广东水稻细菌性条斑病菌致病性分化研究
2018-10-29冯爱卿汪聪颖汪文娟陈尉芹杨健源朱小源
冯爱卿,汪聪颖,汪文娟,陈尉芹,苏 菁,杨健源,陈 深,朱小源
(广东省农业科学院植物保护研究所/广东省植物保护新技术重点实验室,广东 广州 510640)
由细菌性条斑病菌(Xanthomonas oryzaepv.oryzicola,简称Xoc)稻黄单胞菌侵染引起的水稻细菌性条斑病,于1957年首次发现于广东珠江三角洲,是国内植物检疫性水稻病害[1-3]。该病从水稻苗期到抽穗期均可发生,以分蘖末期至抽穗前期发病严重,一般造成水稻减产6%~40%,病害严重时减产40%以上[4-6]。广东处于高温高湿气候、台风暴雨发生频繁,较适合细菌性条斑病的发生。近年随着田间病菌的积累、带菌稻种的调运,细菌性条斑病在广东呈多发或部分区域呈重发态势,已成为威胁广东水稻高产、优质生产的重要细菌性病害。
水稻细菌性条斑病主要由种子带菌传播,具有传染快、根治难度大等特点。选育和应用抗病品种是防治水稻细菌性病害最经济、有效的方法,而水稻品种的抗病性是寄主与病原物互作的结果,病原菌系的分化研究是抗病资源挖掘以及抗病品种选育的的前提。自1965年起,世界各地学者先后对水稻细菌性条斑病病菌的致病力分化进行了系列研究,1965年Goto最早发现来自菲律宾和日本的细菌性条斑病菌菌株对同一品种的致病力不同[7],1972年Shekhawa根据病菌在10个品种上的抗感反应,将来自不同地区的15个菌株分成了8个小种群,首次提出菌株间致病力存在小种特异性差异[8]。大部分学者普遍认为来自不同地区的条斑病病菌的致病力存在分化,菌株与品种间存在特异性互作现象[9]。但是至今为止国内外研究者对细菌性条斑病病菌的致病力划分仍没有统一的标准,所采用的鉴别寄主、接种方法、病情分级标准并不完全相同[10-19],因此不同地区不同生态稻区鉴别出的结果可借鉴性有限。1992年徐羡明等对广东地区的细菌性条斑病病菌致病力进行过研究[12],但距今26年间未见有关广东地区细菌性条斑病病菌致病力分化和菌系监测的报道,为进一步了解广东地区细菌性条斑病病菌致病力分化和近年来主栽品种的抗性情况,为该病的绿色防控提供科学依据,本研究采用目前国内应用较多的细菌性条斑病菌菌系鉴别品种金刚 