森林生态系统性状的空间格局与影响因素研究进展
——基于中国东部样带的整合分析
2018-11-02何念鹏张佳慧刘聪聪王瑞丽徐志伟朱剑兴侯继华于贵瑞
何念鹏, 张佳慧, 刘聪聪, 徐 丽, 陈 智, 刘 远, 王瑞丽, 赵 宁, 徐志伟, 田 静, 王 情, 朱剑兴, 李 颖, 侯继华, 于贵瑞,*
1 中国科学院地理科学与资源研究所 生态系统网络观测与模拟重点实验室, 北京 100101 2 中国科学院大学 资源与环境学院, 北京 100049 3 西北农林科技大学 林学院, 杨陵 712100 4 中国科学院西北生态环境资源研究院 遥感与地理信息科学研究室, 兰州 730000 5 东北师范大学地理科学学院, 长春 130024 6 北京林业大学 林学院, 北京 100083
1 性状研究进展简述
性状或功能性状是生物(植物、动物和微生物等)对外界环境长期适应和进化后所呈现出来的可量度的特征[1]。植物光合作用是生态系统初级生产力的重要来源,因此有关植物性状的研究一直以来受到人们高度重视,甚至人们潜意识地将性状研究等同于植物性状或植物功能性状研究[2]。在实际操作过程中,研究人员更多选择能体现植物对环境适应并(或)影响植物生产力的部分性状,如叶片性状(叶片大小、叶片厚度、比叶面积、氮(N)和磷(P)含量等)和根性状(根大小、根长度、比根长、N和P含量等)[3]。20世纪80年代以来,植物性状研究作为生物学、地学和环境科学交叉研究的纽带,成为研究热点,尤其是涉及碳收支0、养分和水分利用的植物性状更是倍受重视,重点探讨了如下几方面的科学问题:①植物性状在不同时空尺度下的变异规律及其影响因素;②植物如何通过多种性状协同(或权衡)来优化生产力;③如何将器官水平测定的传统性状用于解释自然群落构建的机制;④如何从性状角度探讨陆地生态系统对全球变化(气候变化、氮沉降和酸沉降等)的响应与适应。
近年来,植物性状研究的丰硕成果主要体现在如下几个方面:①种内和种间性状变异及其时空变异规律。研究人员从器官-物种-群落-生态系统、控制实验-野外调查-模型模拟等多个尺度和方法对植物性状种内和种间变异进行了探讨,具体指标如植物个体大小、种子、叶和根形态性状、解剖结构、化学元素含量等[4]。②多种性状相互关联关系及其生产力优化机制。经典的研究成果是从资源优化利用效率角度发展了叶经济学谱系(Leaf economic spectrum, LES)、根系经济学谱系(Root economic spectrum, RES)和木材经济学谱系(Wood economic spectrum, WES)[5- 8]。③性状与种间竞争、自然群落结构维持与功能优化间的关系。在物种水平,研究人员较好地建立了植物性状与生产力的定量关系,但如何将个体水平测定的性状与天然群落的结构和功能相结合,仍是该领域的研究热点与难点[9, 10]。④从植物性状角度探讨植物群落或生态系统对全球变化的响应与适应。研究人员利用所测定的部分植物性状数据,直接或结合模型探讨全球变化要素(气候变化、氮沉降和酸沉降等)对生态系统结构和功能的影响[11-12];其中,利用生态化学计量学(Ecological stoichiometry)的基本理论,可从N∶P平衡需求的角度揭示生态系统N和P元素限制状况以及未来全球变化情景下生态系统生产力的变化规律[13- 16]。
整体而言,目前绝大多数植物性状研究主要针对优势物种或个别模式物种,极少充分考虑天然群落物种组成与群落结构的复杂性[8, 17],因此其结论是否适用于复杂的天然群落尚待验证。即使是著名的世界植物属性数据库(TRY-Plant trait database),也是以收集公开发表数据为基础建立的,在大多数植物群落中仅有为数不多的物种数据。此外,TRY数据库的数据多集中在植物叶片,非常缺乏叶-枝-干-根-种子等配套的植物性状数据。因此,未来的植物性状研究应该向更系统的指标体系、仿真自然群落的模式开展。近几年,国外科学家大量提倡从物种推演到群落的观点,但都面临缺乏一个系统的实测数据库来支持研究方法和理论发展的难题[1, 12]。因此,非常有必要开展系统性调查与测试,并以此为基础探讨和发展从物种水平到群落水平的性状研究的方法论和基本理论。然而,纵观国内外相关研究,一直缺少贴近天然生态系统(植物、动物、微生物、土壤等)的性状调查模式,来引导跨学科、系统性、集成式调查和测定;该调查模式一旦被确立并被广泛采用,将会提供各种系统性的性状调查数据。
本文介绍了在国家基金委重大项目资助下的中国东部南北样带(North-South Transect of Eastern China, NSTEC)森林生态系统性状(植物、微生物、土壤)的综合调查方案及其核心研究结论,从热带雨林至寒温带针叶林3700 km样带上,选择了9个地带性森林生态系统开展详细的调查和测定工作,同时探讨了森林生态系统的性状研究——如何从物种水平拓展至群落水平?如何建立天然森林群落的性状与功能的定量关系?对基于系统性调查数据的性状研究也进行了展望。
2 中国东部南北样带(NSTEC)典型森林生态系统在性状研究中的潜在意义
根据国际地圈生物圈计划 (International Geosphere-Biosphere Program, IGBP) 的要求,全球变化陆地样带都是由一系列沿着某种具有控制陆地生态系统结构、功能和组成、生物圈-大气圈间的痕量气体交换和水分循环的全球变化驱动力带状区域组成,其长度应不小于1000 km,以确保能够覆盖气候和大气模式以及决策尺度,并有足够宽度(数百公里)以涵盖遥感影像范围[18-19]。中国东部南北样带(NSTEC)于2005年被IGBP确认为第15 条标准样带;它的主体从中国东部108—118°E 沿经线由海南岛北上到40°N,然后向东错位10°,再由118—128°E 往北到国界(图1)。NSTEC具有明显的热量梯度与水热组合梯度,覆盖了北半球主要森林类型,形成了世界上独特且完整的以热量驱动的植被连续带[20-21]。
NSTEC从南到北覆盖了热带山地雨林、亚热带阔叶常绿林、温带阔叶落叶林、温带针阔混交林和寒温带针叶林[22- 24];其土壤理化特性随着纬度升高发生显著变化,从低有机质含量的热带红壤逐渐转变为高纬度地区高有机质含量的棕壤[25]。由于该样带具有明显的水热梯度特征,提供了一个理想的天然实验室,也有利于探讨大尺度下各种生态系统性状(植物、微生物和土壤)的变异规律和影响因素,并可采用空间替代环境要素变化的模式探讨生态系统性状的响应与适应机制。由于NSTEC覆盖了北半球几乎所有的主要森林类型,因此其研究结论具有全球意义[20]。研究于2013年沿NSTEC选择了9个地带性森林生态系统(均为原生林)开展系统性状调查研究,从南到北分别是尖峰岭(热带山地雨林)、鼎湖山(南亚热带常绿阔叶林)、九连山(亚热带常绿阔叶林)、神农架(北亚热带常绿阔叶林)、太岳山(暖温带落叶阔叶林)、东灵山(暖温带落叶阔叶林)、长白山(温带红松阔叶混交林)、凉水(温带红松阔叶混交林)和呼中(寒温带针叶林)(图1)。调查样地跨越33个纬度(18.7°N—51.8°N)和14个经度(108.9°E—123.0°E)。 研究区域年均温(MAT)最低为-3.67 ℃,最高达到23.15 ℃;年均降水(MAP)最低为472.96 mm,最高为2265.80 mm(表1)。
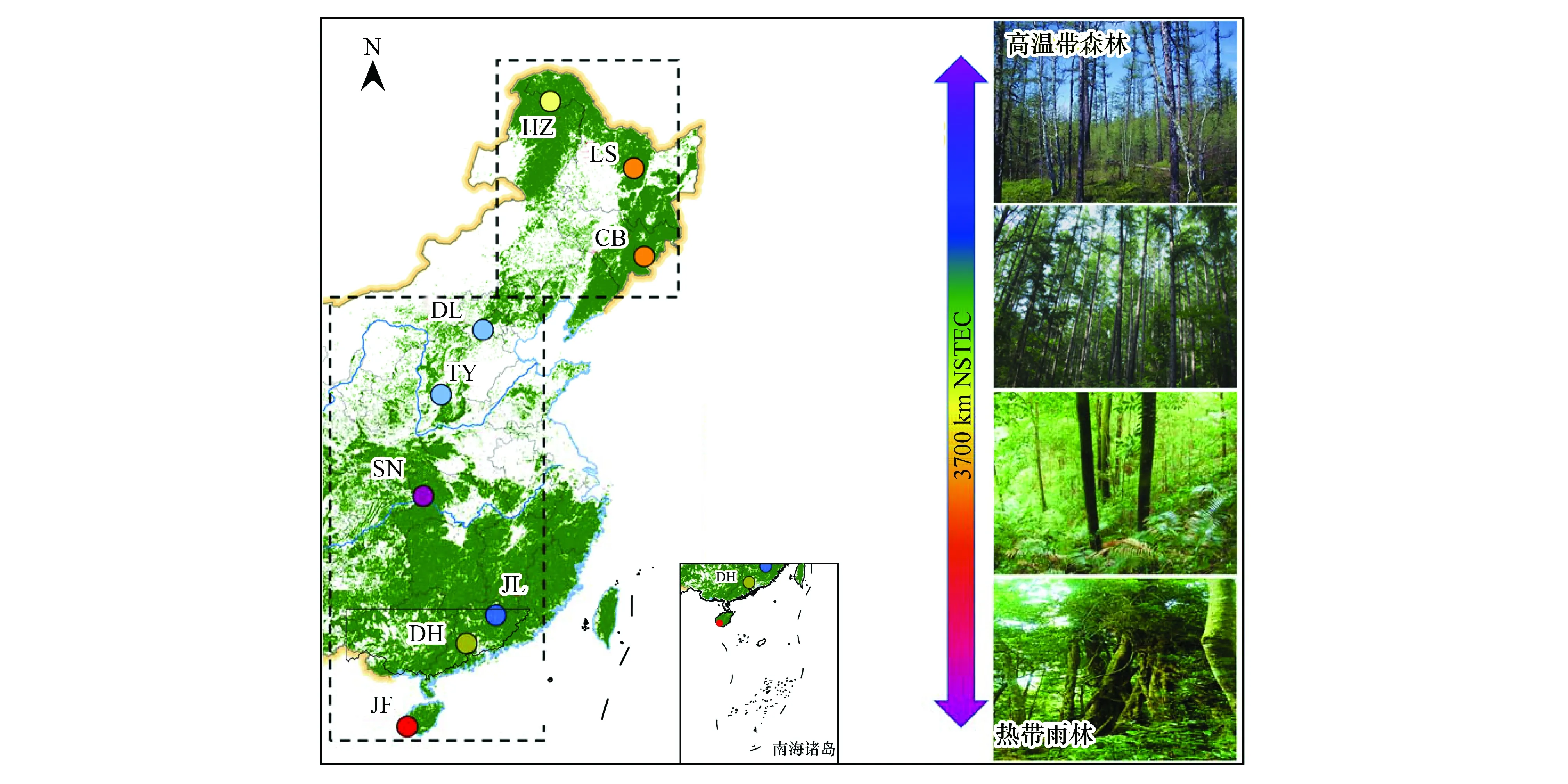
图1 中国东部南北样带及9个地带性森林生态系统空间分布Fig.1 Picture for the North-South Transect of Eastern China (NSTEC) and the nine typical forest ecosystems JF, 尖峰岭; DH, 鼎湖山; JL, 九连山; SN, 神农架; TY, 太岳山; DL, 东灵山; CB, 长白山; LS, 凉水; HZ, 呼中
