克隆植物蛇莓相连分株对异质性 盐胁迫的可塑性响应
2018-10-29陆陈丹阳钱永强孙振元
陆陈丹阳,钱永强,2,王 越,孙振元,2
(1.中国林业科学研究院林业研究所 国家林业局林木培育重点实验室,北京 100091; 2.林木遗传育种国家重点实验室 中国林业科学研究院,北京 100091)
盐分胁迫是世界上危害植物生长较严重的非生物胁迫因素之一[1],其限制了植物的生长和作物产量[2]。目前,全球盐渍化土地约9.55×108hm2,占全球陆地面积的10%[3],其中次生盐渍化土地约7.7×107hm2[4],因盐渍化而荒废的土地在每年以1 000万hm2的趋势增长[5]。由于土地的不合理开发利用和生态环境的恶化,全球土壤盐渍化面积仍在迅速增加[6-7]。在盐渍化地区,植物长势缓慢,种类单一,资源匮乏,严重影响了农业生产、生态环境与可持续发展[8]。目前,植物对盐胁迫的适应性成为了土壤盐渍化领域的研究热点[9]。
克隆植物是一个在自然界中广泛存在的植物类群,其特点是能够通过与基株相连的根、芽、蘖、枝及匍匐茎等的营养生长,形成许多互相联系的无性系分株[10-11]。在自然环境中,植物生长所需的资源(如光照、水分和养分等)在时空上分布极不均匀(资源异质性),克隆植物可以通过相连分株之间物质和能量的交流与共享,实现克隆分株间的生理整合(physiological integration),有效地缓解资源异质性对生长和繁殖带来的不利影响,从而使克隆植物能够适应异质性恶劣环境,在生态系统更新、恢复过程中扮演重要的角色[12]。
蛇莓(Duchesneaindica)原产于中国,是一种典型的克隆植物,为蔷薇科(Rosaceae)蛇莓属(Duchesnea)的多年生草本植物[13]。该植物能够产生较长的地上匍匐茎从而表现出克隆生长习性,匍匐茎在横向生长一段时间后,其顶芽转向上形成节间极度缩短的直立茎,直立茎基部产生许多不定根,从而形成分株[14]。克隆分株间的生理整合使其在资源贫瘠的异质环境下表现出极强的空间拓展和生存竞争能力[15]。异质性生境下野生草莓相连分株间资源交互传输,这种资源共享缓解了局部的资源缺乏,从而增强了远、近端克隆分株部分和克隆片段的生长[16]。近几年对蛇莓的研究主要集中在其化学活性成分和药理,以及小尺度克隆结构和克隆构型对不同环境(如海拔、土壤养分、水分)的可塑性响应等方面[17-19]。但是关于蛇莓等克隆植物克隆生理整合的调控机理研究较少。本研究拟通过不同盐浓度处理蛇莓姊妹株在连接与断开状态下生长与生理变化规律,揭示其生理整合特性对盐胁迫的响应机制,为克隆植物在逆境生理领域提供理论依据,也为蛇莓在贫瘠土地绿化提供理论指导。
1 材料与方法
1.1 材料培养
采用野生蛇莓作为试验材料。通过多世代无性繁殖,获得大量基因型完全一致的蛇莓分株,将单株分株作为母株栽至15 cm×15 cm花盆中,栽培基质为草炭土∶珍珠岩∶蛭石=7∶2∶1,培养3周后母株开始形成大量匍匐茎,将匍匐茎固定于花盆周围预铺的珍珠岩基质上,确保匍匐茎节能及时形成根系,继续培养2~3周后,挑选节间长度适中、长势良好、根系发育状态一致的蛇莓相连分株,在连接状态下将其栽至10 cm×10 cm的花盆中,用作试验材料。温室内气温为26 ℃左右,相对湿度为70%~75%。
1.2 材料处理
选取长势一致的成对蛇莓植株80对,相连分株与断开分株各40对,并标记好姊株(X)、妹株(Y),断开分株组为对照组。配制0(对照)、100、200、300mmol·L-14个浓度水平NaCl溶液进行胁迫,每3 d对蛇莓姊株施加100 mL的NaCl溶液,施加前先用去离子水冲洗基质,以减少前摄盐分基质对NaCl浓度的影响,相连妹株施加等量的水(图1)。培养第14 天时,取样测定有关指标,每个处理10对,生物量与过氧化氢含量部位的测定每个处理3次重复,叶片相对含水量和细胞膜透性的测定每个处理重复8次。

图1 处理示意图Fig. 1 Porcessing diagram
1.3 整株生物量的测定
选取完整植株洗净后,剪除匍匐茎,置于105 ℃烘箱中杀青20 min后,70 ℃下烘干48 h至恒重,称其总生物量。
1.4 植物相对含水量的测定
取植物叶片用去离子水迅速冲洗干净,用吸水纸擦干其表面水分后称其鲜重(Wf)。将叶片完全浸泡于蒸馏水24 h至恒重,称其饱和鲜重(Wt)。将新鲜材料置于105 ℃烘箱中杀青20 min后,70 ℃下烘干48 h至恒重,称其干重(Wd)。
相对含水量RWC=(Wf-Wd)/(Wt-Wd)×100%。
1.5 植物细胞膜透性的测定
取植物叶片用去离子水迅速冲洗干净,用吸水纸擦干其表面水分后,使用直径为5 mm的打孔器避开叶脉打孔,置于10 mL去离子水中完全浸泡12 h,使用DDS-12DW电导率仪测得电导率EC1。再置于沸水浴加热20 min,使质膜完全被破坏,待冷却至室温后,测得电导率EC2。细胞膜透性=EC1/EC2×100%。
