宽体金线蛭5个地理群体遗传多样性的RAPD分析及生长指标的比较研究
2018-10-18姜爱兰王信海丁辰龙吴学军唐金玉王飞飞叶建勇
姜爱兰,王信海,丁辰龙,吴学军,唐金玉,王飞飞,叶建勇
(江苏省农业科学院 宿迁农科所,江苏 宿迁 223800)
宽体金线蛭(Whitmaniapigra),隶属于环节动物门(Annelida)有环带亚门(Clitella)蛭纲(Hirudinea)真蛭亚纲(Euhirudinea)无吻蛭目(Arhynchobdellida)医蛭形亚目(Hirudiniformes)黄蛭科(Haemopidae)金线蛭属(Whitmania),别名蚂蟥、肉钻子、水痴马鳖[1],是传统的名贵中药材,现代研究证明其涎液中含有一种由65~66种氨基酸组成的酸性低分子多肽、抗血凝物质,称为水蛭素、肝素、抗血栓素等[2],可治疗中风、闭经、截瘫、心绞痛、无名肿疼、肿瘤、颈淋巴结核等疾病,能使移植手术后的静脉血管保持畅通,其蜂蜜浸出液亦可治疗角膜溃病和角膜云翳,具有广泛的药用价值[3]。近年来,以水蛭为原料的药品及保健品日渐增多,但由于各种天然水体中大量生活和工业污水的排入,农田中大量施用化肥和农药,加上人为地酷渔滥捕,其种群数量急剧减少,自然资源日渐萎缩,几乎很难形成产量,种质资源也受到严重破坏,从而导致其药材及药品价格不断攀升,因此水蛭的人工养殖日趋受到重视[4],也为渔民致富开辟了新的途径。
目前,宽体金线蛭的研究工作大多集中在繁、养殖技术上[5-14],对其种质资源状况的研究还未见报道,为了解各个地理区域种群的遗传背景和遗传差异状况以及遗传差异与生长指标的相关性,本研究开展了不同地区宽体金线蛭遗传差异性分析及生长指标的比较研究,以期为今后宽体金线蛭种质资源保护和遗传育种及繁、养殖技术研究提供科学依据。
1 材料与方法
1.1 试验材料
宽体金线蛭亲蛭和幼蛭于2016年3~5月分别从江苏宿迁的骆马湖(SQ)、江苏兴化的乌巾荡(XH)、江苏吴江的太湖(WJ)、湖北大悟县的竹竿河(DWX)和黑龙江宾县的松花江(BX)野外采集,活体运回试验基地,置于挂在池塘内的60目网箱(规格:2.0 m×4.0 m×1.2 m)内暂养,暂养前1 d投放足量的活田螺,供亲蛭、幼蛭暂养期间摄食。仔蛭来源于亲蛭人工孵化后收集的幼蛭。
1.2 基因组DNA提取
采用CTAB法提取样品基因组DNA。(1)在65 ℃水浴锅中预热 CTAB 提取液;(2)在液氮中迅速研磨样品,将粉末状材料转入2 mL离心管中,加入预热的CTAB提取液(每克样品加入3~5 mL提取液),65 ℃保温30~60 min,每隔 10 min轻轻颠倒混匀;(3)11000 r/min条件下离心5 min,取上清转入新离心管;(4)加入等体积酚/氯仿(1∶1),充分混匀,11000 r/min离心10 min,取上清转入新离心管;(5)加入等体积氯仿,充分混匀,11000 r/min离心10 min,取上清转入新离心管;(6)重复步骤4、步骤5;(7)加入2/3体积的异丙醇混匀,室温放置,沉淀 15 min;(8)11000 r/min条件下离心6 min,弃上清;(9)将沉淀用70%的乙醇漂洗一次,室温条件下11000 r/min离心2 min,弃上清,重复洗一次;(10)提取产物2~3 μL用 2%琼脂糖凝胶电泳检测,其余置于-20 ℃保存备用。
1.3 RAPD分析
1.3.1 RAPD 反应体系和扩增条件 PCR扩增体系(20 μL)为:DNA 模板 2 μL,primer 1 μL,蓝色Mix 10 μL,去离子水7 μL。PCR扩增程序为:94 ℃预变性3 min,然后进行 40个循环:94 ℃变性30 s,36 ℃复性50 s,72 ℃延伸1.5 min;循环结束后72 ℃延伸10 min,4 ℃保存。
1.3.2 RAPD 扩增产物的检测 用30对引物对5个样品进行PCR扩增,引物信息见表1,将PCR产物在2.0%的琼脂糖凝胶电泳中检测。