3 如何基于NSTEC典型森林生态系统开展系统性的性状调查
如何开展系统性的性状调查,使其更加接近天然森林生态系统?为此,引入了图2的模式。以便所测定的指标体系能更接近复杂的天然森林生态系统,实现从物种水平(或器官水平)尺度上推至群落水平,探讨自然群落水平(或生态系统水平)性状与功能的定量关系。
表1中国东部南北样带9个典型森林生态系统气候、植被和土壤基本状况
Table1Propertiesofclimate,vegetation,andsoilinthenineselectedtypicalforestecosystemsalongtheNorth-SouthTransectofEasternChina(NSTEC)

站点Site经度(°E)Longitude纬度(°N)Latitude海拔/mAltitude年均温/℃Mean annual temperature年降水量/mmMean annual precipitation植被类型Vegetation type土壤类型Soil type尖峰岭108.918.780923.152265.80热带山地雨林砖黄壤 鼎湖山112.523.224021.831927.00南亚热带常绿阔叶林砖红壤 九连山114.424.656218.221769.93亚热带常绿阔叶林红壤 神农架110.531.315108.501446.71北亚热带常绿阔叶林黄棕壤 太岳山112.136.716685.98644.38暖温带落叶阔叶林褐土 东灵山115.440.09726.55539.07暖温带落叶阔叶林棕壤 长白山128.142.47582.79691.00温带红松阔叶混交林暗棕壤 凉水 128.947.24010.01648.34温带红松阔叶混交林暗棕壤 呼中 123.051.8850-3.67472.96寒温带针叶林灰色森林土
土壤类型鉴别参照了中国土壤分类系统(1993)
2013年 7—8月生长旺季,沿NSTEC样带选择了9个地带性森林生态系统为调查对象(图1和表1),为降低人为干扰,实验样地均设置于自然保护区内或野外生态站的长期监测样地内,在每类森林生态系统的典型地段,分别设置4个30 m × 40 m乔木调查样方、每个乔木样方内分别设置2个5 m × 5 m灌木调查样方和4个1 m × 1 m草本调查样方(图3),按中国生态系统研究网络(CERN)调查规范,采用每木检尺详细调查乔、灌、草的群落结构特征,为后续采用生物量加权实现物种-功能群-群落尺度性状数据拓展提供数据支撑[26]。

图2 典型森林生态系统性状调查的逻辑框架Fig.2 Logical frame of the investingated traits (plant, soil microbe, and soil) in typical forest ecosystems
植物样品采集:①乔木样品采集:选择样地内(或周边)长势良好的乔木植株,采用吊塔+人工爬树+高枝剪相结合模式获得顶层向阳树枝;随后手工摘取无病虫害的、完全展开的健康叶片,每个样地分别形成叶或细枝混合样品。在叶片样品采集的同时,采用生长锥采集树芯样品;同时,沿侧根挖掘出完成的细根根系(从1级到5级根[27])。②灌木样品采集:对每个灌木物种,在每个样地选择1—3株健康植株,利用高枝剪或枝剪截取冠层顶端枝条,分别获得叶和枝条的混合样品;同时采用挖掘法获得灌木根系。③草本样品采集:在每个样地内,每个草本物种选取5—30株,采集地上部分(叶、茎)和根系。此外,在草本样方内收集地表凋落物混合样品。
土壤样品采集:利用6 cm直径的土钻,分0—10 cm、10—30 cm、30—50 cm、50—100 cm四层进行土壤取样。在每个样地取30—40钻土壤样品,获得每层土壤样品>10 kg;随后,大约4 kg鲜土壤快速冷藏于-20 ℃冰柜,其他样品遮荫风干。
此次性状调查的系统性体现在:①协同调查了群落结构、群落内每个物种性状、微生物属性、土壤属性、土壤有机质周转及其温度敏感性等(表2)。②调查了样地内出现的所有高等植物种类(乔、灌、草)。比如在呼中寒温带针叶林调查了95个物种、在尖峰岭热带山地雨林调查了208个物种,9个森林生态系统共调查了1177种植物。③开展了乔、灌、草的叶-枝-干-根性状配套调查。④叶片性状包括常规形态、气孔[23, 28]、叶片结构[29, 30]、化学元素[24, 26, 31, 32]、叶绿素含量[33, 34]、非结构性碳水化合物[35- 37]、叶片热值等[22, 38]。⑤采用多种方法系统地测定了土壤微生物组成和酶活性[39]。⑥测定了土壤有机质含量及其组分[25];⑦开展了土壤有机质碳氮矿化及其温度敏感性研究[40- 42]。此次调查涵盖了生态系统植物群落到土壤环境的整体系统性、植物群落乔木、灌木到草本的群落系统性以及植物个体叶、枝、干到根的个体系统性。然而,针对图2中森林生态系统冠层界面的功能参数,不同于传统叶片水平测定的功能参数,更多是指生态系统尺度的生产力和水分利用效率等,主要是通过涡度相关技术来进行测量或模型模拟;它并非此次野外调查数据的主要内容,该数据共享于中国通量网(ChinaFlux)。

图3 森林群落调查样方的设置 Fig.3 Design of investigating plots for forest communities乔木样方面积30 m × 40 m,其中还设置2个5 m × 5 m 灌木样方(S1和S2)和4个 1 m × 1 m 草本样方(H1—H4)
除揭示了多个性状的空间变异规律及其影响因素外,基于上述系统性的调查数据,发展了基于植物群落结构和生物量加权将植物性状从器官推导至群落的新方法(公式1和2),并率先探讨了部分植物性状与天然森林群落初级生产力(Gross Primary Productivity, GPP)、净初级生产力(Net Primary Productivity, NPP)、净生态系统生产力(Net Ecosystem Productivity, NEP)、水分利用效率(Water Use Efficiency, WUE)等的定量关系,推动了天然森林群落性状与功能关系的新认识,并提供了可借鉴的方法学。其中,公式1适于以质量(或重量)为基本量纲的性状指标,而公式2适用于以叶面积或厚度为量纲的指标。通过公式1和2,能把绝大多数器官水平测定的植物性状参数转化为群落水平,并实现以单位面积为基础转化,可为探讨性状与天然森林群落功能的关系奠定坚实的基础,因为大尺度天然森林群落生产力和水分利用效率等功能参数,大多都是以单位土地面积为基础(通量观测、遥感观测或模型模拟)。
(1)
(2)
式中,Traitcomm是群落水平性状值;trait是物种水平性状值;n是森林群落中的物种丰富度;j=1, 2, 3, 4分别代表了叶、枝、干和根;Biomassij是指每个器官生物量在整个群落生物量中所占的比例;LAIi是指植物群落中i物种的叶面积指数[23, 28]。
表2中国东部南北样带森林生态系统的性状指标体系与测定方法
Table2TheparametersandmeasuredmethodsfordifferenttraitsinforestecosystemsalongtheNorth-SouthTransectofEasternChina(NSTEC)

