隆肛蛙体长及其两性异形的海拔变异研究*
2018-10-18钟茂君尹清孝史清茂章世鹏廖文波
金 龙 钟茂君 罗 怡 尹清孝 史清茂 章世鹏 张 融 廖文波
(1.西华师范大学西南野生动植物资源保护教育部重点实验室,四川 南充 637009; 2.米仓山国家级自然保护区管理局,四川 旺苍 628200; 3.西华师范大学两栖爬行类生态适应研究中心,四川 南充 637009)
自 1847 年贝格曼提出温血动物在寒冷的环境比在暖和的环境有更大的身体以来,大量的生态学者对动物体长与环境梯度变化的相互关系进行了研究[1-6].贝格曼认为身体大的动物在寒冷的环境中可以降低表面积从而导致能量散失减少,其可视为动物的热量保护机制.然而,部分学者对贝格曼定律提出了质疑[7],他们认为贝格曼定律作为动物的热量保护机制是无效的[7].近年来部分研究证明了哺乳动物和鸟类体长的海拔和纬度变异与贝格曼定律一致[2, 8].然而,研究者对变温动物体长的海拔和纬度变异也进行了验证,部分学者发现变温动物的体长变异与贝格曼定律一致[9-10],但另一部分学者发现变温动物的体长变异与贝格曼定律相反[11-13].
关于两栖动物体长的地理变异特征,Ashton发现 9 种无尾类体长变异基本与贝格曼定律一致,仅 2 种有尾类体长的海拔和纬度变异明显符合贝格曼定律[14].同样,华西蟾蜍(Bufoandrewsi)、华西雨蛙(Hylaannectanschuanxiensis)、四川湍蛙(Amolopsmantzorum)和泽陆蛙(Fejervaryalimnocharis)的体长随海拔变异满足于贝格曼定律[15-18],黑斑蛙(Rananigromaculata)体长的地理变化与贝格曼定律相反[19],绿臭蛙(Odorranagrahami)[20]、高原林蛙(Ranakukunoris)[21]体长的地理变化与贝格曼定律不一致.因此,检验两栖动物体长的地理变异是否符合贝格曼定律需要更多的物种进行验证.
动物雌性和雄性的体长存在差异性,并且这种差异性存在显著的变化[22-25].两种主要假设用于解释体长两性异形的变化,性选择假说认为雄体间强烈的竞争导致雄性偏大的两性异形,其变化与伦斯法则一致[26-27];相反,生殖力选择假说认为增加雌性的生殖力导致雌性偏大的两性异形,其变化与反伦斯法则一致[28-30].廖文波等对华西蟾蜍、中国林蛙和泽陆蛙不同地理种群的体长两性异形的地理变异和 39 种两栖动物类群体长两性异形进行了系统研究,结果发现:两栖动物均为雌性偏大的体长两性异形.华西蟾蜍、中国林蛙和泽陆蛙雌雄体长的差异性(SSD)的变化特征与伦斯法则一致[30-32],其原因是生殖力选择导致更大的雌体,而且两性的年龄差异也是导致体长差异的主要因素;相反,两栖动物类群体长两性异形不符合伦斯法则,其原因是性选择对雄性体长的影响抵消了生殖力选择对雌性大小的影响[33].由此可见,两栖动物雌雄体长的差异性的进化特征是否符合伦斯法则是由性选择和生殖力选择的共同作用所致.
隆肛蛙(Feiranaquadranus)是中国特有两栖类[34].成体栖息于河流、水沟和积水坑,也见于河边山林中,白天多伏于较大石下[35].目前对隆肛蛙的分布、生境状况、生活史和遗传分化有部分报道[34-36].笔者于 2017 年 3—8 月对隆肛蛙体长及两性异形的海拔变异进行了研究,检验了体长的海拔变异是否与贝格曼定律一致以及体长两性异形变化是否符合伦斯法则.通过对隆肛蛙体长变异及两性异形演化的研究,能够为两栖类生活史性状的地理变异机理提供理论指导.
1 材料与方法
1.1 研究区域
四川米仓山国家级自然保护区地理位置介于北纬32°29′~32°41′,东经106°24′~106°39′ 之间.地处四川省广元市旺苍县境的东北部,位于米仓山—大巴山山脉西段南坡.行政区域包括鼓城乡(鼓城村、金竹村、跃进村、关口村)和檬子乡(柏杨村、店坪村).保护区总面积为23 400 hm2(核心区面积为9 203.1 hm2,缓冲区面积为4 057.8 hm2,实验区面积为10 139.1 hm2).保护区属中亚热带湿润季风气候,受地理地貌的影响,四季分明,雨量充沛(年平均降雨量1 200 mm),光热资源丰富(年平均温度16.2 ℃,年平均日照时数为1 352.52 h),无霜期较长(全年无霜期约为260 d),山地气候明显[37].