30、IR24、IRBB21、IRBB14、IRBB5、IRBB4[20-23]和本地区高感品种五山丝苗为鉴别品种,对分离自广东不同稻区的72个细菌性条斑病病菌致病力进行了测试,并利用优势菌系对18个广东主栽品种的抗性进行了鉴定,明确了广东地区现阶段细菌性条斑病病菌的致病性分化以及优势致病型,为细菌性条斑病抗病资源挖掘、抗病育种以及病害综合防治提供科学依据。
1 材料与方法
1.1 试验材料
1.1.1 供试鉴别品种和主栽品种 细菌性条斑病菌株致病力鉴定的的鉴别品种包括5个近等基因系材料 IRBB4、IRBB5、IRBB14、IRBB21、IR24、金刚 30[20-23]和五山丝苗,其中金刚30和五山丝苗为高感品种。广东主栽品种18个,其中9个常规稻(新银占、合美占、粤晶丝苗2号、玉香油占、白香占、金农丝苗、美香占2号、黄华占、五山丝苗)、9个杂交稻(广8优2168、广8优169、深优9516、天优3618、华优665、深两优58香油占、五优308、深优9708、天优华占)。
1.1.2 菌株来源 水稻细菌性条斑病菌致病力分化测试菌株来自广州、阳春、茂名、新会、兴宁、惠州、广宁、信宜、揭西、龙川、雷州、澄海、肇庆等地,共72个菌株,由广东省农业科学院植物保护研究所于2016—2017年采集、分离和保存。
1.2 试验方法
1.2.1 菌株接种与调查 2017年晚造将供试品种播种在搪瓷盆内,每个品种种植量为15~20粒,每个盆内种植24个品种,每个菌株4次重复。接种时期为秧苗大部分长至4叶龄左右,采用人工喷雾方法,每盆喷菌液量为20~30 mL,以叶片布满液珠为止。接种菌株在NA培养基28℃条件下培养,接种菌龄为48~72 h,以麦法兰氏分度计比浊法,配制3×108CUF/mL细菌悬浮液进行接种。接种后置于保湿培养箱中24 h,温度为28℃,湿度保持在95%以上。然后放于恒温室中定时喷雾保湿,温度设置为28~32℃,21 d后当高感品种发病充分时调查各鉴别品种和主栽品种抗感情况。
1.2.2 品种抗感评价标准 按病斑特征和国际水稻所分级标准综合评价鉴别品种的抗感性,抗性分级标准:高抗(HR),无病征;抗(R),病斑占叶面积小于1% ;中抗(MR),病斑占叶面积1%~5%;中感(MS),病斑占叶面积6%~25%;感(S),病斑占叶面积26%~50%;高感(HS),病斑占叶面积51%~100%[24]。品种叶片呈现水渍状、黄色、扩展长条斑,菌脓较多为感病反应,品种叶片只呈现黄褐至黑褐色短条斑,菌脓较少,此症状为抗病反应。计算鉴别品种抗性频率:

