稻米储藏品质劣变机制研究进展与展望
2018-10-18费月新曹玉洁吴敏吴洪恺
费月新 曹玉洁 吴敏 吴洪恺
(浙江农林大学农业与食品科学学院,杭州311300;*通讯作者:hongkaiwu@163.com)
水稻是我国的主要粮食作物,全国近三分之二的人口以稻米为主食。基于稻谷生产的季节性,为了满足人们的常年需求以及粮食安全的需要,不得不进行相当规模的储备。另外,从战略和防灾角度考虑,国家必须实施国储制度,强化宏观调控能力。长期以来,国家、地方粮食储备在以丰补欠、平抑粮食价格等方面起了积极的作用,为我国社会稳定、经济发展做出了积极贡献。据统计,目前我国稻谷库存量约占全年稻谷产量的40%,每年超过千亿公斤的稻谷需要储藏,平均储藏时间约16个月。稻谷作为一个有生命的活体,在储藏期间会进行呼吸作用等生化反应,并且受储藏条件的影响,稻谷本身也会发生一系列的变化,导致品质下降,即为大米的陈化。我国每年稻米储藏过程中因陈化造成的损失占总储存量的3%以上,在高温高湿地区(如浙江省)陈化尤为严重,损失更大,给国家造成了重大经济损失。国内外学者对稻米陈化的原因进行了广泛深入的研究,并取得了重要进展。
1 稻米储藏过程中化学组成的变化
稻米主要由淀粉、蛋白质和脂肪等物质组成。陈化过程中,这3类物质总量上变异不显著,但它们的化学组成发生了明显的变化[1]。淀粉是稻米的主要成分,占精米干质量的90%左右[2],因此,稻米的淀粉含量及其特性对稻米储藏品质起至关重要的作用。稻米淀粉特性和直链淀粉含量对稻米含水量的影响很大,而含水量是影响稻米储藏品质的最重要环境因素之一。因此,稻米淀粉的构成类型可在一定程度上决定储藏品质[3]。张淑娟等[4]认为,储藏1年后,稻米的总直链淀粉含量无显著变化,但不溶性直链淀粉含量则逐渐增加。邱明发等[5]认为,不溶性直链淀粉含量增加的原因是稻米在储藏过程中淀粉脱分支酶仍保持其活性,并作用于1,6-糖苷键使支链淀粉脱支,所以储藏后不溶性直链淀粉含量增加,支链淀粉含量减少。Rehman等[6-7]研究发现,陈化稻米中的还原糖含量略有增加。Patindol等[8]研究表明,陈化稻米的支链淀粉的短链(DP=6-12)比例增加。Huang等[9]研究了内源性淀粉酶对糯米淀粉精细结构的影响,发现在较高温度条件下,储藏较长一段时间后,长链比例下降,短链比例增加。这些研究结果都一致表明,稻米储藏期间可能发生了淀粉的降解。
储藏蛋白是稻米的第二大成分,占精米干质量的7%~10%,根据溶解度的不同可分为4种蛋白,即清蛋白、球蛋白、醇溶蛋白和谷蛋白[10]。在大多数谷物中,醇溶谷蛋白是主要的贮藏蛋白,而在水稻中谷蛋白是主要的储藏蛋白,占总储藏蛋白的60%~80%[11]。有研究提出,与新鲜稻米相比,陈化稻米的贮藏蛋白(丙醇提取的醇溶蛋白)含量减少,推测陈化过程中贮藏蛋白通过二硫键或其他活性基团形成了更大的蛋白分子[12],或改变了蛋白质结构,进而影响蛋白质的性质,如溶解性、热性能和亲水性/亲油性等[13]。