下,沙蜇当龄螅状体足囊繁殖和横裂生殖的差异。研究结果对进一步揭示积温升高对沙蜇繁殖生活史特征的作用及探索沙蜇暴发机理有重要的学术价值。
1 实验材料与方法
1.1 试验材料
实验所用沙蜇螅状体为2016年9月22日在大连旅顺塔河湾采捕的沙蜇亲体在育苗室经自然繁育获得,螅状体附着在30 cm×40 cm的聚乙烯波纹板上,培养于1.8 m×1.2 m×0.8 m的水槽中,培养用水取自辽宁省大连市黑石礁近岸,经黑暗沉淀并沙滤后使用,盐度30—32,pH 8.1—8.3,溶解氧大于5 mg/L,前期暂养期间水温随室温自然波动,范围在20—24℃,试验开始前一周螅状体的培养水温约为20℃。前期暂养期间每周投喂一次人工孵化的卤虫 (Artemiasalina)无节幼体,体长约300 μm。
1.2 试验方法
将上述波纹板附着有沙蜇螅状体的区域剪切成约3 cm×10 cm的小片,使每片上螅状体个数为10个,剔除多余的螅状体与足囊。选取总数为30个螅状体的3个小片设为一个试验组的3个平行,置于15 cm×15 cm×3 cm的密封隔离盒中,用智能光照培养箱(GXZ—280B; GXZ—280C)控制试验水温。试验期间用画图兼拍照的方法将每盒中的螅状体按所处波纹板的位置进行编号,统计观察每个螅状体生成的足囊数、足囊萌发情况与横裂生殖过程。
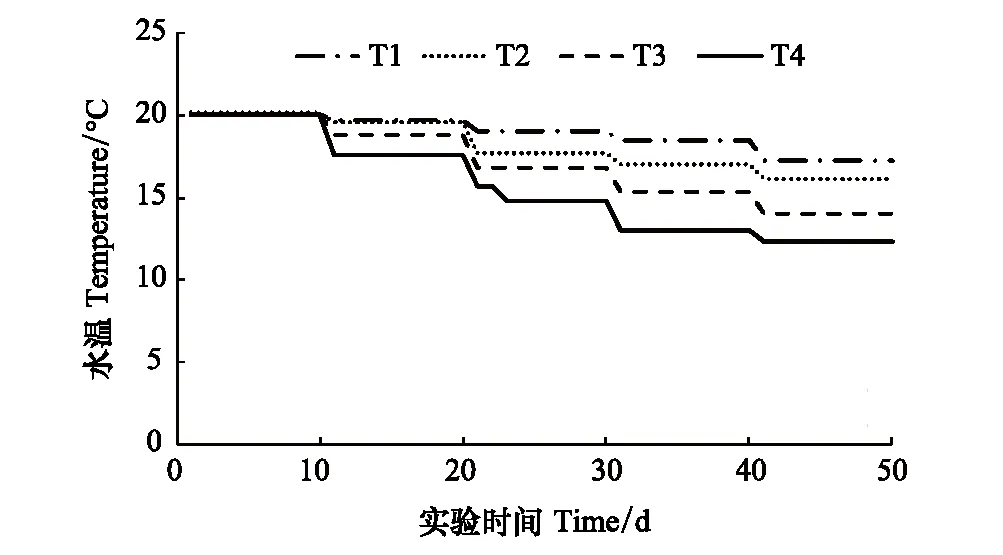
图1 水温曲线图 Fig.1 Temperature curve T1: 秋季高积温组, Autumn high accumulated temperature group; T2: 黄海北部大连星海湾沿岸水域秋季平均降温曲线, Average autumn cooling curve of xinghai coastal waters in north of yellow sea; T3: 人为设置的对照组, Artificial control group; T4: 秋季低积温组, Autumn low accumulated temperature group