1.6 植物过氧化氢含量的测定
配制3,3’-二氨基联苯胺(DAB)染色液,使用NaOH将pH调至5.8,浸泡蛇莓叶片,于28 ℃下避光保存8 h,光照8 h进行充分染色。倒净并吸取染色液后加入80%乙醇溶液,沸水浴加热至叶片完全脱色。静置至室温后取出叶片拍照记录,将照片使用Image J处理,计算叶片染色部位与叶片总面积之比(图2)。
1.7 统计分析
采用Valladares等[20]的方法计算各指标的可塑性指数,即4个盐浓度梯度下,某变量的最大值与最小值之差除以最大值。使用Microsoft Office Excel 2007软件进行数据的整理和作图,SPSS 18.0统计分析软件进行单因素方差分析(ANOVA),并用Duncan’s法检验其差异显著性。

图2 不同NaCl浓度处理对蛇莓匍匐茎相连与断开状态下的叶片过氧化氢氧化面积比例Fig. 2 Effect of different salt concentration on H2O2 proportion of the proximal ramets with connecting or severing stolon
2 结果与分析
2.1 在相连或断开状态下盐胁迫对蛇莓姊株的影响
在300 mmol·L-1NaCl胁迫下,匍匐茎连接状态下蛇莓姊株的生物量显著高于匍匐茎断开蛇莓姊株的(P<0.05)。其他浓度0、100、200 mmol·L-1NaCl处理的则相反,断开姊株高于连接的姊株,但差异不显著(P>0.05)(图3);与断开姊株相比,100和200 mmol·L-1NaCl胁迫下,相连蛇莓叶片相对含水量分别上升了13.35%、18.30%,二者差异显著(P<0.05);第14天时,300 mmol·L-1的NaCl胁迫下的蛇莓单株出现死亡;100、200 mmol·L-1浓度NaCl胁迫下,蛇莓姊株在断开状态下叶片细胞膜透性显著高于匍匐茎连接状态下叶片细胞膜透性(P<0.05),不同状态下蛇莓断开姊株叶片细胞膜透性随NaCl浓度的升高而显著增大(P<0.05),100和200 mmol·L-1NaCl胁迫下断开姊株叶片细胞膜透性较匍匐茎连接状态分别上升了18.91%、59.83%;在100 mmol·L-1NaCl胁迫下的蛇莓姊株在不同状态下H2O2含量的差异不显著(P>0.05);200 mmol·L-1浓度的NaCl处理下,蛇莓姊株在断开姊株叶片H2O2含量明显大于匍匐茎连接姊株叶片(P<0.05)。
比较4种盐浓度处理蛇莓姊(X)株在匍匐茎连接或断开状态时生物量、相对含水量、细胞膜透性和过氧化氢含量的可塑性指数可看出,过氧化氢含量的可塑性指数最高(0.76),生物量与细胞膜透性次之,分别为0.6和0.58,相对含水量最小(0.2),整体来看蛇莓受到盐胁迫时生理可塑性(细胞膜透性与过氧化氢含量)>生长可塑性(生物量和相对含水量)。在该4项指标中,蛇莓姊株在匍匐茎断开状态下的可塑性指数均大于其在相连时的可塑性指数。
2.2 盐胁迫对蛇莓相连姊妹株的影响
当相连姊株受不同浓度NaCl处理时,其相连妹株间的生物量、相对含水量与细胞膜透性与对照间无显著变化(P>0.05);而随着姊株受到NaCl处理浓度的升高,与其相连的妹株叶片的H2O2含量较对照也随之大幅度升高,100、200和300 mmol·L-1NaCl胁迫下叶片的H2O2含量较对照显著上升(P<0.05),分别上升了46.16%、57.15%、56.25%(图4)。

图3 不同NaCl浓度处理对匍匐茎相连与断开状态下蛇莓姊(X)株生物量、相对含水量、细胞膜透性和H2O2含量的影响Fig. 3 Effect of different salt concentrations on total biomass, relative water content (RWC), membrane permeability, and H2O2 proportion of the proximal ramets (X) with connecting or severing stolon
不同小写字母表示不同盐浓度处理间的差异显著(P<0.05);不同大写字母表示相连与断开状态间差异显著(P<0.05)。图4同。
Different lowercase letters indicate significant difference among different NaCl concentration treatments at the 0.05 level and different capital letters indicate significant difference between connected and severed stolon at the 0.05 level; similarly for the Figure 4.