1.2 研究方法
2017年3—8月,采用实地样线法对米仓山国家级自然保护区内3个不同地理种群及保护区附近的1个种群的隆肛蛙进行采集(表1),在每个地理种群,首先沿河(溪)流随机布设样线,沿样线行进,夜间仔细搜索样线两侧的隆肛蛙,样线的宽度为3~10 m,长度在1~5 km,样线上行进的速度根据
调查工具确定,步行宜为1 km/h.发现隆肛蛙时,记录数量并抓捕个体,记录个体的地理位置,然后将所有个体带回野外实验基地,鉴定个体的性别,使用游标卡尺测量个体体长.
我们利用一般线形模型(generalised linear models,GLM)以体长为独立变量,海拔和性别为固定因素来检测体长的地理变异是否符合贝格曼定律.同样利用海拔和性别的交互作用对体长影响来检验两性异形的变化是否与伦斯法则一致.使用压轴回归分析(reduced major axis regression, RMA)以雌体体长为因变量,雄性体长为自变量来分析两性体长的异速生长关系,当βRAM> 1 时,两性异性的变化与伦斯法则一致,相反,当βRAM< 1 时,其变化与伦斯法则相反[22].
2 结 果
调查期间在 4 个研究种群共采集了 111 个个体,其中雄体 55 个,雌体 56个,性比差异不显著(χ2=0.009,df=1,P=0.924).每个种群隆肛蛙的雌性和雄性体长和性比差异显著(表1;所有P< 0.05).不同地理种群的隆肛蛙体长存在显著的差异性(F3,110=5.853,P=0.001),两性的体长差异不显著(F1,110=1.622,P=0.206),雌性和雄性的体长并不随海拔增加而增加(图 1),其与贝格曼定律不一致.海拔和性别交互作用对体长的影响显著,表明两性异形存在海拔变化(F3,110=2.719,P=0.048).雌性体长和雄性体长异速生长显著(βRAM=0.627),隆肛蛙两性异形随体长增加而减少(图2),其与伦斯法则相反.

表1 米仓山国家级自然保护区隆肛蛙不同地理种群的采样点的地理位置、样本数和体长
3 讨 论
如果 50% 以上的温血动物在更冷的地方有更大的身体,那么温血动物身体大小的地理变异符合贝格曼定律[38].根据该标准,Ashton 发现超过 50%的两栖动物身体大小的地理变异满足贝格曼定律[14].以前的研究表明,部分物种不同地理种群的体长海拔变异与贝格曼定律一致[15-18],少量物种的体长海拔变异与贝格曼定律相反[19],其原因归结于高海拔寒冷环境温度导致更大的身体,低海拔暖和的环境温度导致更小的体长[39],这也与贝格曼最早提出的动物热保护机制一致.身体大的个体在寒冷的环境中可以储存更多的能量来提高存活力和越冬能力[40].然而,本研究发现隆肛蛙不同种群的体长并未随海拔的增加而减少,这与贝格曼定律不一致.两栖动物体长的海拔差异应考虑遗传因素的影响[41].因此在研究两栖动物不同地理种群的体长变异是否符合贝格曼定律时,必须考虑遗传因素对隆肛蛙体长的影响.本研究并未考虑遗传因素的影响,将来需要进一步探讨遗传因素对其体长的影响.此外,低的发育温度将导致变温动物有更大的变态幼体[42],而且温度对体长的影响可以解释两栖动物在寒冷的地区有更大的体长.