根据2010—2016年课题组对自然水温的监测结果,模拟渤海与黄海北部海区秋季不同降温条件,设4个温度变化组(T1、T2、T3、T4),其中,T1为秋季高积温组、T2为黄海北部大连星海湾沿岸水域秋季平均降温曲线、T3为人为设置的对照组、T4为秋季低积温组(图1)。每个温度曲线下设置3种饵料量组,F1组中的每个螅状体投喂1个卤虫,F2组中的每个螅状体投喂3个卤虫,F3组中的每个螅状体均为过量投喂(约每个螅状体投喂10个卤虫),根据Uye等[14]对卤虫C值的测定结果(0.83 μg),估算F1、F2和F3的饵料量分别为0.42、1.25、4.2 μg C 螅状体-1d-1。
结合近岸自然海水盐度历年波动范围(28—33),参考以往海水盐度对沙蜇螅状体影响的研究结果[15],用暴晒24 h的自来水将海水盐度调至28作为试验海水盐度,盐度计型号为PAL—06S。试验期间螅状体每2天投喂一次,用吸有卤虫的吸管对准喂食,保证每个螅状体摄食都达到饵料设置条件,投喂2—4 h后换水。每天用解剖镜(Olympus SZX9)观察螅状体的无性繁殖行为,连续观察50 d。
1.3 指标测定与数据统计分析
形成足囊的螅状体比例
RP=NP/N0
平均每个螅状体形成的足囊数
NPP =P0/Nt
横裂率
STR/%=100 ×NS/N0
足囊萌发率
GRP/%=100 ×Pe/ P0
试验进行第t d的繁殖倍数
RM=Pt/N0
式中,N0为试验开始时螅状体亲体个数,NP产生足囊的亲代螅状体个数,Nt为试验结束时螅状体的个数,NS为发生横裂生殖的螅状体个数,P0为试验期间形成的足囊个数的总和,Pe为试验结束时萌发的足囊个数,Pt为试验进行到第t天时实心足囊个数和萌发螅状体个数的总和。
试验数据用平均数±标准差(Mean ± SD)表示,用Excel 2016和SPSS 25.0软件对数据进行统计分析,采用Duncan’s 多重比较检验组间差异。
2 结果与分析
2.1 秋季降温期水温和饵料量对沙蜇足囊繁殖的影响
2.1.1 水温对沙蜇足囊繁殖的影响
试验过程中,各温度组均有螅状体发生了足囊繁殖行为,T1、T2和T3组形成足囊的螅状体比例较多,组间差异不显著,最高比例出现在T2组,为84.44%,T4组形成足囊的螅状体比例最少,为53.33%,且与其他3组的差异显著。各温度组中每个螅状体形成的足囊数范围为0—6个,形成足囊数最多的螅状体出现在T1组,平均每个螅状体形成的足囊数为2.71个,T2组次之,平均为2.22个,T3与T4组较少,平均分别为1.47与1.36个。如表4,方差分析结果表明,水温对沙蜇形成足囊的螅状体比例和平均每个螅状体形成的足囊数的影响极显著(P<0.001),2指标均有随积温的减少而降低的趋势(表1)。