Teo等[14]研究表明,新鲜稻米分离出的淀粉中添加谷蛋白,其糊化特性和老化稻米相似。对老化大米样品进行β-巯基乙醇处理后与未处理的样品相比,峰值黏度显著增加(P<0.01),可能是由于β-巯基乙醇处理后蛋白质二硫键断裂,导致更多的水渗透和淀粉颗粒的肿胀,从而增加糊化程度[15]。同样,谢新华等[16]用二硫苏糖醇处理打破稻米中蛋白质的二硫键后,发现稻米淀粉完全糊化的时间缩短,糊化温度降低,推测二硫键结合形成的蛋白质网络的变化引起了稻米吸水性改变。
稻米中脂肪约占精米干质量的0.8%左右。脂肪酶(Lipases,E.C.3.1.1.3)是一种甘油酯水解酶,可以水解脂肪分子中的甘油酯键,将脂肪甘油三酯水解为甘油及游离脂肪酸,是脂肪分解代谢中第一步反应的关键酶[17]。Takano等[18-19]证实,稻米在储藏过程中发生了脂肪的降解,产生大量的游离脂肪酸(包括饱和脂肪酸和不饱和脂肪酸)。Tsuzuki等[20]研究表明,陈化稻米的饱和脂肪酸和不饱和脂肪酸总含量与新鲜稻米的差异不显著,但陈化稻米的不饱和脂肪酸含量显著低于新鲜稻米;新鲜稻米中亚油酸占脂肪酸总量的50%左右,而陈化的稻米中油酸含量较高,亚麻二烯酸和亚麻三烯酸含量较少[21],表明在高温储藏条件下不饱和脂肪酸是不稳定的,会发生氧化。有研究证实,储存期间脂肪的氧化对稻米的陈化起重要作用,脂肪氧化酶-3(Lox-3)作为关键酶催化这个反应[22]。Hatanaka等[23-24]研究表明,稻米中脂肪氧化酶(lipoxygenase,E.C.1.13.11.12,简称Lox)主要以亚油酸和亚麻酸为底物,降解生成亚油酸氢过氧化物和亚麻酸氢过氧化物,并且在反应过程中产生醛、酮等挥发性腥臭难闻的物质。基于生物信息学分析,水稻基因组编码14个Lox蛋白[25],但是只有 Lox1、Lox2、Lox3 三种同工酶[26-28]和 HILox[29-30]被克隆和证实。Huang等[28]研究表明,与野生型种子相比,过表达OsLox2的种子在正常环境下能快速发芽,但是在人工老化的环境下发芽会变慢;RNA干涉后,结果却相反,说明Lox2和种子的寿命密切相关。日本科学家于1992年用单克隆抗体鉴定出能提高水稻耐储藏性的脂肪氧化酶Lox-3缺失体,可在一定程度上提高稻米的耐储藏性。Xu等[31]对水稻Lox3反义抑制的研究发现,降低Lox3表达水平能延长水稻储藏时间,进一步表明lox-3基因的耐储藏功能。中国科学家对lox-3基因的生产应用进行了大量的研究,但至今未有大面积推广的报道。
酚酸是稻米中的一种微量成分,在储藏中表现出一定的功能。在稻米储藏期间,结合酚酸含量没有显著变化,而陈化稻米中游离酚酸含量比新鲜稻米低(P<0.05),推测在陈化过程中,游离酚酸和不饱和脂肪酸之间发生了化学反应[32]。Juliano等[33]以10个国际水稻研究所育成的水稻品种为材料,研究发现谷壳的抗氧化物质酚的组成水平与耐储性呈正相关,而且这些酚对于水稻种子抗衰老所起的作用甚至超过维生素E和维生素B。