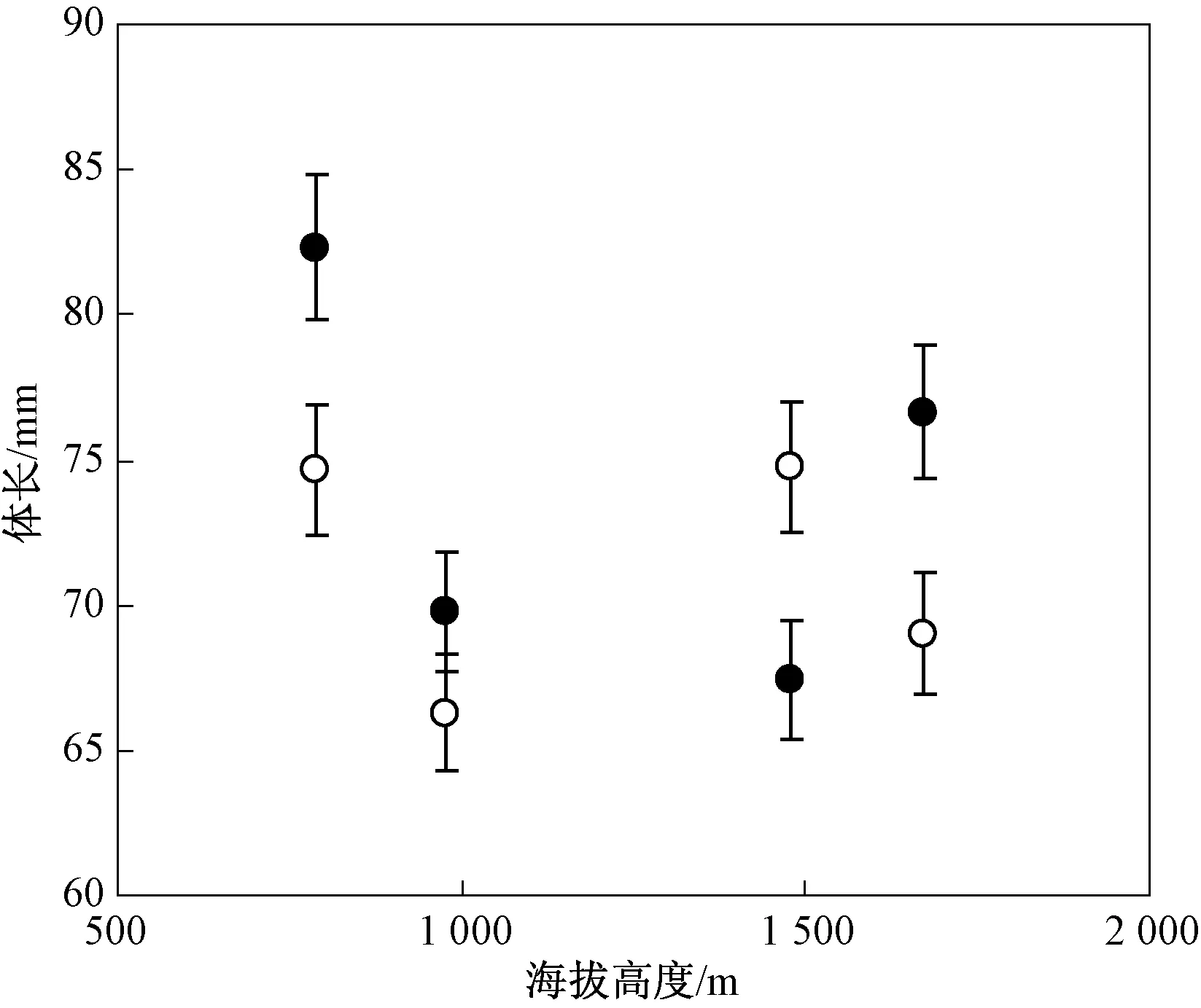
图1 隆肛蛙不同海拔种群的身体大小的变化(雌性:实心圆;雄性:空心圆)
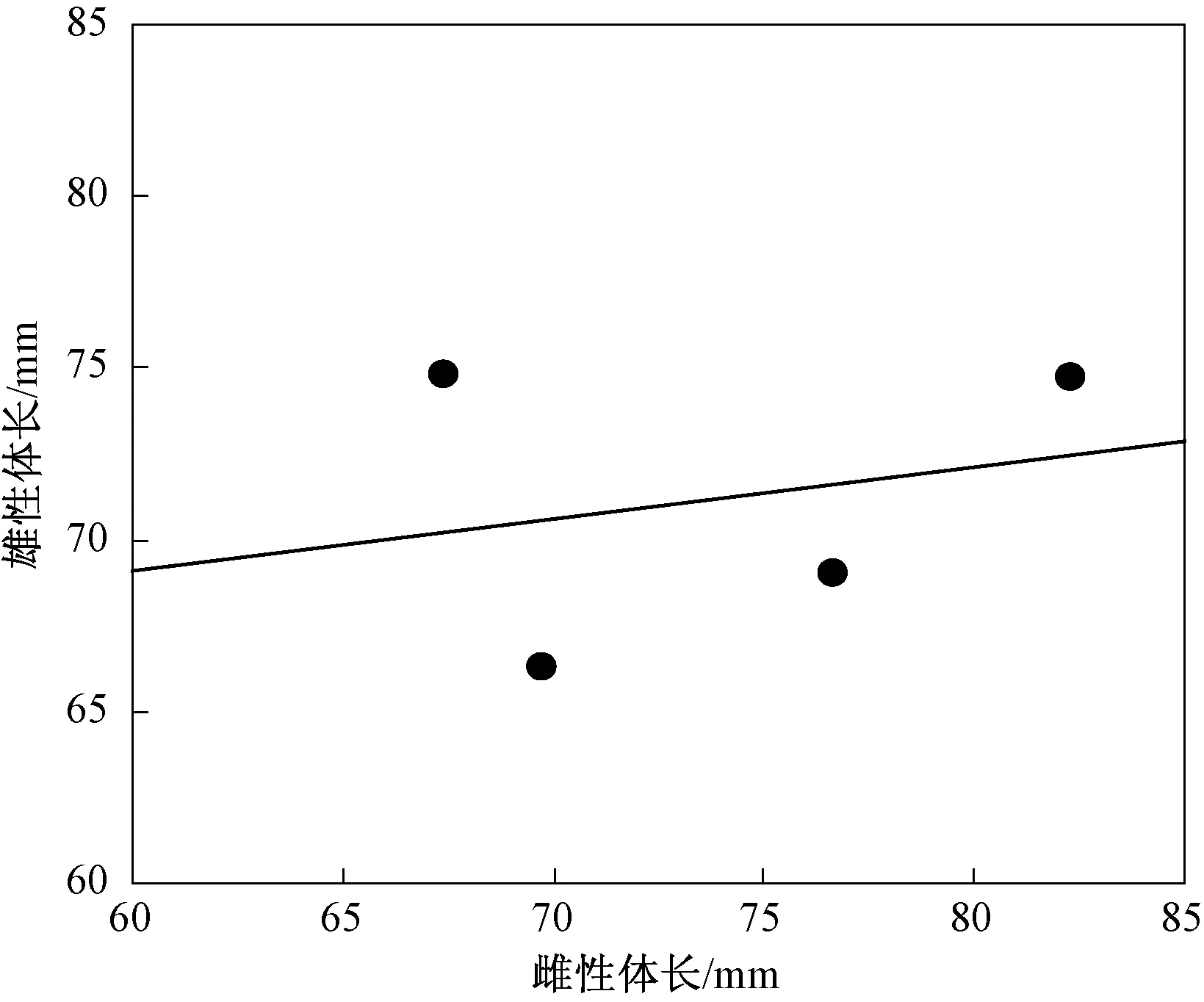
图2 隆肛蛙不同海拔种群雌性体长与雄性体长的异速生长关系(βRAM=0.627)
以前研究表明个体年龄也明显影响体长,当控制个体年龄对体长的影响时,其体长的海拔变异仍然符合贝格曼定律[18-19].由于本研究未对个体进行年龄鉴定,因此未控制年龄的影响,然而大量研究都表明两栖动物年龄结构对研究体长变异与贝格曼定律的关系影响不明显[17-19, 43].
除了温度对两栖动物体长影响外,环境因素(湿度和降雨量)也影响两栖动物的体长.由于两栖动物需要湿润的环境来保持皮肤呼吸,它们的体长受空气湿度限制[44].由于相对减少的表面积,身体大的个体对干燥有更大的忍受性[44].同样水的利用性也影响个体的体长,干燥地区的两栖动物有更大的身体[45].因此,在研究两栖动物体长的海拔变异时,我们还应该考虑大气湿度、水利用性和降雨量等因素的影响.
廖文波等对 39 个两栖类物种的两性异形进行了系统研究,结果发现,体长两性异形不符合伦斯法则[33],其原因是性选择对雄性体长的影响抵消了生殖力选择对雌性大小的影响.然而,华西蟾蜍、中国林蛙和泽陆蛙的不同地理种群的体长两性异形的地理变异与伦斯法则一致[18, 30, 32],其原因是由于生殖力选择大的雌性引起的.本研究发现隆肛蛙不同种群体长两性异形的变化与伦斯法则相反,这可能是生殖力选择更大的雌体导致的.由此可见,两栖动物不同地理种群体长两性异形的进化特征是否符合伦斯法则是由性选择和生殖力选择的共同作用决定的.