各试验组中足囊萌发数量均较少,范围为0—3个,平均每个试验组足囊萌发数最多的组是T1,为1.56个,其次为T2和T3,分别为1.22个和1个,T4组最少为0.33个。平均每个试验组足囊萌发率最高的是T3,达6.12%,其次为T1和T2,分别为5.93%和5.21%。T4组的足囊萌发数和足囊萌发率均最少,分别为0.33个和1.18%,与其他3组的差异显著(P<0.05)(表1)。试验结果表明,低积温条件下的足囊萌发也较低。
2.1.2 饵料量对沙蜇足囊繁殖的影响
试验过程中,各饵料组均有螅状体产生足囊,F1和F2组形成足囊的螅状体比例较多,分别为84.17%和80%,F3最少,为48.33%,与F1和F2组的差异显著。各饵料组中每个螅状体形成足囊数最多的螅状体出现在F2组,平均每个螅状体形成的足囊数为2.16个,F1组次之,平均为2.03个,F3组最少,平均仅为1.66个。平均每个试验组足囊萌发数和萌发率均为F2组最多,平均分别为1.83个和7.93%,F1次之,平均分别为1.08个和4.7%,F3最少,平均分别为0.17个和1.2%(表2)。方差分析结果表明,饵料量对沙蜇螅状体足囊繁殖的影响显著(P<0.01)(表4)。

表1 温度对沙蜇螅状体足囊繁殖的影响
同列数据后不同字母表示处理间差异显著(P<0.05),T1: 秋季高积温组, Autumn high accumulated temperature group; T2: 黄海北部大连星海湾沿岸水域秋季平均降温曲线, Average autumn cooling curve of Xinghai coastal waters in north of yellow sea; T3: 人为设置的对照组, Artificial control group; T4: 秋季低积温组, Autumn low accumulated temperature group

表2 饵料量对沙蜇螅状体足囊繁殖的影响