性状参数Parameters of trait测试方法/工具Measurement method or instrument 植物性状叶片形态(叶片大小, 厚度, 比叶面积)扫描、游标卡尺和电子天平气孔(气孔大小, 密度, 相对面积)印迹法/高倍电子显微镜解剖结构(叶片厚度, 海绵组织厚度, 栅栏组织厚度)固定-染色-切片/电子显微镜 多元素含量(碳, 氮, 磷, 钾, 钙, 镁, 硫, 铝, 锰, 铁, 钠, 锌, 铜, 铅, 镍, 钴)元素分析仪叶绿素含量(叶绿素a, 叶绿素b)乙醇法非结构性碳水化合物(可溶性糖, 淀粉)比色法热值燃烧法枝多元素含量(碳, 氮, 磷, 钾, 钙, 镁, 硫, 铝, 锰, 铁, 钠, 锌, 铜, 铅, 镍, 钴)元素分析仪干多元素含量(碳, 氮, 磷, 钾, 钙, 镁, 硫, 铝, 锰, 铁, 钠, 锌, 铜, 铅, 镍, 钴)元素分析仪根形态(根直径, 长度,体积, 比根长)根序法多元素含量(碳, 氮, 磷, 钾, 钙, 镁, 硫, 铝, 锰, 铁, 钠, 锌, 铜, 铅, 镍, 钴)元素分析仪土壤微生物性状微生物微生物生物量及群落组成氯仿熏蒸法+磷脂脂肪酸标记法微生物群落代谢活性Biolog Ecoplate微平板培养法微生物细菌多样性高通量测序酶酶活力微孔板荧光法土壤性状物理性质土壤质地(沙粒, 粘粒, 粉粒)激光粒度仪土壤含水量烘干法化学性质多元素含量(碳, 氮, 磷, 钾, 钙, 镁,铝, 锰, 铁, 钠, 锌, 铜, 铅, 镍, 钴)元素分析仪土壤pHpH计有机碳组成土壤有机碳重铬酸钾氧化法易氧化有机碳高锰酸钾氧化法腐殖酸碳萃取法腐殖质碳萃取法土壤有机质周转碳矿化碳矿化速率培养+连续自动测试碳矿化温度敏感性培养+连续自动测试氮矿化氮矿化速率培养+破坏性测试氮矿化潜力原位与培养N15双标氮矿化温度敏感性培养+破坏性测试
4 森林植物性状的空间变异及其影响因素
4.1 叶片性状
4.1.1 叶片形态性状的纬度格局及其影响因素
叶片形态性状可以很好地表征植物对外界环境的响应与适应,备受科研人员的广泛关注[8, 17, 32]。通过对NSTEC样带9个典型森林生态系统1100多种植物叶片性状数据的分析,发现叶片形态属性的纬度变异性在物种水平和群落水平上存在差异[43]。在物种水平上,随纬度增加,植物叶片大小(LA)、叶片厚度(LT) 和叶片干物质含量(LDMC)均显著降低,叶片比叶面积(SLA)增加;然而,LA和LT沿纬度梯度的变化趋势却较弱(R2=0.03-0.04)。然而,利用物种水平数据+群落结构数据的尺度拓展方法(公式1或公式2),发现在群落水平上叶片大小(CWM-LA)和比叶面积(CWM-SLA)随着纬度增加呈现先增加到最大值而后降低的趋势。群落叶片厚度(CWM-LT)随纬度增加而增加,群落叶片干物质含量(CWM-LDMC)变化趋势与之相反。相关研究结果表明:在探讨植物性状大尺度空间变异规律和影响因素时,物种水平算术平均的传统方法与结合群落结构和相对生物量(或叶片面积)的权重加权法存在很大的差异,有些结果甚至截然相反;至于物种水平算术平均法vs.群落结构和生物量加权法在探讨植物性状格局及其影响因素的优缺点,会因研究人员背景或研究内容的变化有所变化。
利用巢式方差分析,将物种水平上叶片形态属性的纬度变异性区分为样地内和样地间的变异性。结果表明,物种水平的叶片形态属性38.2%—50.9%的变异性存在于样地内部共存物种之间,只有16.0%—37.2%的变异性能够被样地之间的差异(包括气候和土壤因子的差异)所解释。此外,与SLA和LDMC相比,LA和LT中较多的变异性存在于样地内部(> 49%)。植物功能型是LA、LT、SLA和LDMC空间变化的主要影响因素,能够解释物种水平的叶片属性19.43%—41.57%的变异性,而环境因素仅仅解释了总变异的1.65%—9.13%。这和全球及区域尺度上植物功能型对叶片形态、养分和生理属性空间变异的解释程度大于环境因素的结论一致[44, 45]。气候和土壤N含量是影响群落水平的叶片形态属性沿纬度变化的主要因素,气候和土壤养分条件作为主要的环境滤镜,通过改变群落内的物种组成和群落结构,进而决定了叶片形态属性的群落加权平均值在不同环境条件下的差异[43]。
研究揭示了森林植物叶片形态性状的纬度变异规律与影响因素,但在以下几方面还需探讨:①由于种间变异解释了叶片功能性状的大部分变异,物种自身系统发育是否对叶片功能性状产生影响以及产生多大影响?②从热带雨林到寒温带针叶林群落的构建机制是否存在差异?③叶片形态性状如何影响天然森林生态系统的功能?
4.1.2 叶片气孔性状的纬度格局及其影响因素
气孔是植物体内水分和二氧化碳与外界环境进行交换的主要通道,其形态和行为不仅可体现植物自身对环境的适应对策,对全球大气碳循环和水循环也具有深远影响[46]。迄今为止,国内外科学家有关气孔性状的研究大多集中在特定区域的少数物种[46- 48];在区域尺度上,天然森林群落的气孔形态及其对环境变化的响应和适应依然模糊不清。
采用印迹法和高倍电镜对NSTEC样带9个地带性森林1100多种植物气孔性状进行了调查,具体指标包括气孔密度(SD)和长度(SL)。结果表明:SD和SL在物种水平上的纬度格局较弱,但在群落水平上随着纬度的升高而明显降低,这种变异是由温度和降水驱动的。在物种水平上,SD与SL呈现负相关关系;在群落水平上,SD与SL却表现出显著的正相关。表明了群落气孔属性和物种个体气孔属性两者对环境变化的适应机制存在着差异。物种水平上SD与SL之间的权衡关系是植物个体对环境变化的适应进化的结果,而群落水平上的气孔特征则更多受到群落构建过程的影响。在群落水平上的SD能够解释NPP空间变异的 51%[23]。为了探究气孔性状的影响因素及其与生态系统水分利用效率的联系,提出了气孔面积指数(Stomatal area fraction,f,%)[28],并利用群落结构和生物量加权法计算了每个植物群落的气孔面积指数(f),发现f随着纬度的增加表现出先升高后降低的趋势,干旱度指数是主要的驱动因素。同时还发现f与生态系统WUE显著正相关,表明植物群落通过增大f来优化生态系统的水分利用效率。相关研究在区域尺度上为气孔性状影响生态系统水分利用效率提供了直接证据,提高了人们对气孔性状适应环境的认识。
通过对NSTEC植物气孔性状的系统性调查,填补了大尺度气孔数据的空白[28]。然而,目前有关叶片气孔性状的分析仍然主要集中在个体、功能群和群落的格局及影响因素;气孔作为最重要的植物叶片功能性状之一,其如何在群落维持和构建机制上发挥重要的作用仍需进一步探讨。
4.1.3 叶片解剖结构的纬度格局及其影响因素
叶片解剖结构(上下表皮细胞、栅栏组织、海绵组织等)具有较大的可塑性,在不同的选择压力下会形成各种适应类型[49, 50]。植物同化二氧化碳的同时也伴随着蒸腾作用带来的水分丧失,一方面植物要减少水分丧失,达到高的水分利用效率,另一方面实现碳的最大化获取,充分发挥光合作用而大量损失水分,导致了低水分利用效率[51]。叶绿素主要分布在栅栏组织和海绵组织中,因此,解剖结构是叶片进行光合作用过程中影响光合效率和气体交换的主要部分[52]。随着环境梯度的变化,植物会如何调整叶片各个解剖结构之间的比例关系,叶片解剖结构是否影响着自然生态系统的生产力和水分利用效率,值得深入探讨。
利用固定-染色-切片技术,对NSTEC的1100多种植物叶片的解剖结构进行了测定[29, 30],具体指标包括上表皮厚度(AD)、下表皮厚度(AB)、叶厚度(LT)、栅栏组织厚度/海绵组织厚度(PT/ST)、栅栏组织厚度/叶片厚度(CTR)以及海绵组织厚度/叶片厚度(SR)。实验结果表明:在物种水平上AD、AB、LT、PT/ST、CTR、SR的均值分别是11.59 μm、8.65 μm、119.66 μm、64.84%、26.82%、48.81%。在功能群水平上,乔木、灌木和草本的6个解剖结构指标之间具有显著的差异 (P<0.05)。利用群落结构和生物量加权推导至群落水平后,AD、AB、LT、PT/ST、CTR、SR 均值分别是8.68 μm、3.83 μm、111.55 μm、83.43%、20.89%、28.89%。从物种、功能群到群落水平,解剖结构指标均具有显著的纬度格局,但是不同研究层次,解剖结构指标随纬度的变化趋势有较大差异。解剖结构受到温度、降水共同影响,并与干旱指数显著相关。在群落水平,解剖结构与GPP和WUE具有显著相关性,其中GPP随着SR的增加而增加,随着PT/ST的增加而降低;WUE随着PT/ST的增加而增加,而随着SR的增加而降低[30]。