2 结果与分析
2.1 鉴别品种对测试细菌性条斑病菌的抗性频率
根据72 个水稻细菌性条斑病菌菌株在7个鉴别品种上的致病性反应计算各鉴别品种的抗性频率。金刚30、五山丝苗、IRBB4、IR24、IRBB21、IRBB14、IRBB5对 72个 水 稻 细 菌性条斑病菌菌株的抗性频率依次为11.11%、11.11%、38.89%、40.28%、48.61%、63.89%、97.22%。结果表明各近等基因系材料对测试菌株的致病反应存在一定差异,IRBB5可抗绝大部分测试菌株,说明所测试菌株中存在强致病型菌株,IRBB14、IRBB21抗频中等,IRBB4、IR24相对较差,而金刚30、五山丝苗对大部分菌株都表现感病,说明本研究选用的这一套鉴别品种和鉴别方法对广东地区的细菌性条斑病病菌具有较好的鉴别力,鉴别结果可靠、有效。
2.2 广东细菌性条斑病菌菌株致病型
根据72个菌株在7个鉴别品种上的抗感反应,将测试菌株划分为C1~C19共19个致病型(表1)。C17、C18、C19为强致病力菌株,占测试菌株的27.78%;C6、C7、C8、C9、C10、C11、C12、C13、C14、C15、C16 为中等致病力菌株,占测试菌株的38.88%;C2、C3、C4、C5为弱致病力菌株,占测试菌株的27.78%;C1为无致病力菌株,占测试菌株的5.56%。其中优势致病型为C18(SSSSRSS)和C5(SRRRRRS),分别占25%、23.61%,强毒菌系C19(SSSSSSS)占1.39%。由此可见,广东地区的细菌性条斑病菌菌株的致病力分化明显,水稻品种与菌株之间存在致病专化性现象,强毒菌系已出现,应加强监测。
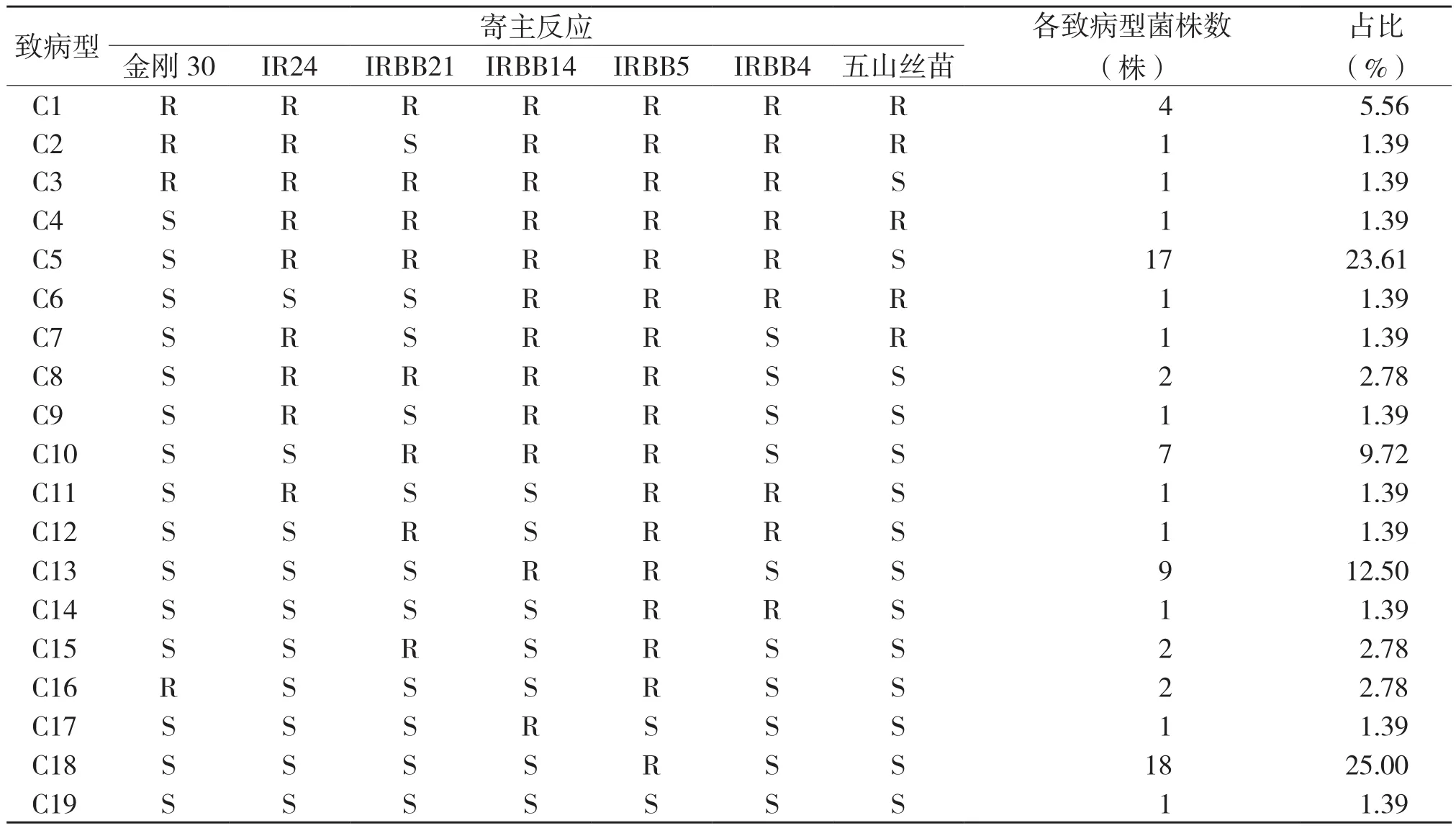
表1 广东细菌性条斑病菌菌株19个致病型在7个鉴别品种上的反应模式
2.3 强致病力菌株的地域分布及其对主栽品种的抗性