同列数据后不同字母表示处理间差异显著(P<0.05),F1: 每次投喂的饵料量为0.42 μg C 螅状体-1d-1, The amount of food supply per feeding is 0.42 μg C polyp-1d-1; F2: 每次投喂的饵料量为1.25 μg C 螅状体-1d-1, The amount of food supply per feeding is 1.25 μg C polyp-1d-1; F3: 每次投喂的饵料量为4.2 μg C 螅状体-1d-1, The amount of food supply per feeding is 4.2 μg C polyp-1d-1
2.1.3 水温和饵料量交互作用对沙蜇足囊繁殖的影响
各试验组中生成足囊数最多的螅状体出现在T1F1和T1F2组,最多达6个。平均每个螅状体产生足囊数最多的组为T1F2和T1F3,均为2.83个(表3)。形成足囊的螅状体比例最高的组为T2F1组,为100%。双因素方差分析结果表明,尽管温度和饵料量对形成足囊的螅状体比例和平均每个螅状体形成的足囊数的影响达到极显著(P<0.001)或显著(P<0.01)水平,但两者的交互作用对这2个指标影响均不显著(P>0.05)(表4)。
各试验组中足囊萌发数量最多的螅状体出现在T1F2组,达3个,平均每个试验组足囊萌发数最多的组也是T1F2组,达2.62个,平均每个试验组足囊萌发率最高的是T2F2组,达10.25%(表3)。双因素方差分析结果表明,尽管温度和饵料量对足囊萌发数和足囊萌发率达到极显著(P<0.001)或显著(P<0.01)水平,但两者的交互作用对这2个指标影响均不显著(P>0.05)(表4)。
2.2 秋季降温期水温和饵料量对沙蜇横裂生殖的影响
试验期间,只有T2F1组中的螅状体均未发生横裂生殖现象,其他试验组中螅状体虽然有横裂生殖现象,但横裂率均较低,范围在0—23.33%,且每个螅状体放碟数量仅为1—2个,平均每组放碟量范围为0—2.33个(表7)。
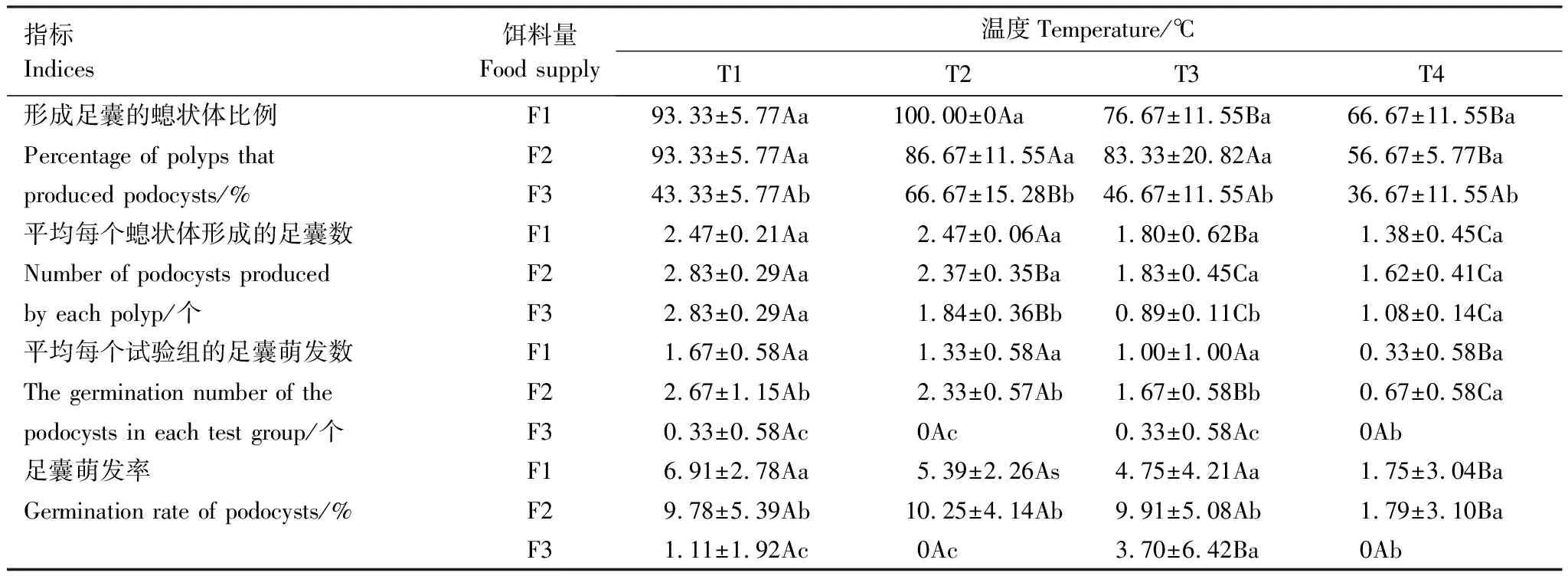
表3 温度与饵料量交互对沙蜇螅状体足囊繁殖的影响
同行数据后不同的大写字母和同列数据后不同的小写字母均表示处理间差异显著(P<0.05)

表4 沙蜇螅状体足囊繁殖的方差分析
2.2.1 水温对沙蜇横裂生殖的影响
各温度组中,T3组中的横裂率和放碟量最多,分别为11.11%和1.22个,T1和T2组次之,T4组横裂率和放碟量最低,分别为6.67%和0.67个(表5),多重比较结果表明各温度组间横裂率和放碟量的差异均不显著(P>0.05),方差分析结果表明水温对沙蜇横裂率和放碟量的影响不显著(P>0.05)(表8)。