已有研究报道了从热带至寒温带森林的植物叶片解剖结构数据[30],探究了叶片解剖结构在物种、功能群和群落水平上的纬度分布格局及其影响因素,在区域尺度上建立了叶片解剖结构与生态系统生产力和水分利用效率的联系,为揭示天然群落中性状与生态功能的联系提供了新证据。未来应加强系统进化对叶片解剖结构的影响研究,从长期适应与进化角度探讨叶片解剖结构的生态学意义。
4.1.4 叶片叶绿素含量的纬度格局及其影响因素
光合作用是植物生长的重要能量来源和物质基础。植物光合反应可分为原初反应、电子传递和光合磷酸化、碳同化三个过程。叶片中的叶绿素a(Chlorophyll a, Chl a)和叶绿素b(Chlorophyll b, Chl b)是原初反应的必要条件;然而二者功能却又有所不同。大多数Chl a和所有Chl b都能够吸收太阳光(Chl a主要吸收红橙光,Chl b主要吸收蓝紫光),但是只有少量处于激发态的Chl a能在原初反应中传递电子,并将光能转化成电能,从而为电子传递和光合磷酸化以及碳同化做准备。据此推测:叶绿素总量(Chlorophyll content, Chl a+b)以及Chl a 与Chl b 比值(Chl a/b)直接或间接地影响植物的光合效率,是反映植物光合能力的重要指标[53]。鉴于叶绿素对植物光合作用的重要意义,科学家们对叶绿素荧光[54]、叶绿素的合成分解[55, 56]等进行了大量研究。然而,目前对于叶片叶绿素含量的研究大多停留在区域优势物种或某物种叶绿素的影响因素[57, 58]等,对于大尺度群落叶绿素的研究鲜有报道。
利用经典的乙醇法提取,测定了NSTEC上9个地带性森林群落内1100多种植物叶片叶绿素含量(Chl a、Chl b、Chl a+b、Chl a/b),分析发现叶绿素在物种间和不同生活型间都存在显著差异;在物种水平时,叶片叶绿素含量仅存在较弱的纬度格局。进一步分析发现,叶绿素几乎不受系统发育、气候、土壤的影响,较大的种间变异是导致物种水平叶绿素微弱纬度格局的重要原因。然而,当利用群落结构和生物量数据加权法拓展后,发现群落水平的叶片叶绿素含量随纬度的升高而降低;进一步分析发现Chl与GPP显著正相关(R2=0.32),即Chl一定程度上能够反映生态系统GPP,但其作为单一性状对GPP空间变异的解释度远低于先前人们的预期。相关研究从群落的角度揭示了自然群落叶绿素纬度变异规律、影响因素及其与GPP的定量关系,为植物功能性状与功能的研究提供了一个范例[33-34],也为生态模型构建与优化提供了理论依据和重要参数。
近年来,许多学者用叶绿素含量建立了很好的光合模型,并且发现用叶绿素代替光合效率比传统上用N含量代替光合效率具有更好的效果[53]。然而,类似模型都是将对少数物种的实验结果直接应用于复杂的自然群落,需要来自群落水平的性状与功能数据的实验验证;此外,还应考虑不同区域环境异质情况下,叶绿素与GPP定量关系的变异规律等。
4.1.5 叶片非结构性碳水化合物的纬度格局与影响因素
碳水化合物是植物光合作用的重要产物,常分为结构性碳水化合物和非结构性碳水化合物(Non-structural carbohydrates, NSC)。NSC主要用于植物生理代谢活动,其含量的变化很大程度的影响着植株代谢和生长过程、植物碳吸收与碳消耗之间的平衡关系和植物对外界环境变化的重要适应策略[59]。在垂直地带性上,温度驱动机制下的林线形成(温度对植物叶片非结构性碳水化合物的影响)有两种对立的生物生理假说——碳限制假说[60-61]和生长限制假说[62-63]。然而,在大尺度的自然森林生态系统中,如沿着不同气候带(从热带到寒温带、随着温度的降低),植物叶片NSC的变化趋势会支持何种假说,不得而知。
以NSTEC样带上9个地带性森林生态系统1100多种植物叶片为对象,测定了其可溶性糖和淀粉含量 (NSC=可溶性糖+淀粉)。在物种水平,植物叶片可溶性糖、淀粉和NSC的变化范围分别为 6.48—218.45 mg/g、7.56—200.41 mg/g 和19.47—379.72 mg/g,平均值分别为45.70、47.50 和93.20 mg/g。此外,不同生活型间叶片NSC差异显著,草本叶片可溶性糖、淀粉和NSC最高,分别为49.89、60.12和110.00 mg/g;落叶乔木叶片可溶性糖、淀粉和NSC含量显著高于常绿乔木叶片(P<0.05);针叶乔木的叶片可溶性糖、淀粉和NSC含量显著高于阔叶乔木叶片。植物叶片可溶性糖、淀粉和NSC含量具有明显的纬度地带性分布规律,沿纬度带从南到北呈递增趋势。然而, 叶片可溶性糖和淀粉的分配策略(比值)沿纬度没有明显的变化趋势,与温度和降水之间也无显著相关性[35- 37]。
相关研究探讨了植物叶片非结构性碳水化合物及其组分的空间格局及控制因素,对了解气候变化对树木生理生态过程的响应和适应机制具有一定的意义。未来还应加强NSC在不同器官间的分配特征研究,应考虑多种因素如气候、植被和土壤对植物叶片NSC的影响机制;此外,在复杂的天然森林群落中,系统发育、光合途径对植物叶片NSC的影响也需要进一步阐述。
4.2 细根性状的纬度格局及其影响因素
细根在植物水分和养分吸收中具有重要作用,由于野外取样和测定难度较大,有关细根的研究仍然不足[27];目前关于细根的功能性状之间是否存在类似于叶经济学谱的规律仍存在激烈争论(图4)。根系经济学谱理论(RES)认为:细根中与资源获取和资源贮存相关的性状之间存在权衡关系。在RES的一端是快速回报的物种,具有低的根直径(RD)、高的比根长(SRL)和根氮含量(RN),同时具有高的根系呼吸速率和短的寿命;相反的特征存在于慢速回报的物种内。然而,根的多维度假说认为,根特征之间不存在唯一的权衡关系,(与叶片相比)根系特征受到更为复杂的环境限制、具有更高的系统发育保守性,同时还会受到菌根侵染的影响。此外,由于细根的划分标准存在差异,也阻碍了对细根性状变异及其相互关系的深入研究;近年来研究提出和发展了“根序法”和“功能划分法”,极大推动了细根的研究工作[27, 64]。
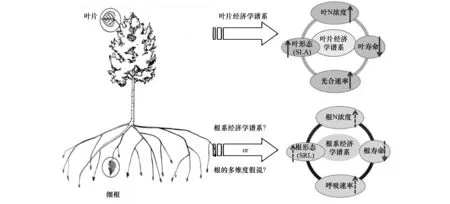
图4 细根功能性状之间关系的相关假说Fig.4 Main hypotheses on the traits of fine foot
在NSTEC的9个地带性森林中选择了群落内的常见物种(包括乔木、灌木和草本植物),开展了较系统的一级根的功能性状测定[65]。根据主成分分析(PCA),在物种和群落水平上均发现了2个主成分轴(PC1和PC2)。其中,PC1主要与群落水平比根长(CWM_SRL)和根直径(CWM_RD)相关,代表了根厚度的变化,PC2主要与根养分性状相关。此外,不同气候区域的细根功能性状存在差异。亚热带地区的植物细根倾向于具有细且长的根,而北方针叶林地区的植物根系具有相反的功能性状。一些温带森林(如长白山)中的物种具有高的氮含量和低的碳氮比。
细根形态和化学指标之间的独立性支持了细根的多维度性假说。PC1主要代表了根形态性状(如RD和SRL)的变异性;PC2与根的养分性状相关,这反映了细根从土壤中吸收养分的重要性。同时,在物种水平上与根形态性状相比,根的养分性状表现了较低的系统发育保守性和对土壤环境更高的可塑性[66]。进一步分析发现:系统发育作用能够解释木本和草本植物51.4%和49.4%的变异性;并且大多数变异性发生在分支水平上(clade),反映了不同进化时期系统发育分支的根性状的巨大差异。与物种水平不同,群落水平的细根性状变异性受到了环境滤镜和生物之间相互作用的双重影响,这些因素最终决定了群落内的物种组成和功能性状分布[67]。例如,在群落水平上,随着年均温增加,PC1先降低,而后略微增加;相反的格局表现在PC2和土壤磷含量的关系中。