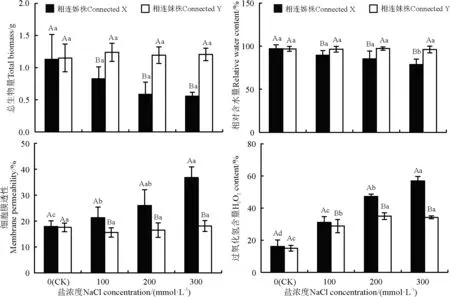
图4 不同NaCl浓度处理成对蛇莓相连姊(X)妹(Y)株生物量、相对含水量、细胞膜透性和H2O2含量的影响Fig.4 Effect of different salt concentrations on total biomass, relative water content (RWC), membrane permeability, and H2O2 proportion of proximal ramets (X) and distal ramets (Y) with connecting stolons
100、200、300 mmol·L-1的NaCl胁迫下的蛇莓相连分株中,姊株与妹株之间的生物量出现了差异,姊株生物量显著低于妹株生物量(P<0.05),姊株比妹株的生物量分别下降了33.15%、51.12%、53.74%(图4)。100、200、300 mmol·L-1的NaCl胁迫下,姊株与妹株之间的相对含水量、细胞膜透性、H2O2含量均出现了显著差异(P<0.05),在100、200和300 mmol·L-1NaCl胁迫下,姊株叶片的相对含水量比妹株的下降了7.34%、12.26%、17.97%;100、200、300 mmol·L-1的NaCl胁迫下,姊株叶片的细胞膜透性和过氧化氢含量比妹株显著上升(P<0.05),分别上升了36.96%、57.56%、103.44%和7.85%、35.36%、66.17%。
3 讨论与结论
环境异质性在自然环境中普遍存在,这使得植物获取必需资源的难度大大增加[21]。相对于非克隆植物,生理整合特性增强了克隆植物对异质性环境的适应能力[22],如克隆植物空心莲子草(Alternantheraphiloxeroides)的克隆整合特性使其对酸雨胁迫有着较好的适应性[23]。生物量是植物耐盐性最直接的指标,也是植物对盐分胁迫的综合表现[24-25]。异质性盐胁迫下,蛇莓断开姊株生物量显著下降,而在各浓度处理下的相连姊株生物量则未呈现明显差异,说明在蛇莓相连分株中,未处理的相连妹株会通过生理整合作用对受到NaCl处理的姊株起到缓解作用,克隆植物沙鞭(Pasammochloavillosa)在风蚀条件下也有着类似的表现[26]。而随着盐浓度的增加,蛇莓姊株受到NaCl胁迫加重时,相连妹株的生物量、叶片相对含水量与细胞膜透性并未随之受到影响。这表明,妹株的生长并不因为姊株受到胁迫而受到明显抑制。因此,在异质性胁迫下未受胁迫或胁迫较轻的分株能够保持较为正常的生长。
克隆植物的相连分株通过横生结构(间隔子)互相连接,为分株间的物质传输和交换提供了可能[27]。蛇莓姊株相连状态下叶片的相对含水量与断开状态下相比,随着浓度的升高,差异显著,证明了蛇莓相连分株中的克隆整合能够缓解姊株受到的胁迫作用。Alpert等[28]、罗栋等[29]和Zhang等[30]在野牛草(Buchloedactyloides)、草莓(Fragariachiloensis)、空心莲子草等克隆植物中发现,处于高养分的分株可给低养分的分株提供显著的支持,实现资源共享。而当受害程度积累后,受胁迫相连姊株的叶片细胞膜透性、过氧化氢含量与未受胁迫姊株比出现显著差异,且浓度越高差异越显著,说明这种缓解作用具有一定的限度。甚至,在入侵性克隆植物空心莲子草具有较强的匍匐茎断裂机制,当受胁迫株受害严重时,近株端的匍匐茎干枯,具有主动“切断”生理整合的可能[31]。有的克隆植物长期于逆境环境时,会减少或中断克隆分株间的资源共享,从而利用更多的资源抵御胁迫环境[32]。生长于干旱沙漠的赖草(Leymussecalinus)在产生新分株时则较少依赖相邻分株供给的资源,以利于减少资源运输时的消耗[33]。