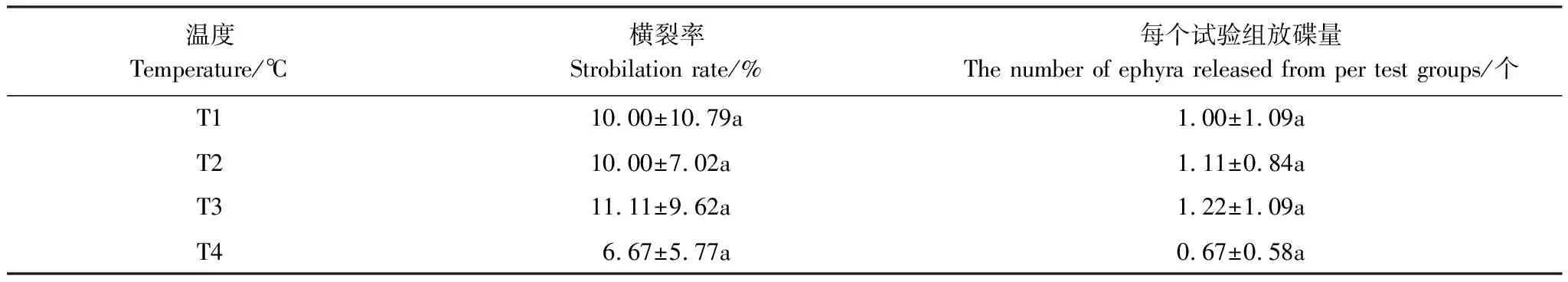
表5 温度对沙蜇螅状体横裂生殖的影响
同列数据后不同字母表示处理间差异显著(P<0.05)
2.2.2 饵料量对沙蜇横裂生殖的影响
各饵料组中,沙蜇螅状体横裂率和每个试验组放碟数均较低,但是随着饵料量的增多,两个指标均有不断增加的趋势,在F3组达到最大,横裂率和每个试验组放碟数分别为14.17%和1.42个(表6)。经组间多重分析结果表明F1和F2差异不显著(P>0.05),但与F3差异显著(P<0.05)。方差分析结果表明饵料量对沙蜇横裂率和放碟量的影响不显著(P>0.05)(表8)。

表6 饵料量对沙蜇螅状体横裂生殖的影响
同列数据后不同字母表示处理间差异显著(P<0.05)
2.2.3 水温和饵料量交互作用对沙蜇横裂生殖的影响
试验期间,发生横裂生殖的螅状体中只有T2F2和T3F2两组中各有一个螅状体释放2个碟状体,其他仅释放一个碟状体。横裂率和每个试验组放碟量均为T1F3组最多,分别为23.33%和2.33个,但与其他组间的差异并不显著(P>0.05)(表7)。经过双因素方差分析结果表明,温度与饵料量的交互作用对沙蜇横裂率和放碟量的影响均不显著(P>0.05)(表8)。
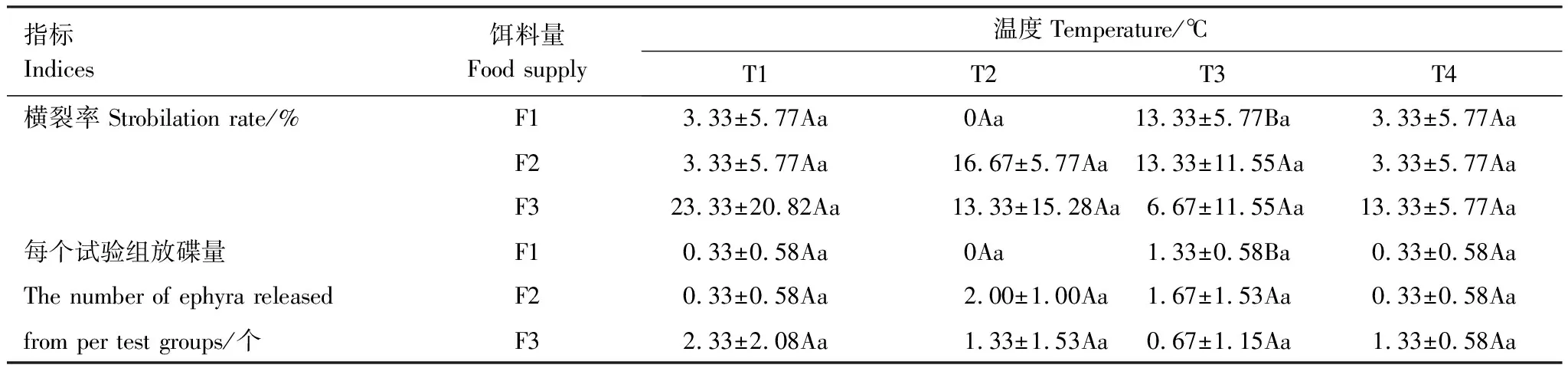
表7 温度与饵料量交互对沙蜇螅状体横裂生殖的影响
同行数据后不同的大写字母和同列数据后不同的小写字母均表示处理间差异显著(P<0.05)
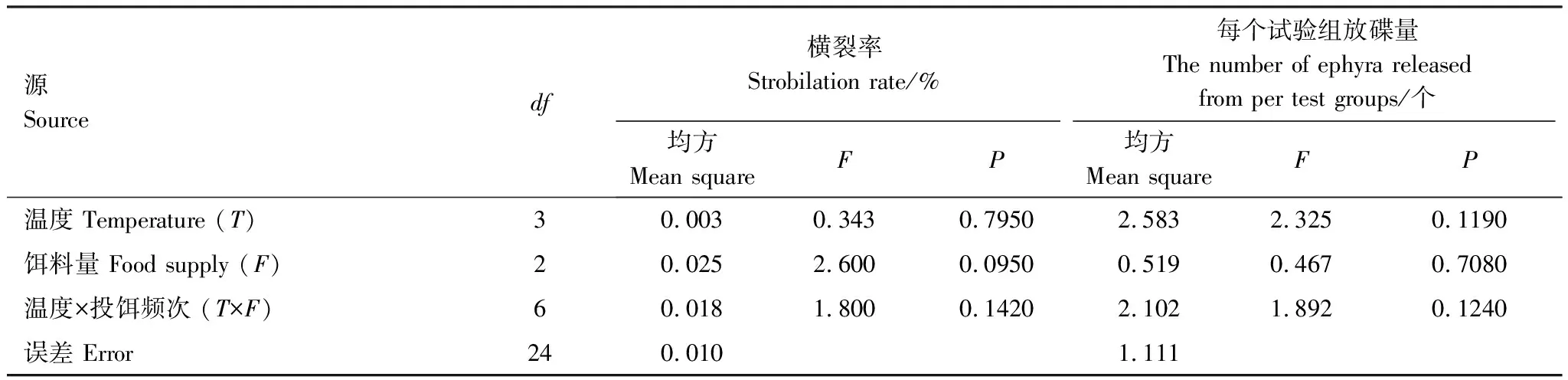
表8 沙蜇螅状体横裂生殖的方差分析
3 讨论
3.1 秋季降温期水温对沙蜇足囊繁殖的影响
沙蜇生活史由有性繁殖的浮游水母体和无性繁殖的底栖螅状体行世代交替组成[3,16],足囊繁殖是沙蜇唯一的螅状体无性繁殖方式[15-16],在螅状体自身种群繁衍过程中起着非常重要的作用[17-18]。在众多影响水母足囊繁殖的环境条件中,温度被认为是最关键的调控因子[19]。我国学者鲁男最早报道了海蜇螅状体在15—30℃范围内,随温度升高,海蜇足囊繁殖能力增强[20]。