目前,大多数研究只涉及根的形态和养分性状,而对于更能准确地刻画根系功能的解剖结构,尤其是涉及真菌侵染方面的性状,以及和根系构建特征相关的性状(如分支强度)还有待加强。部分研究已经表明细根的菌根侵染率和分支强度是细根资源吸收策略的重要组成部分[68, 69],因此,未来需要将菌根特征和根的构建特征考虑在内,扩展根性状的研究维度,深入探究根系对环境的适应策略。
4.3 根-叶性状关联性
叶片与细根分别是植物体地上和地下最活跃的器官,理论上其性状应存在着密切联系[70];因此,理解叶片和细根功能性状之间的关系,能够为植物资源利用策略和生态系统过程模型优化提供重要依据。然而,目前对于植物地上/地下器官功能性状之间的关联关系存在较大争议。一种假说认为,植物各器官功能性状之间的关系是一维的。外界环境的选择压力和生物物理限制共同导致了植物功能性状聚集在从“快”到“慢”的策略轴上(即资源获取-保守策略)[70]。另一种假说则认为,植物不同的器官由于所受到的环境限制和行使的功能均存在差异,因而会独立进化来适应不同的环境条件。因此,植物不同器官功能性状之间的关系是多维度的[66, 71],这有利于提高群落内物种共存和生态系统结构与功能的稳定性。
利用在NSTEC获取的木本和草本植物的叶片和细根性状,根据根和叶养分获取和形态特征对应角度,选择了5组性状来探讨根-叶性状关系。5组分别为:叶片厚度 vs 细根直径、比叶面积vs 比根长、叶片组织密度vs细根组织密度、叶片碳含量vs 细根碳含量、叶片氮含量vs 细根氮含量。实验结果表明:形态特征之间的关系取决于是否考虑系统发育因素,而叶片氮和细根氮含量之间存在紧密的正相关关系[72]。因此,根养分特征可以作为连接不同植物器官以及植物与环境之间关系的桥梁。叶片和细根之间形态特征的解耦可能与地上和地下不同的选择压力有关。叶片特征受到最大化光合速率且最小化水分损失的驱动,因此叶片特征存在一维的权衡关系。与叶片不同,根性状受到了更为复杂的环境因子(土壤养分和物理条件)和生物因素(菌根侵染和其他根系微生物)的影响[71],导致了叶片和细根的形态特征的解耦。
同时,利用NSTEC地带性森林生态系统中植物叶片和细根的16种元素含量碳(C)、氮(N)、钾(K)、钙(Ca)、镁(Mg)、硫(S)、磷(P)、铝(Al)、锰(Mn)、铁(Fe)、钠(Na)、锌(Zn)、铜(Cu)、铅(Pb)、镍(Ni)、钴(Co),探讨了植物叶片和细根多元素协同变异规律。与细根相比,叶片中具有较高含量的大量元素(N,K,Ca,Mg,S和P,除C以外),而细根中具有较高含量的微量金属元素(Al,Fe,Na,Zn,Cu,Pb,Ni和 Co,除了Mn)[32]。进一步分析发现,植物谱系是影响植物叶片和根系元素含量变异的重要因素。植物分类(种科目水平的叠加)对叶片和细根元素含量变异的解释率均值分别为48.9%和42.6%。植物分类对叶片和细根中大量元素(C,N,K,Ca,Mg,S和P)的解释率分别达到63.7% 和 61.4%,而对微量元素(Al,Mn,Fe,Na,Zn,Cu,Pb,Ni和Co)的解释率分别为37.6%和28.0%。此外,植物分类对元素变异的解释率随着元素含量的升高而升高,并且在叶片和细根中存在相同的趋势。整体而言,与微量元素相比,大量元素的变异受到环境的影响较小。叶片N,K,Fe,Na和细根Pb,Ni变异较大地受到降水的影响,而叶片P和细根P,Al含量变化更多的受温度的影响。其中,叶片部分与之前对于叶片生物地理格局受到降水和温度影响的研究结果相类似[73, 74]。
由于叶片和地下根系的功能性状均存在较大的系统发育保守性,加之不同研究区域或物种选择的差异,导致根叶性状关系的相关研究结论存在较大争议。此外,叶片和细根性状之间的关联关系将在不同植物科或分支上存在差异,还有待进一步验证;此外,未来还应利用植物谱系结构更深入地探讨植物营养获取策略的进化格局及其驱动机制[31]。
4.4 植物C、N、P化学计量特征及其分配策略
生态化学计量学是研究生物多种化学元素平衡的科学,它通过元素比例在各个生命过程及生物地球化学循环中的数量关系,来认识和解决不同层次(基因,细胞,器官,有机体,群落,生态系统)的生态问题[75]。目前,已经从物种水平到区域尺度上对叶片C、N、P元素分配规律进行了大量研究[8, 74, 76]。但受测试难度和经费的限制,真正基于植物群落水平系统测试数据来开展相关研究的报道非常少,尤其是区域到全球尺度。在复杂的天然森林群落中,三个假设仍待系统的实测数据来证实;它们分别是:①元素分配假说(Element Allocation Hypothesis),即越活跃器官被分配更多的营养元素以实现功能最大化;②元素可塑性假说(Element Plasticity Hypothesis),即地下器官因其更复杂的环境因子和生物因素的影响而具有更高的元素可塑性;③ N-P异速分配假说 (N-P Allometry Hypothesis),即N-P异速分配指数在不同植物器官或不同功能群间存在一定的保守性。
利用NSTEC地带性森林群落中1100多种植物叶-枝-干-根的测定数据,发现叶片(植物器官中普遍认为是最活跃的)的N、P含量以及N∶P的值均高于枝(或茎)和根,一定程度上验证元素分配假说[77]。对不同功能群植物而言,N和P含量表现为草本高于木本,N∶P则表现为相反的结果,在一定程度上反映草本和木本不同的生存策略。如果用变异系数来表征元素可塑性,物种间N和P的变异系数均表现为根高于叶和枝,即根元素可塑性更高,与元素可塑性假说一致。对于N-P异速分配策略研究,尺度选择是一个非常重要的影响因素;在大尺度上,N-P异速分配斜率均小于1且在不同器官间或不同功能群间没有显著差异,证明了其较强的保守性(N-P异速分配假说);然而,在站点尺度,异速分配斜率有呈现大于1的情况[77]。基于长白山垂直样带的相关数据,叶片、茎、细根的N-P异速分配关系在不同生长型植物中存在差异[24]。在垂直梯度上,叶片、茎和细根的N、P分配对环境变化的响应一致,反映了植物各器官共存的基本机理。
在探讨植物元素化学计量学和分配策略时,还应在更大的范围对这三个假说进行验证,并探讨植物多元素的分配策略。此外,鉴于尺度选择对研究结果的巨大影响,未来在进行尺度扩展时需要考虑其合理性并选择适当的方法。
4.5 森林生态系统C、N、P化学计量特征及其纬度格局
现阶段,生态化学计量学的研究大多聚焦于物种或植物器官,对土壤和微生物也有一定研究;但由于天然森林群落结构的复杂性以及叶-枝-干-根的变异性,对植物群落C、N、P化学计量特征的研究十分缺乏。一方面造成生态化学计量学从分子到全球的联系难以完整,另一方面使相关生态模型因难以获得不同区域的相关数据而不能得到优化。此外,目前对群落化学计量特征的研究大多是以优势种为测定对象(或收集公开数据),再通过物种水平直接算数平均法推导,这种做法是否能真实反映复杂的天然森林群落还有待实验数据验证。
已有研究获取了NSTEC样带9个典型森林所有物种叶-枝-干-根、土壤等的C、N、P含量数据,采用群落结构和生物量加权法分析中国森林生态系统不同组分的C∶N∶P 化学计量特征[26]。该研究基于实测数据在区域尺度给出了生态系统不同组分以及植物不同器官的C∶N∶P化学计量特征(图5),并发现C∶P与N∶P有随纬度增加而减小的趋势。经过对气候(温度和降水)以及土壤养分(N、P含量)冗余分析发现,气候和土壤共同影响N∶P的变异,其中气候是主要影响因素。通过对N∶P比值分析,结合N∶P >16为磷限制的假设[78],发现中国森林在南方受到磷元素限制更为突出。此外,利用内稳性模型,计算了植物不同器官N∶P的内稳性指数,发现叶片的元素内稳性高于其他器官,说明越活跃的器官元素稳定性越高。需要指出的是:群落结构和生物量加权所获得的植物C∶N∶P化学计量特征,与物种水平直接算术平均值差异很大;此外,与国际上通过收集数据评估的结论相比,也存在很大差异[26]。
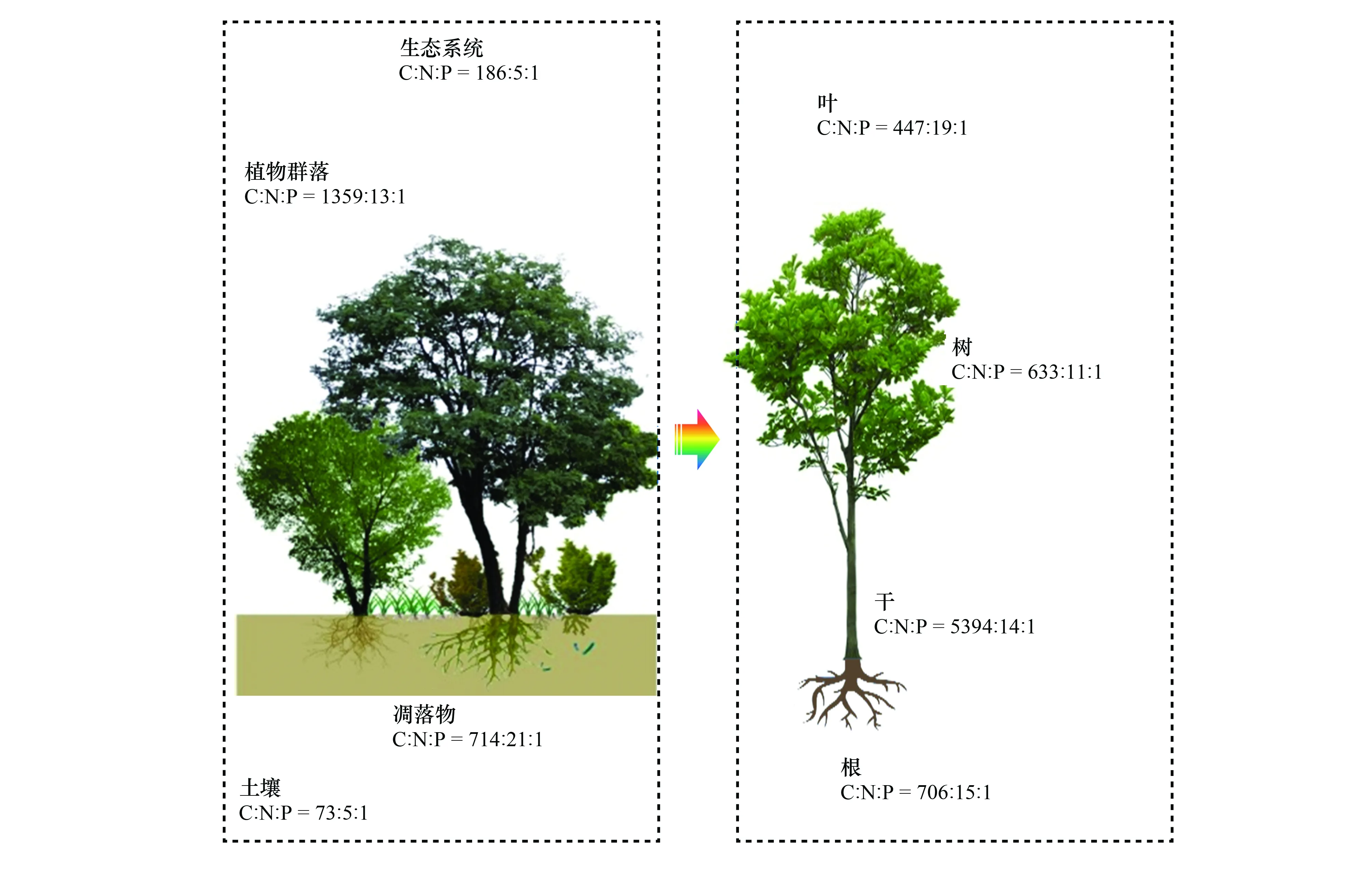