Khov Kuong等[34]对水稻中的锌、铁、维生素A在高温高湿环境储藏后发生的变化研究表明,高温高湿储藏环境下,锌、铁含量没有变化,但是维生素A的含量减少了90%。
2 稻米储藏过程中淀粉和非淀粉成分之间的相互作用
Shibuya等[35]研究表明,陈化稻米与新鲜稻米的米粉糊化特征是不同的,但相应的淀粉糊化特征相似,暗示淀粉和非淀粉成分之间的相互作用影响了稻米理化特性。他们进一步研究认为,储存过程中稻米胚乳细胞壁被内源木聚糖酶分解,导致淀粉谱特征发生了变化。因此,仅对单一组分研究难以解释稻米在陈化过程中质构特性的变化,而且稻米作为一个有机体,其各种组分是不可能孤立存在,而是相互作用,相互影响。Zhou等[36-37]研究发现,新鲜稻米样品用半纤维素酶处理后,峰值粘度(RVA)和峰值温度(DSC)有所下降,然而,通过对老化稻米样品半纤维素酶处理前后的对比发现,整个糊化曲线无显著性差异;用纤维素酶处理陈化水稻样品,与未经处理的样品相比,RVA和DSC曲线的特征均产生显著差异。Briffaz等[38-39]推测,水稻的细胞壁对稻米结构和蒸煮特性有至关重要的作用,细胞壁残留参与了陈化过程,纤维素酶处理改良了老化水稻细胞壁结构并增强了RVA和DSC在运行时水的渗透性。
Baldwin等[40]用二次离子质谱发现了小麦和大米淀粉表面脂质的存在。姜培彦[41]认为,淀粉表面脂质会使淀粉溶胀性降低,并导致直链淀粉含量溶出减少,降低糊化温度和热焓值(enthalpy),并且对淀粉流变性质也会产生影响。谷物在陈化时,脂质过氧化会产生大量的活性丙二醛(MDA)[42]。Wu等[43]以脂质过氧化的产物MDA作为对蛋白质结构特性影响的指标,发现MDA含量增加,则蛋白质的溶解度、表面疏水性降低。
3 稻米储藏过程中物性学变化
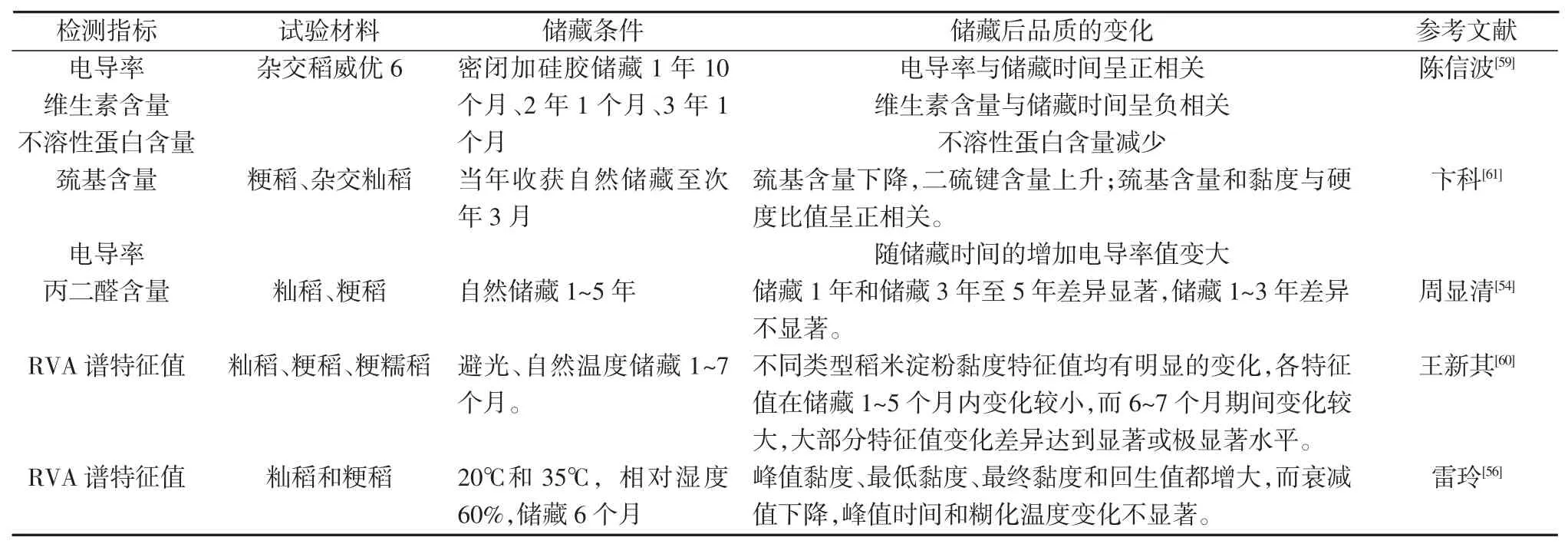
表1 稻米储藏品质评价指标及其效果
有研究指出,陈化稻米更难与水结合、膨胀,并且在热水裂解时淀粉颗粒对水破坏性表现高抗,导致糊化过程中大米滤出更少的淀粉成分[37,44]。陈化导致了淀粉微晶束结构加强,即在加热过程中当水分子渗入到淀粉的结晶区和非定形区时,膨胀力不足以使这一部分具有加强结构的淀粉分子发生结构破裂和转移(糊化以及溶出)[45]。稻米的蒸煮特性和流变性能的表现十分广泛,很大程度上是由膨胀、淀粉糊化和回生特性决定[1,46]。陈化过程中稻米的最敏感指标之一是糊化特性的变化[47-49]。快速粘度分析仪(RVA)已广泛用于稻米糊化特性的评价。