Kawahara等[21]认为环境条件对足囊繁殖的调控作用是沙蜇暴发的重要原因,并用试验证明了环境异常波动下足囊的萌发量显著高于稳定环境中的。Feng等[22]研究表明,沙蜇足囊繁殖的适宜温度范围为18—25℃。本试验结果表明,水温对沙蜇足囊繁殖的影响极显著,T1温度组足囊繁殖效率最高,且各足囊繁殖的指标均随积温的减少而下降,说明在秋季降温时期积温越高沙蜇螅状体足囊繁殖各指标越高,这与以往的研究结果相一致。
在本试验中,T1温度组的温度下降最慢,T2、T3次之,T4温度下降最快,显然这四个温度组的积温也是由高到低的排列顺序。在图2中,在饵料量相同的情况下,可看出无论是F1、F2还是F3饵料量组,T4温度组的繁殖倍数远低于其他组,同时T3温度组也是低于T1和T2温度组,而T1和T2温度组差距并不大。另一方面,各温度组随着温度的降低,积温增加的速率也逐渐变慢,各试验组繁殖倍数的增幅也是逐渐变慢。总之在秋季降温时,水温下降越慢,即积温越高,螅状体形成足囊的数量越多。

图2 相同饵料量条件下,不同降温条件对沙蜇螅状体繁殖倍数的影响 Fig.2 Effect of temperature on the reproduction multiple of N. nomurai polyps in same food supplyingF1: 每次投喂的饵料量为0.42 μg C 螅状体-1d-1, The amount of food supply per feeding is 0.42 μg C polyp-1d-1; F2: 每次投喂的饵料量为1.25 μg C 螅状体-1d-1, The amount of food supply per feeding is 1.25 μg C polyp-1d-1; F3: 每次投喂的饵料量为4.2 μg C 螅状体-1d-1, The amount of food supply per feeding is 4.2 μg C polyp-1d-1
3.2 饵料量对沙蜇足囊繁殖的影响
多数研究表明,饵料的营养供应是水母进行无性繁殖必不可少的基础条件,多数水母种类在饵料充足的条件下形成足囊,例如,白色霞水母的足囊繁殖随饵料量的增加而增加[23];丰富的营养条件下,海蜇螅状体形成足囊活跃,个体大,易于萌发[24]。有些种类则相反,饵料匮乏是螅状体生成足囊的诱导条件,例如,海月水母(Aureliaauritas.l.)