图5 中国森林生态系统各组分及植物各器官C∶N∶P化学计量特征Fig.5 Variation of C∶N∶P stoichiometry in different plant organs and different ecosystem components in Chinese forests
相关研究填补了生态系统及不同组分、植物不同器官C∶N∶P化学计量学特征的实测数据的空白,并定量揭示了从热带雨林至寒温带针叶林的纬度变异规律与影响因素。群落尺度的元素化学计量特征及其纬度变异规律,可为全球C、N、P耦合模型提供基础数据和理论支持。
5 土壤微生物和酶活性的纬度格局及其影响因素
5.1 土壤微生物群落的生物地理格局及其影响因子
传统上,土壤微生物的地理分布被认为是随机的,可在条件允许的任何环境中扩散和生长[79];同时,土壤微生物的空间分布特征与大型动植物存在明显区别[80, 81]。近年来,随着土壤微生物群落结构的大尺度空间分布格局研究的增多,越来越多结果表明土壤微生物具有地带性的分布规律,并将其归因于生态系统地上与地下联系、地理分隔和微生物的限制性扩散[82]。
采用磷脂脂肪酸法测定了NSTEC样带上9个地带性森林土壤微生物群落。实验结果表明:温带森林的土壤总磷脂脂肪酸(PLFA)要高于热带及亚热带。土壤细菌生物量的纬度格局与土壤总 PLFA的纬度格局一致。土壤真菌生物量无明显的纬度变异。热带及亚热带土壤真菌/细菌(F/B)、革兰氏阳性菌/革兰氏阴性菌(G+/G-)值要高于北方温带及暖温带森林土壤。NSTEC具有明显的水热组合梯度,其气候因子是土壤微生物生物量表现出纬度变异的重要原因。
在空间格局上,土壤微生物的群落结构可能受各种生物与非生物因素的影响[83]。NSTEC数据也表明温度、降水、土壤温度对土壤微生物群落结构变异的影响最大[84],在区域尺度,气候可以通过影响土壤温度及含水量来影响土壤微生物群落结构[85]。每种土壤微生物都有最适温度,当环境温度改变时,高于或低于其最适温度,微生物必定会被环境淘汰,土壤微生物群落结构进而发生改变。土壤水分增加将会导致某些细胞溶解,土壤含水量降低则导致土壤微生物胞质皱缩或质壁分离,从而改变土壤微生物群落结构。与其他环境因子相比,土壤酸碱度(pH)及植物功能群特征对NSTEC土壤微生物群落结构的纬度变异影响较小。植物功能群特征中,只有乔木比叶面积的群落权重值与土壤总PLFA、细菌、放线菌及G+呈显著或极显著负相关,而其他植物性状与土壤微生物生物量无显著相关关系。
5.2 土壤微生物功能多样性的生物地理格局及其影响因子
土壤微生物的代谢多样性是分析微生物对环境适应能力的有效指标[86]。有关环境因素及人类活动对土壤微生物的结构、多样性及酶活性的影响的研究已有很多[84, 87],并发现土壤微生物群落及酶活性具有生物地理分布格局。微生物的功能多样性还会随着植被类型及气候条件的变化而变化[88];不同气候区土壤微生物对碳源的利用能力不同,如北方森林土壤微生物更多的利用羧酸类碳源,而对氨基酸类的碳源的利用能力较弱[89]。
采用Biolog EcoPlate微平板培养法,测定了NSTEC样带的土壤微生物群落代谢活性。其中Biolog EcoPlate微孔板的总体颜色变化可用平均孔颜色变化率表示(Average well color development,AWCD),它可反映土壤微生物活性。实验结果表明:培养起始24 h内,AWCD变化不明显,而培养24—96 h间 AWCD快速增长,此时微生物活性旺盛,随后缓慢增长直至趋于稳定。土壤微生物群落的碳源代谢强度变化趋势与AWCD一致,呼中和东灵山森林土壤微生物群落的碳源代谢强度最低,分别为65.7和59.7。尖峰岭森林土壤最高为157.6;而其他7个站点土壤微生物群落碳源代谢强度无显著差异,变化范围为130—140。整体而言,在NSTEC样带土壤微生物碳源代谢强度无显著的纬度变异规律。
利用96 h的微生物吸光值对土壤微生物群落功能多样性指数进行了计算,结果显示不同的多样性指数均呈现相似的空间变化规律,从热带雨林到寒温带针叶林,土壤微生物群落功能多样性具有空间变异性。土壤微生物对碳源的利用能力随着纬度变化而变化,并可聚为4类,相同气候带森林土壤微生物的碳源代谢能力相似。通过对土壤微生物对6种碳源类型的利用程度的分析,发现土壤微生物对碳源的利用主要集中在糖类、羧酸、氨基酸和聚合物等4类物质,四者相对吸光值约为85%。土壤微生物对胺类及芳香类物质的利用程度较低,其相对吸光值之和约为15%。
在所有影响因素中,植物功能群特征是影响土壤微生物功能多样性纬度变异的主要因子。通常,含氮量高的植物生长速率更快,其凋落物也更有利于细菌生长[90],而含氮量低的植物凋落物则更利于真菌生长[84,90]。在NSTEC样带上,较高的叶片氮含量一定程度上限制土壤微生物的碳源利用能力[91]。
5.3 土壤细菌多样性的纬度格局及其影响因子
土壤微生物多样性的生物地理学研究,有助于深入挖掘土壤中的未知生物资源,深刻理解土壤中微生物多样性的产生、维持机制,并预测陆地生态系统结构和功能的演变方向[82, 92]。随着分子生物学的快速发展,越来越多研究表明土壤微生物群落组成和多样性在大尺度地理空间上呈现规律性分布[87, 93]。环境因素、历史进化因素(距离分隔、物理屏障、扩散限制和环境异质性等)或者二者共同作用是造成土壤微生物群落空间分布格局的主要原因[87, 93, 94]。
以NSTEC样带9个地带性森林土壤为研究对象,借助高通量测序手段,分析了不同森林土壤细菌α多样性(OTU 丰富度和Faith’s 谱系多样性)和群落结构的变化[95]。实验结果表明:亚热带和热带森林生态系统中细菌OTU 丰富度和Faith’s 谱系多样性显著低于温带森林生态系统,而土壤有机质的可利用性是引起细菌OTU 丰富度和Faith’s 谱系多样性变异的主要因素。此外,不同森林土壤细菌群落结构也存在显著变化。环境指标和地理距离可以解释80.5%的不同森林土壤微生物群落结构的空间变异;其中,气候因子、土壤和植物因素分别贡献了11.9%、14.2% 和7.8%。通过偏蒙特尔分析发现,温度、降水、土壤酸碱度和土壤有机质的可利用性是引起细菌群落结构变异的最主要的驱动因子。虽然植物叶片碳氮比会一定程度影响到细菌丰富度,但植物功能性状对土壤细菌多样性的贡献,远不如气候和土壤。
5.4 土壤酶活性的生物地理格局及其影响因子
土壤酶是土壤生物过程的主要调节者[96],与土壤有机质分解、矿质营养元素循环、能量转移等密切相关[97],在生态系统生物地球化学循环中具有重要作用。开展大尺度土壤酶活性的生态化学计量关系及影响因素的研究,能够为揭示区域尺度土壤碳循环的生物调控机制提供依据[98]。
利用微孔板荧光法,测定了NSTEC样带9个地带性森林土壤的酶活性,主要是与土壤碳循环相关的土壤β- 1,4-葡萄糖苷酶(BG)、与土壤氮循环相关的土壤β- 1,4-N-乙酰葡糖氨糖苷酶(NAG)和L-亮氨酸氨基肽(LAP)以及与土壤磷循环相关的酸性磷酸酶(AP)。实验结果表明:不同气候带的森林土壤BG、NAG、LAP、AP酶活性具有较大差异。在样带尺度,BG 和NAG 酶活性从寒温带森林到暖温带森林逐渐升高,随后又逐渐降低。总体上,北方森林土壤BG和NAG酶活性高于南方森林土壤。相反,土壤AP酶活性则表现为南方森林高于北方森林。根据热带雨林土壤较低的BG∶AP、NAG∶AP酶活性比值(0.21、0.13),可推断磷是该地区养分物质循环过程的主要限制元素。热带、亚热带森林土壤AP酶活性较高,而BG∶AP和(NAG+LAP)∶AP酶活性比值较低,表明亚热带及热带森林土壤磷有效性较低,生态系统更受磷元素的限制。对全球土壤AP酶活性的整合分析发现,随着纬度降低、气候逐渐趋于湿热,土壤AP酶活性逐渐升高,而BG∶AP、(NAG+LAP)∶AP酶活性比值逐渐减小[99]。