RVA谱一般用7个特征值来描述,分别是峰值黏度(peak viscosity,PKV)、热浆粘度(hot paste viscosity,HPV)、冷胶粘度(cool paste viscosity,CPV)、崩解值(breakdown viscosity,BDV,最高粘度-热浆粘度)、消减值(setback viscosity,SBV,冷胶粘度-最高粘度)、糊化温度(pasting temperature,PaT)和峰值时间(peak time,PeT)。研究表明,峰值粘度(PKV)和崩解值(BDV)是评价大米陈化最敏感的指标[9,50]。RVA分析表明,高温储藏后,稻米的PKV和BDV增加[8,51]。宋伟等[52]以粳稻为材料研究表明,储藏温度和时间对峰值粘度的影响均达极显著水平,峰值粘度随着储藏时间的延长呈上升趋势,储藏温度越高,上升趋势越明显。RVA和DSC数据表明,高温储存引起的稻米陈化导致淀粉颗粒与水化合和膨胀更加困难[50]。
4 储藏品质劣变机制研究展望
如上所述,前人对稻米陈化的谷物化学和物性学进行了大量的研究,表明稻米陈化是一个复杂的过程,淀粉、蛋白质、脂类以及各成分互作对储藏品质均有重要作用。稻米陈化过程中不同品种之间的糊化特性变化明显不同,一般表现为籼稻比粳稻耐储藏[8-9,53],说明稻米储藏特性存在基因型差异,是由遗传控制的。但是,稻米陈化的遗传学机理研究未见报道。要研究稻米储藏品质的遗传机理,首先必须明确储藏品质的评价指标[54]。
许斌等[55]研究表明,脂肪酸值与储粮品质有很好的相关性,是反映粮食储藏品质变化的灵敏指标。储藏温度高、时间长,脂肪酸含量增加[56]。因此,脂肪酸值是粮食储存品质的重要指标。GB/T20569-2006规定,籼稻不宜存的脂肪酸值阈值为37 mg/100 g,粳稻为30 mg/100 g[57]。GB/T20569-2006的测定方法中,脂肪酸值的测量很容易产生误差。例如,在利用无水乙醇作为萃取剂时会有醇溶性蛋白萃取出来,醇溶性蛋白属于酸性蛋白质,因而测得的脂肪酸值偏高[58]。除脂肪酸值外,电导率[54,59]、丙二醛含量[54]、淀粉 RVA 谱特征值[56,60]、巯基含量[61]等指标都可以用来评价稻米的储藏品质,这些指标反应了稻米储藏前后的变化,呈现了非常复杂的相关关系(表1)。可见,稻米储藏品质是一个复杂的遗传性状。
遗传性状研究所用的群体类型很多,常用的有F2、重组自交系(RIL)以及双单倍体(DH)群体等,利用这些群体研究控制某些性状的QTL数目、染色体上的位置及遗传效应等取得了很好的结果。但是这些群体的遗传背景复杂,在进行分析时存在一定的遗传“噪音”,不适宜用于复杂的遗传性状研究[62]。染色体片段代换系是指一套代换系的供体插入片段相互重叠,可以覆盖整个基因组,每个系与背景亲本只有1个或少数几个供体片段的差异,遗传背景相近,是研究复杂遗传性状的理想群体。明确稻米陈化的遗传机理,可以从遗传上改良水稻的贮藏特性,培育耐储藏的水稻品种,这是一条经济有效的途径。