螅状体在高温(26℃)和低饵料供应条件(3.3 μg C 螅状体-1d-1)下形成更多的足囊[11,25]。本文作者曾研究过不同饵料量条件(0.42、1.25、2.08和2.91 μg C 螅状体-1d-1)对沙蜇螅状体在不同恒定水温下足囊繁殖的影响[13],研究表明,在10—27.5℃范围,足囊繁殖速度随水温与饵料量的增加而增加,高温条件下,饵料量的影响作用明显,如饵料不足则螅状体死亡率增加。
本文试验结果中,F2组(1.25 μg C 螅状体-1d-1)的足囊繁殖指标好于F1组(0.42 μg C 螅状体-1d-1),与以往研究结果一致,但F3组(4.2 μg C 螅状体-1d-1)中足囊繁殖的4个指标均明显低于F1和F2组(图3),可能与F3组中的饵料量超过螅状体所需有关,说明饵料量并不是越多越好。在实验过程中也发现,尽管F3组中投喂的卤虫可以都被沙蜇螅状体的触手抓满,但一段时间后有部分卤虫尸体会沉积在附近的片子上,螅状体并不能将所有的卤虫完全消化掉,有些卤虫尸体会黏附在螅状体根部,甚至有些和足囊以及新萌发的幼螅状体黏附在一起,换水时不易清除,这些卤虫尸体长期沉积在螅状体附近,导致螅状体脱落、畸形以及死亡现象频发,且足囊上黏附卤虫尸体也会导致足囊萌发出的螅状体死亡。综合以往研究认为,饵料量对沙蜇螅状体生长及繁殖的影响也是有一定适宜范围的,在适宜饵料量范围内,螅状体足囊繁殖各指标随饵料量的增加而增加,当饵料量超过螅状体所需,沙蜇螅状体的摄食状态及足囊繁殖均受到负面影响,主要原因可能与过量未被消化的饵料残尸有关,过多的能量没有得到有效的转化,导致不利于螅状体生存的水质条件,促使螅状体厌食、脱板、甚至死亡等现象,不利于螅状体种群的繁衍,详细机理还需进一步研究。

图3 相同降温条件条件下,不同饵料量对沙蜇螅状体平均繁殖倍数的影响Fig.3 Effect of food supplying on the reproduction multiple of N. nomurai polyps in same temperature
3.3 有效积温与足囊数量的相关性分析