C17(SSSRSSS)、C18(SSSSRSS)、C19(SSSSSSS)为广东地区的强致病力菌株,目前主要分布在广州、惠州、阳春、茂名、新会、广宁、雷州地区,其中C17分布惠州1株,C18分布广州4株、阳春4株、茂名3株、新会3株、惠州2株、广宁1株、雷州1株,C19分布阳春1株,尤其是阳春地区已出现强毒菌系C19(SSSSSSS),其致病谱广、毒性强,应加强监测。同时主栽品种美香占2号、五优308、黄华占、广8优169、粤晶丝苗2号、广8优2168、华占均对以上3种强致病力菌株表现感病。因此对这些品种在以上地区的布局需慎重和加强防治。
2.4 优势致病型菌株对广东主栽品种的抗性
18个广东主栽品种对优势致病型菌株C18(SSSSRSS,来自茂名)均表现感病(表2),其中中感、感、高感品种数分别为1、12、5个,只有新银占表现中感。由此可见,目前广东主栽品种对细菌性条斑病的总体抗性较低。
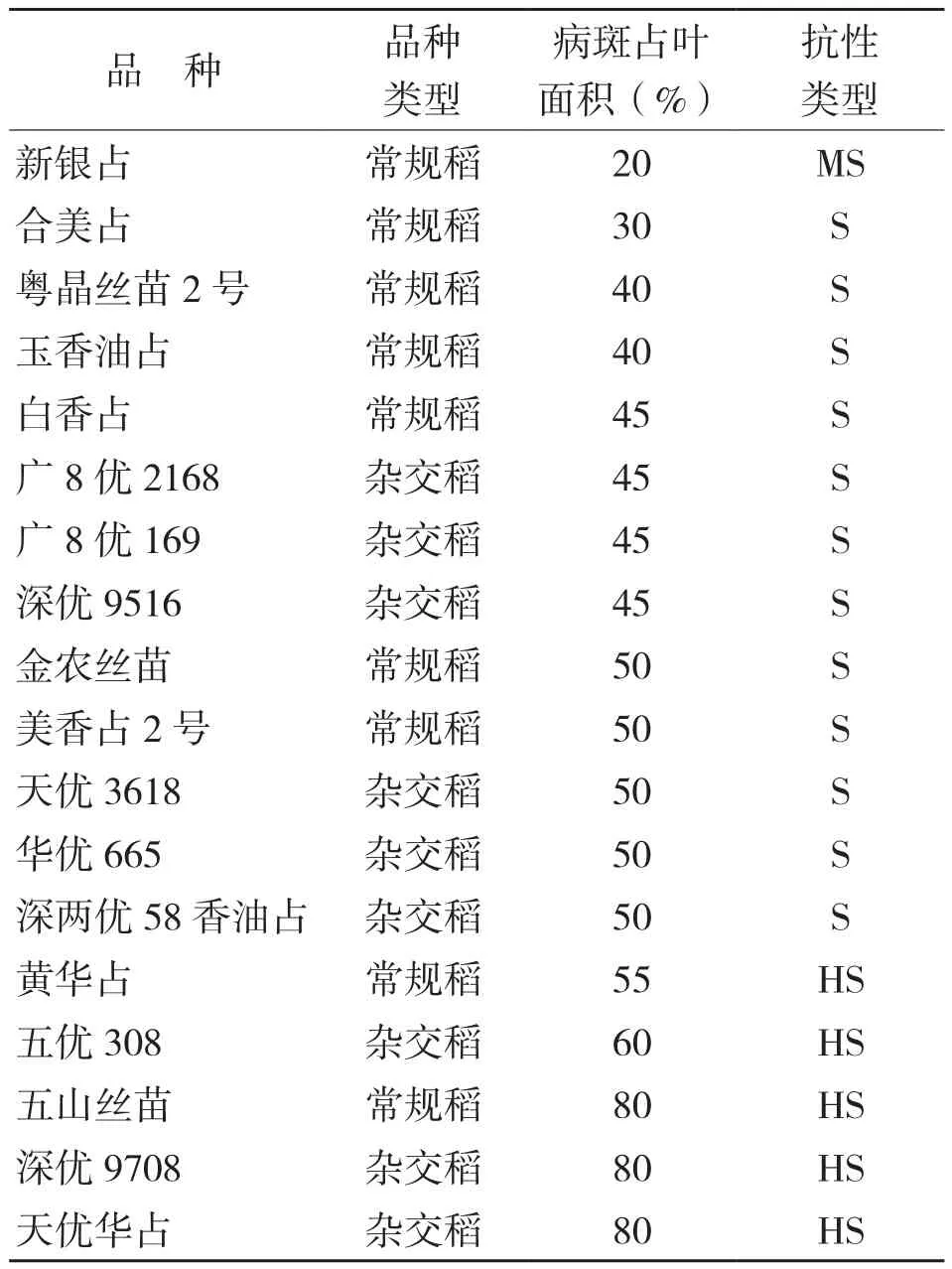
表2 优势致病型菌株对广东部分主栽品种的抗性鉴定结果
3 结论与讨论
水稻细菌性条斑病菌主要通过气孔侵入叶肉薄壁组织增殖侵染水稻叶片,有别于白叶枯病菌主要从伤口侵染水稻,因此水稻细菌性条斑病的接种方法更具多样化,如喷雾法、针刺法、渗入法等。国内已有多篇研究论文证明喷雾法与针刺法、苗期鉴定和成株期所鉴定的结果具有较高的相关性[25-33]。在细菌性条斑病菌致病性分化研究中,Adhikari等[34]、徐羡明等[12]、种藏文等[13]、向建国等[17]与本研究一样采用喷雾法进行接种,其中Adhikari等[34]、徐羡明等[12]、种藏文等[13]与本研究一样以病斑占叶面积的百分率来评价菌株毒力,均能使感病的品种充分发病,证明细菌性条斑病菌菌株间存在分化和分成不同的致病型,与本研究结果一致。Adhikari等[34]以8个IR品种将来源于菲律宾的32个菌株分为3种毒力类型,徐羡明等[12]以DD1OO、DV85、BJ1、南粳 15、辛尼斯、IR8和水源290等7个鉴别品种将来自广东的31个菌株分为6个菌群,种藏文等[13]以V64、010、龙广4号将来自福建的108个菌株分为3个菌系群。因此,应用苗期喷雾接种法和病斑占叶面积的百分率来评价菌株毒力是可行、简便和具有一定可靠性的。而在多次的苗期喷雾接种试验中,我们发现只要是表现抗病的品种,其病斑均表现为长度较短、菌脓少或没有、病斑边缘黑褐色,甚至没有病斑;而感病品种病斑呈现水渍状、黄色、扩展成长条斑,菌脓较多。因此,本研究将病斑特征和病斑占叶面积的百分率一起作为品种的抗感评价标准,使研究结果较准确可靠。