森林土壤BG、NAG酶活性的空间分布与年均降水量格局呈显著负相关,并随着年平均温度的升高而显著降低,但没有发现土壤LAP和AP酶活性空间分布与年均降水量、年均温显著相关,年均温可能是影响全球尺度土壤酶活性的一个关键因子。年均降水量通过影响土壤含水量来影响底物的扩散速率[100],从而影响土壤酶活性。在NSTEC样带上,土壤BG、NAG酶活性与土壤酸碱度呈显著正相关,而土壤AP酶活性与土壤酸碱度呈负相关[90, 101]。
6 土壤有机质的纬度格局与影响因素
6.1 土壤有机质组分的纬度格局与影响因素
土壤有机质十分复杂,根据其周转时间可分为活性有机碳和稳定有机碳[102]。土壤活性有机碳易被土壤微生物分解利用,且易氧化分解并对环境和人为干扰的响应更敏感[103, 104];土壤活性碳组分空间格局及其影响因素的分析,有助于阐明土壤有机碳(SOC)在人为干扰或气候变化情景下的早期变化和轨迹[105]。
以NSTEC样带9个典型森林土壤为研究对象,分析测试了其表层SOC、易氧化有机碳(EOC)、土壤微生物碳(MBC)和可溶性有机碳(DOC)含量[106]。实验结果表明:SOC含量随纬度增加呈增加趋势,在凉水最大,尖峰岭最小。土壤EOC含量随纬度增加也呈增长趋势,9个森林站点EOC含量差异显著并在凉水最大。MBC随纬度增加整体呈增加趋势,在呼中最大而东灵山最小;不同地区间差异很大,最大值与最小值间可达7.85倍。DOC含量的纬度变化格局与其他碳组分完全相反,其含量随纬度增加呈降低趋势。从气候区来看,SOC和EOC整体表现为热带森林<亚热带森林<温带森林;DOC则表现为热带森林>亚热带森林>温带森林。
基于结构方程的分析,气候、植被生物量、土壤质地和土壤微生物分别解释了SOC、EOC、MBC和DOC纬度变异的74%、65%、51%和76%。其中,SOC和EOC纬度格局主要受气候和土壤质地的影响,植被生物量和土壤微生物的影响相对较弱。MBC纬度格局的主要控制因素是气候,土壤质地对其影响十分微弱。对于DOC纬度格局,气候和植被生物量表现为正效应,土壤质地则表现为负效应。
6.2 土壤腐殖碳组分纬度格局及其影响因素
土壤腐殖质是土壤稳定性碳库的主要来源,长期而言土壤腐殖质的损耗将造成大量二氧化碳从土壤向大气输入[107];同时,土壤腐殖质的化学抵制力是控制土壤和大气间碳循环的重要过程[108]。因此,大尺度的土壤腐殖碳组分及其格局形成机制,将为碳动力模型的发展及验证提供数据支撑。
以NSTEC样带9个地带性森林土壤为研究对象,分析测试了其表层土壤可提取腐殖质碳(HEC)、胡敏酸碳(HAC)、富里酸碳(FAC)和胡敏素碳(HUC)含量。实验结果表明:森林土壤腐殖碳组分HAC、FAC、HUC和HEC含量范围分别为:0.44—1.62 g kg-1、7.80 —18.30 g kg-1、11.37—49.98 g kg-1和8.53—19.94 g kg-1;它们在9个森林类型间都存在显著性差异。整体而言,森林土壤腐殖碳组分HAC、FAC、HUC和HEC均随纬度的增加呈增长趋势。按气候带类型划分,森林土壤腐殖质碳组分都表现为热带<亚热带<温带的特点,HAC和HEC在热带、亚热带和温带之间都差异显著[109]。
气候、土壤质地和土壤微生物对森林土壤腐殖质碳组分HAC、FAC、HUC和HEC的解释率分别为91.32%、90.94%、94.22%和94.13%。气候因素对HAC、FAC和HEC的影响最大,分别为78.72%、88.61%和89.8%;土壤质地对HUC空间格局的影响最大,可解释其85.92%空间变异。
6.3 土壤和植被碳贮量空间分布格局
科学家围绕森林碳贮量的空间格局及其控制因素开展了大量的研究,但绝大多数区域尺度的研究都是单独探讨植被碳贮量和土壤碳贮量的空间格局及其影响因素[110- 113],而同时考虑森林植被和土壤碳贮量的空间格局的研究却鲜有报道,限制了人们对整个森林生态系统碳贮量的空间分布格局和控制机制的认识。
以中国东部森林为研究对象,收集和整理了2004—2014年间中国东部森林公开发表科研论文中的植被(地上生物量+根生物量)和土壤碳贮量数据,共包括1471个样地的植被数据和1999个样地的土壤数据。在随机取样下,植被和土壤碳贮量存在纬度格局;如果仅考虑成熟林,则植被和土壤碳贮量表现出更强的纬度格局。此外,随着统计尺度的增大(纬度2°和5°),森林植被和土壤碳贮量的纬度分布格局越发明显[25]。在2°纬度下统计,成熟林植被碳贮量从南向北由196.31 Mg C/hm2降低到51.71 Mg C/hm2,并随纬度增加呈显著的负指数关系;然而,0—10 cm土壤碳贮量随着纬度的增加却呈指数增加,由南向北土壤平均碳贮量从35.29 Mg C/hm2增加到123.28 Mg C/hm2;出人意料,森林生态系统(植被+土壤)的碳贮量没有明显纬度格局。植被和土壤碳贮量的分配比例(植被:土壤)随纬度增加呈显著的负对数关系,该比例系数能一定程度地为森林碳密度估算模型的建立提供科学指导。
气候因素不仅决定着森林植被和土壤碳贮量的纬度分布格局,而且还控制着森林植被和土壤碳贮量分配比例的变化规律;然而,气候对植被碳贮量和土壤碳贮量的控制机制完全相反,对中国东部植被碳贮量而言降雨量更重要,而对土壤碳贮量而言温度更重要[25]。随着纬度的增加,地表凋落物呈逐渐增加的变化趋势[114]。利用多源数据(收集文献数据和中国科学院碳专项实测数据),何念鹏等还发展了基于经典Logstic 生长方程的中国森林植被碳固持模型(Forest Carbon Sequestration Model, FCS model),并成功地对2010—2050植被固碳潜力及其空间格局进行了预测[115]。相关研究结论揭示了全球气候变化下森林的固碳效应,同时建议在理论研究或模型优化时应将植被和土壤分开考虑。
7 土壤有机质周转速率及其温度敏感性
7.1 土壤碳矿化温度敏感性的空间变异及其影响因素
土壤有机质分解所释放的二氧化碳是陆地生态系统与大气间最大的气体交换通量之一[116],土壤有机质分解对温度变化非常敏感;因此准确地评估温度变化对土壤有机质分解的影响,对预测土壤碳库变化及其对全球气候变化的反馈具有重要意义。温度敏感性(Q10)被用来表示土壤有机质分解对温度变化的响应,它为不同生态系统类型或研究间提供了一个规范的和可比较的参数,同时也是许多碳循环模型的重要参数[117]。然而,由于各个独立研究中传统培养模式与测试方法的差异(且集成在单点或单个区域),导致不同研究结果间可比性较差,使许多科学家通常在模型中将Q10设定为常数(2.0或1.5),是模型预测不确定性的重要来源[118, 119]。如何革新土壤有机质分解温度敏感性的研究方法,对于降低不同区域研究间的不确定性、准确评估土壤呼吸对气候变化的响应及其反馈均具有重要意义。
2013年以来,何念鹏等自主研发了土壤有机质温度敏感性的测定系统[120, 121],发展了“自动变温培养+连续自动测试”新模式。新模式除了较好地克服了传统模式(恒温培养+间断测试)存在的主要问题(微生物适应性、底物消耗不均、操作过程繁琐、样品测试量大),还被成功应用于研究不同生态系统类型或情景下土壤有机质温度敏感性[41, 122- 125]。基于NSTEC样带的研究,发现土壤有机质基础分解速率(20 ℃)在不同森林类型间差异显著,且随着纬度的升高而显著升高。而且土壤有机质分解的温度敏感性(Q10)在不同森林类型间差异显著,随着纬度的升高显著升高,为低温区更敏感假设提供了直接的证据[41]。在此基础上,对中国东部森林土壤以及内蒙和西藏草地土壤温度敏感性进行了测定,发现Q10变化范围从1.16到3.19(均值1.63),且在不同生态系统间差异显著:高寒草地(2.01)> 温带草地(1.81)> 热带森林(1.59)> 温带森林(1.55)> 亚热带森林(1.52)[42]。