由于饵料量F3组受过量饵料影响,螅状体有死亡脱板现象,我们选取饵料量F1和F2组数据的平均值与有效积温进行了相关性分析,根据前期研究结果,选取螅状体形成足囊的生物学零度为7.5℃[13],经线性拟合,发现二者具有较好的线性关系(图4)。这表明,沙蜇螅状体形成足囊的数量与有效积温密切相关,螅状体形成足囊的数量随有效积温增加而增多,这也进一步说明了全球变化背景下,积温升高可能导致沙蜇螅状体在越冬前夏秋季的高温期生成更多的足囊,促进诱发翌年水母暴发。
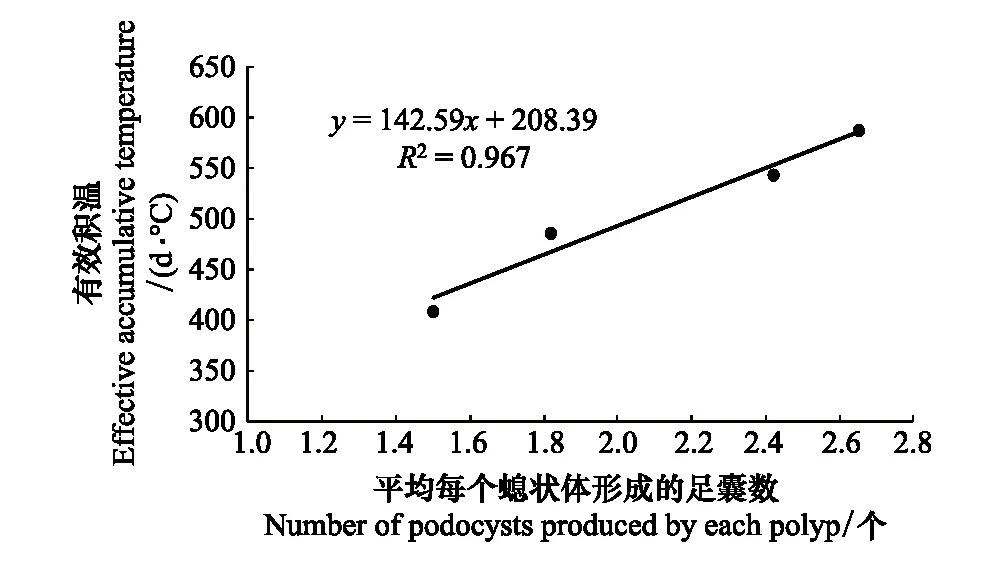
图4 平均每个螅状体生成的足囊数和有效积温的关系 Fig.4 Relationship between Number of podocysts produced by each polyp and effective accumulative temperature
3.4 秋季降温期沙蜇的横裂生殖
横裂生殖是大多数钵水母从螅状体变态发育成水母体所必须经历的一个过程。以往试验研究表明,适宜沙蜇横裂生殖的水温范围在10—18℃[26- 28],但自然海域调查结果表明,沙蜇仅在春季发生横裂生殖释放出水母体[29- 32],未有相关报道发现自然海域中见到沙蜇水母体出现在秋季。本试验过程中,除T2F1组,其他各组中均有螅状体发生了横裂生殖现象,实验结果证明了沙蜇螅状体秋季可发生横裂生殖,但发生横裂生殖的螅状体比例和放碟数量均较少。值得注意的是,本试验对象是沙蜇秋季新繁殖出的当龄螅状体,而在自然海域中,还有越年存活或夏秋季通过足囊繁殖萌发出来的螅状体种群,这部分螅状体因为比有性繁殖的当龄螅状体更早地在海底生长摄食,相对会积攒更多的能量,也许会发生更高的横裂率和释放更多的碟状体,相关研究还有待进一步详细研究。Feng等[27]报道了水温对沙蜇横裂生殖的影响是显著的,本试验结果表明水温和饵料量对沙蜇横裂生殖的影响均不显著,这主要与本试验中的横裂率和放碟数量非常少有关,本试验对象为秋季新繁育出的当龄螅状体,而Feng等进行的试验对象是越年的螅状体,比本试验对象中的螅状体有更多的生长发育时间,进而可以积攒更多的能量用于横裂生殖,这可能是造成差异的主要原因。本试验结果中,积温较少组沙蜇螅状体的横裂率和每个试验组放碟量较少,横裂率和每个试验组放碟量均随饵料量的增加而增大,说明高积温和充足的饵料供应可使螅状体积攒更多能量用于进行横裂生殖。
综上,本试验结果说明,全球变化背景下,积温升高、饵料量小幅增加均可能导致沙蜇秋季有性繁殖的当龄螅状体在越冬前生成更多的足囊,促进诱发翌年水母暴发。