细胞膜是细胞与外界环境进行物质交换的主要通道,对维持细胞正常代谢起着重要作用[34]。在逆境条件下,胁迫因子积累到一定时间或强度时,细胞膜直接受到逆境的伤害,当其受到损伤后,细胞内电解质扩散出细胞,通过测定细胞膜透性的变化,反映逆境胁迫对细胞膜的伤害程度[35]。本研究表明,蛇莓姊株在与妹株相连或断开状态下,受胁迫姊株的膜系统均受到一定程度的损伤,并随着胁迫时间的延长、浓度的升高而加剧。姊株在匍匐茎断开状态下的细胞膜透性明显高于其在连接状态下的细胞膜透性,与此同时相连分株的妹株与空白对照相比,其细胞膜透性也呈现出差异。
研究过程发现,在300 mmol·L-1的NaCl溶液处理14 d时,受胁迫蛇莓姊株在单株状态下时已死亡,而在与妹株相连时还依然存活,且各项指标中前者的可塑性指数均高于后者,说明单株状态的姊株受胁迫程度较重,连株状态的未受胁迫妹株通过匍匐茎对相连姊株受到的胁迫起到缓解作用。与此同时,在匍匐茎相连状态的处理组中,未受胁迫的妹株受到姊株的牵连并不显著,继而妹株进行较为正常的生长。由此表明,在高浓度盐胁迫下,克隆植物相连分株的适应性明显优于非克隆植物或克隆植物单株,这与前人研究在异质环境中克隆植物结缕草(Zoysiajaponica)克隆分株在连接或断开下的影响[36]以及克隆植物白三叶(Trifoliumrepens)、蛇莓、活血丹(Glechomalongituba)对异质性UV-B辐射的响应有着相似的发现[37]。
盐胁迫作为影响植物正常生长与生理代谢的主要逆境因子[38-39],会诱导植物细胞内产生活性氧,发生氧化反应,将生物大分子降解成各种小分子物质并生成具强氧化性的过氧化物,导致膜质过氧化[40]。3,3’-二氨基联苯胺(DAB)染色技术是对植物内源H2O2原位定位的染色方法,DAB可与H2O2在产生处反应,形成褐色斑点,因此利用DAB染色可以对叶片进行组织化学原位测定[41],并通过图片处理即可计算过氧化氢的聚集情况。本研究结果表明,在中、高浓度NaCl处理时,匍匐茎断开时的姊株叶片积累的H2O2明显高于相连分株姊株叶片;在各浓度胁迫中,未受处理的相连分株中的妹株与空白对照比也呈现明显差异;姊、妹株的差异随处理时间的延长而逐渐显现。这表明,在蛇莓相连分株中,匍匐茎具有传递信号的功能,并通过生理整合的作用缓解受胁迫植株的伤害。然而具体的信号分子及其响应机制尚不明朗,还需做进一步探究。
盐胁迫抑制了蛇莓植株的生长,使得细胞膜透性、过氧化氢氧化活性升高;克隆植物蛇莓分株在连接状态下,一定程度上缓解了盐胁迫对蛇莓姊株生长造成的氧化伤害,维持了膜系统的稳定性与完整性,从而增强了蛇莓姊株的耐盐性。虽然本研究从蛇莓植株生长与生理角度揭示了克隆植物蛇莓分株在相连与断开状态下对盐胁迫的响应,但对异质环境胁迫感知的信号分子及传递形式仍不明确。因此,结合形态反映、生理代谢调节,进一步分离和鉴定逆境响应相关信号分子并明确其调控网络,将有助于更全面解析蛇莓对异质逆境的响应及适应性调控机制。