在未来气候变暖的背景下,需要特别关注高纬度森林和高寒草地生态系统。在不同的生态系统中,Q10与底物质量之间显著的负相关关系均支持碳-质量温度假说,表明该假说也适用于较大空间尺度的陆地生态系统。环境因子和土壤因子共解释了52%的Q10空间变异;其中,土壤pH值和电导率是最主要因素。在中国东部森林,这两个因素又主要受到气候因素所影响,因此,未来应当考虑将这两个因素加入模型,更准确的模拟及预测有机质分解对气候变化的响应与反馈。
7.2 土壤氮矿化速率及其温度敏感性的空间格局与影响因素
7.2.1 土壤总氮矿化速率的空间格局与影响因素
氮素是植物生长的重要营养元素,土壤氮素有效性一定程度上决定了陆地生态系统的净初级生产力大小[126]。土壤总铵化速率和总硝化速率是土壤供氮能力的重要指标,硝化作用指由微生物将有机氮或铵态氮转化为硝态氮的过程,是植物直接吸收利用的仅次于铵态氮的另一种无机氮形式[40, 127]。 尽管一些研究在单站点尺度上分析了土壤的铵化或者硝化作用,但是在较大的区域尺度上,森林土壤总铵化和总硝化作用对于不同的环境因素变化的响应及其机制仍不清楚。
采用15N库稀释法进行野外原位标记和室内标记的培养实验,探讨了从热带到寒温带9种配对森林土壤(原生林vs.次生林)的潜在铵化和硝化速率[40]。实验结果表明:野外平均铵化速率是(4.9 ± 0.5) mg N kg-1d-1,而平均潜在铵化速率是(32.0 ± 8.6) mg N kg-1d-1。野外平均硝化速率为(1.7 ± 0.3) mg N kg-1d-1,潜在硝化速率为(3.2 ± 0.6) mg N kg-1d-1。 在样带尺度上,铵化速率显著高于硝化速率,并且在不同森林中铵化速率和硝化速率具有较大的变异性。铵化速率和硝化速率与气象因素(温度和降水)之间存在显著的相关关系。结构方程模型表明:土壤氮含量,微生物氮库以及细菌的丰度是影响中国东部森林土壤铵化和硝化速率的主要因素。土壤因素对铵化和硝化速率较强的控制,意味着只有准确地定量土壤因素及其对土壤氮转化过程影响,才能更好地改进土壤氮模型。
7.2.2 土壤净氮矿化及其温度敏感性的空间格局及其影响因素
在全球变暖的背景下,土壤净氮矿化速率对温度变化响应的微小变化,都会改变土壤的氮素有效性,并一定程度上影响陆地生态系统的初级生产力[128]。净氮矿化作用对温度变化的响应也可用温度敏感性(Q10)表示,这一参数被广泛用在土壤N循环模型中[129]。过去关于土壤净氮矿化温度敏感性的研究主要集中在单点上,而在大尺度上土壤净氮矿化及其温度敏感性的研究极少[130]。
基于1990—2014年间公开发表论文的土壤净氮矿化及其温度数据,探讨了净氮矿化及其温度敏感性的空间格局及调控机制[131]。结果显示:在中国陆地生态系统中,土壤净氮矿化速率的均值为(2.78 ± 0.18) mg N kg-1d-1,它随着纬度的升高显著降低。另外,土壤净氮矿化速率随着海拔高度的增加呈显著的对数降低趋势。在不同森林植被类型中,常绿针叶林和常绿阔叶林土壤净氮矿化速率高于落叶针叶林和落叶阔叶林。土壤净氮矿化的Q10均值为1.58,并在不同的生态系统间差异显著。在森林生态系统中,常绿针叶林的Q10显著高于落叶阔叶林和常绿阔叶林。土壤净氮矿化的Q10随着纬度和海拔的增加均呈显著增加的趋势,支持越寒冷的区域其温度敏感性越高的理论假设。通径分析结果显示:土壤净氮矿化速率主要受到年均温和土壤有机质含量的影响,而Q10主要受到土壤总氮含量的影响。此外,关于全球土壤氮矿化的整合研究也表明,土壤净氮矿化的Q10存在明显的纬度格局且在生态系统类型间差异显著[130]。这些结果表明在气候变暖的背景下,寒冷区域土壤的氮素有效性可能会增加,进而促进生态系统初级生产力的增加。
8 挑战与展望
通过对近年来NSTEC典型森林样带性状研究的简要总结,可以清楚地看出在如下几个方面取得较好的进展:①生态系统性状(植物、微生物、土壤)的系统化测试,构建了一个系统的、独特的性状数据库,并探讨基于典型森林生态系统性状研究的新调查模式,为更好地基于性状数据探讨生态系统结构与功能及其对环境适应奠定了基础。该模式的提出和成功的应用,将会一定程度上引领未来性状研究的调查方式,并促进系统性调查数据的快速积累。②基于所测定的大量数据,不仅按传统途径探讨了植物、微生物和土壤性状的纬度变异规律与影响因素,其中如气孔性状、解剖结构性状、叶绿素含量、非结构性碳水化合物含量、土壤碳矿化、土壤净氮矿化、温度敏感性、生态系统各组分C∶N∶P化学计量特征等的大尺度分析,填补了相关研究的空白。更为重要的是以系统性的调查数据为基础,发展了基于植物群落结构和生物量(叶片面积)加权法的从器官至群落的拓展方法,实现了单位土地面积(或单位器官)基础的群落性状推导,较好地解决了国内外性状研究所面临的巨大挑战和难题。③定量揭示了天然森林生态系统中(部分)性状与初级生产力、净初级生产力、水分利用效率等的定量关系,为今后进一步探讨天然森林群落性状与功能的关系提供了新的方法学依据和范例。
由于新调查模式所获得的系统性数据,更接近自然群落或生态系统,因此,除了现有的分析角度或思路,还应该具有更多的潜在用途。首先,从传统生态学角度可以深入探讨如下几方面的科学问题:①综合分析叶片或根系性状的相互关系,甚至考虑分级化的“叶片性状网络”“根系性状网络”,再从点-样带-区域多层次探讨性状网络变异及其机制。如上所述,简单地从几个指标探讨的关系有其科学价值和进步意义,但还难以真实地揭示自然群落中植物性状间的关系,其研究类似于“瞎子摸象”,难以揭示其整体关系与影响机制;类似的思路还适用于土壤微生物,是否可以建立 “微生物性状网络”。②利用系统化的调查数据(植物群落结构-土壤微生物群落结构-土壤理化性质-气候因子、群落内每个植物物种叶-枝-干-根性状、叶片形态-气孔-解剖结构-化学元素-叶绿素含量-非结构性碳水化合物等),深入探讨森林生态系统群落结构在点-样带-区域尺度的维持机制,探讨经典生态学的核心科学问题“进化vs.适应、环境过滤效应vs.竞争效应、生态位分化理论vs.中性理论”等,推动生态学理论研究的发展;③ 结合点-样带-区域尺度的生态系统生产力参数,从叶片性状协同优化(或权衡)角度,深入探讨森林生态系统生产力优化机制。
同时,能否为传统性状研究切实服务于宏观生态学或宏观地理学提供技术探讨的平台?如果能实现该目标,既可极大拓展性状研究的范畴,又能使传统性发展数据更好地服务于当前区域甚至全球生态环境问题。然而,传统性状数据与遥感、模型和能量数据间存在巨大的裂隙,使它们之间很难相互验证或支持;具体表现为:①传统性状数据大多是在个体或群落类少数物种测试,缺乏相配套的群落结构数据,使其数据很难推导到群落尺度;然而,宏观生态学的主要观测尺度都是群落尺度;②传统性状数据的测试指标的量纲更多是根据自身要求,集中在器官水平的含量或密度,而宏观生态学的量纲大多在单位面积或体积,因此二者间如何转化也是一个非常棘手的难题;③传统的性状与功能的关系,大多是在器官或物种水平建立的(如叶片氮含量与光合速率的关系、叶片叶绿素含量与光合速率的关系等),其是否适用于复杂的群落还有待商榷。上述三个难点是相互关联的,只有通过系统性的性状调查数据才能逐一解决。此外,为了更好地构建性状研究(植物、动物、微生物等)与宏观生态学研究的桥梁,需要从更高层次提出新的概念,如生态系统性状(Ecosystem trait)。在此,将生态系统性状定义为以单位土地面积为基础,在群落尺度能体现生物(植物、动物、微生物)对环境适应性和生产力优化的性状;即生态系统性状是由植物群落性状、动物群落性状、土壤微生物群落性状等组成。它可以在相对统一的空间尺度和量纲上将植物群落性状、动物群落性状、土壤微生物群落性状、土壤属性和气候要素等紧密联系起来,为深入研究不同尺度生物-非生物性状间的关系奠定坚实基础,并以群落性状为核心连接宏观生态学的主要研究途径(遥感观测、通量观测和模型模拟),推动传统性状指标与宏观生态学的应用相结合。