在细菌性条斑病菌致病性分化研究中,近年来国内多采用金刚30、IR24、IRBB21、IRBB14、IRBB5、IRBB4等为鉴别品种。何涛等[20]将来自安徽省的72个菌株划分为6个致病型,张荣胜等[21]将来自江苏、云南、江西、湖南和安徽等省的69个菌株划分为13个致病型,郭亚辉等[22]将来自南方稻区的62个菌株划分为6个小种(致病型),李信申等[23]将江西省的129个菌株划分为9个致病型。其中何涛等、张荣胜等、李信申等研究的菌系中均出现了强毒菌系(SSSSSS),安徽、江苏、云南、江西、湖南等省份均有分布,而强致病力菌株(SSSSRS)在张荣胜等、李信申等研究的菌系中均有出现,且李信申等研究表明强致病力菌株(SSSSRS)是江西省的优势致病力菌株。以上研究者采用的人工接种法是针刺法,而本研究采用的接种方法是喷雾法,两种方法的研究结果均表明细菌性条斑病菌致病性存在明显分化。国内不同的研究表明,细菌性条斑病菌强致病型(SSSSRS、SSSSSS)菌株已经出现,在不同地域的比例不同。
在本研究中亦发现近年来国内采用的鉴别品种也有一定的局限性,需要进一步改进:(1)金刚30均比其他几个近等基因系品种的生育期要短,早抽穗,而五山丝苗的生育期与其他几个近等基因系品种的生育期更相近,且对菌株的抗性频率与金刚30相当,甚至更感病,因此可考虑将五山丝苗替代金刚30;(2)目前这套鉴别品种只有籼稻品种,没有粳稻品种;(3)有些鉴别品种对病菌的鉴别力差异不大,在本研究中,IRBB4、IR24、IRBB21的鉴别力较相近(72个测试菌株中3个品种表现抗的菌株分别为28、29、30个,其中IRBB4与IR24共同表现抗的菌株有26个,IRBB4与IRBB21共同表现抗的菌株有24个),因此将来应研究筛选差异更明显的鉴别品种。
广东是我国的主要水稻产区之一,同时也是最适合细菌性条斑病菌生长的区域。本研究结果表明该地区的优势致病型为强致力菌株,分布广泛,强毒菌系已出现,且主栽品种的总体抗性较低,抗性品种缺乏。因此,以后应在加强监测病菌致病力变化动态的基础上,加快抗性资源的挖掘以及抗病品种的应用,为广东地区细菌性条斑病的防控提供有力保障。