1.4 生长试验
分别测定仔蛭和幼蛭的生长指标。
仔蛭生长指标测定:分别从不同地理种群同一天孵出的仔蛭暂养槽内随机取180条仔蛭分3组放于编好号的玻璃钢水缸(尺寸:40 cm×50 cm×30 cm)中进行成活率统计和生长指标比较试验,玻璃钢水缸顶部用80目网片封住,水深10 cm左右,内部放2~3段PVC管,供幼稚栖息,每天换水2/3,投喂冰冻轮虫和初孵丰年虫。每隔5 d投喂壳宽3~5 mm左右的活的小田螺,每5 d从每缸内随机取15条仔蛭称重,21 d后取出全部称重,计算平均体质量。
表1 RAPD引物信息
幼蛭生长指标测定:分别从不同地理种群的暂养网箱内随机取120条3~4 g幼蛭平均分3组放于同池塘编好号的60目水蛭养殖专用网箱(尺寸:100 cm×100 cm×110 cm)内进行成活率统计和生长指标比较试验。池塘水温平均24.9±1.2 ℃,pH 7.6~8.4,溶解氧6.13~7.02 mg/L,水深85 cm左右,网箱内部放栽种苦草的花盆2盆,直径110 mm的PVC管2~3段,供幼蛭栖息。每隔5 d投喂壳宽9~12 mm左右的活的田螺,对每条幼蛭称重,计算平均体质量。
生长指标的平均日增重、相对增重率和特定增重率分别按下列公式计算:
平均日增重:AGR/(g/d)=(W2-W1)/(t2-t1)
相对增重率:RGR/%=100×(W2-W1)/[W1×(t2-t1)]
特定增重率:SGR/%=100×(lnW2-lnW1)/(t2-t1)
式中:W2表示蛭第t2天平均体质量(g),W1表示蛭第t1天平均体质量(g)。
1.5 数据统计分析
RAPD分析的PCR扩增产物的电泳位置在凝胶的某个相同迁移率位置上有DNA条带记为1,无DNA条带记为0,如表2,形成RAPD的表型数据矩阵用NTSYS软件计算相似系数(DICE系数),并且按照遗传距离进行UPGMA聚类分析。采用SPSS 13统计软件对试验数据进行单因素方差分析(One-way ANOVA),检验不同地域对宽体金线蛭生长指标影响的显著性,用最小极差法(LSD)中的Duncan氏新复极差法进行多重比较,P<0.05视为具有统计学意义[15-16]。
表2 01数据统计(部分)
2 结果与分析
2.1 RAPD实验检测结果
利用RAPD分子标记技术对5个样品进行遗传多样性分析,30条引物扩增如图1、图2。
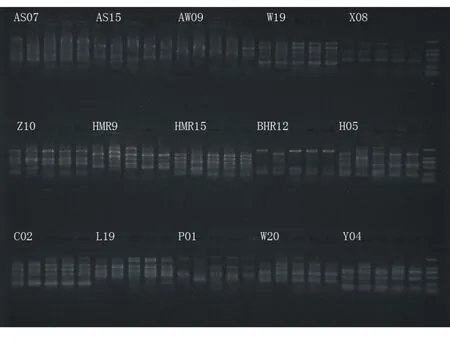
图1 PCR扩增产物检测结果P1~P15(M:DL2000)
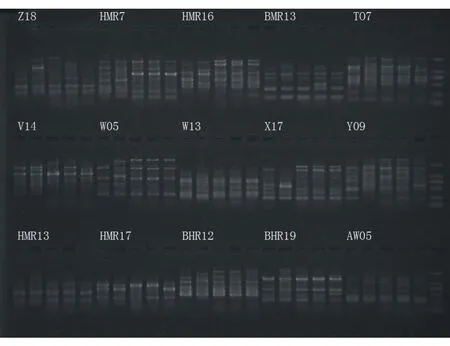
图2 PCR扩增产物检测结果P16~P30(M:DL2000)
2.2 供试材料的亲缘关系分析
利用30对引物在5个样品中获得157条扩增片段,如表2。在NTSYS软件中计算样品间的遗传相似系数(GS值),得到供试材料遗传相似矩阵。遗传相似系数越大,表明亲缘关系越近,遗传相似系数越小,表明亲缘关系越远。由表3可知,5个样品的GS值范围为:0.6433~0.7707,变幅为0.1274。其中江苏兴化样品和黑龙江宾县样品之间的遗传相似系数最小,为0.6433,表明这两个样品之间的亲缘关系较远,遗传差异性大。江苏兴化样品和江苏宿迁样品之间的遗传相似系数最大,为0.7707,表明江苏兴化样品和江苏宿迁样品之间的亲缘关系较近,遗传差异性较小。
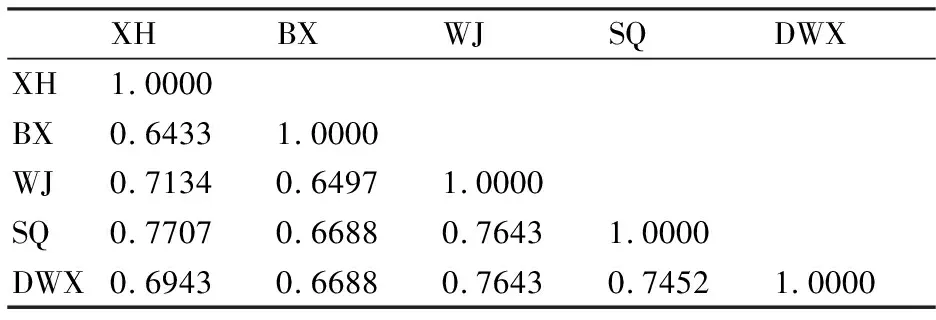
表3 5个不同地理群体之间的遗传相似系数矩阵
2.3 RAPD分子标记聚类分析
利用UPGMA法进行聚类分析,根据遗传相似系数对各样品进行分类分析,聚类图如图3,从得到的聚类分析图上可以看出,5个样品分成2类,江苏兴化、江苏宿迁、江苏吴江、湖北大悟县是第一类,黑龙江宾县是第二类。其中第一类又分为2个亚类,第一亚类为江苏兴化、江苏宿迁,第二亚类为江苏吴江、湖北大悟县。
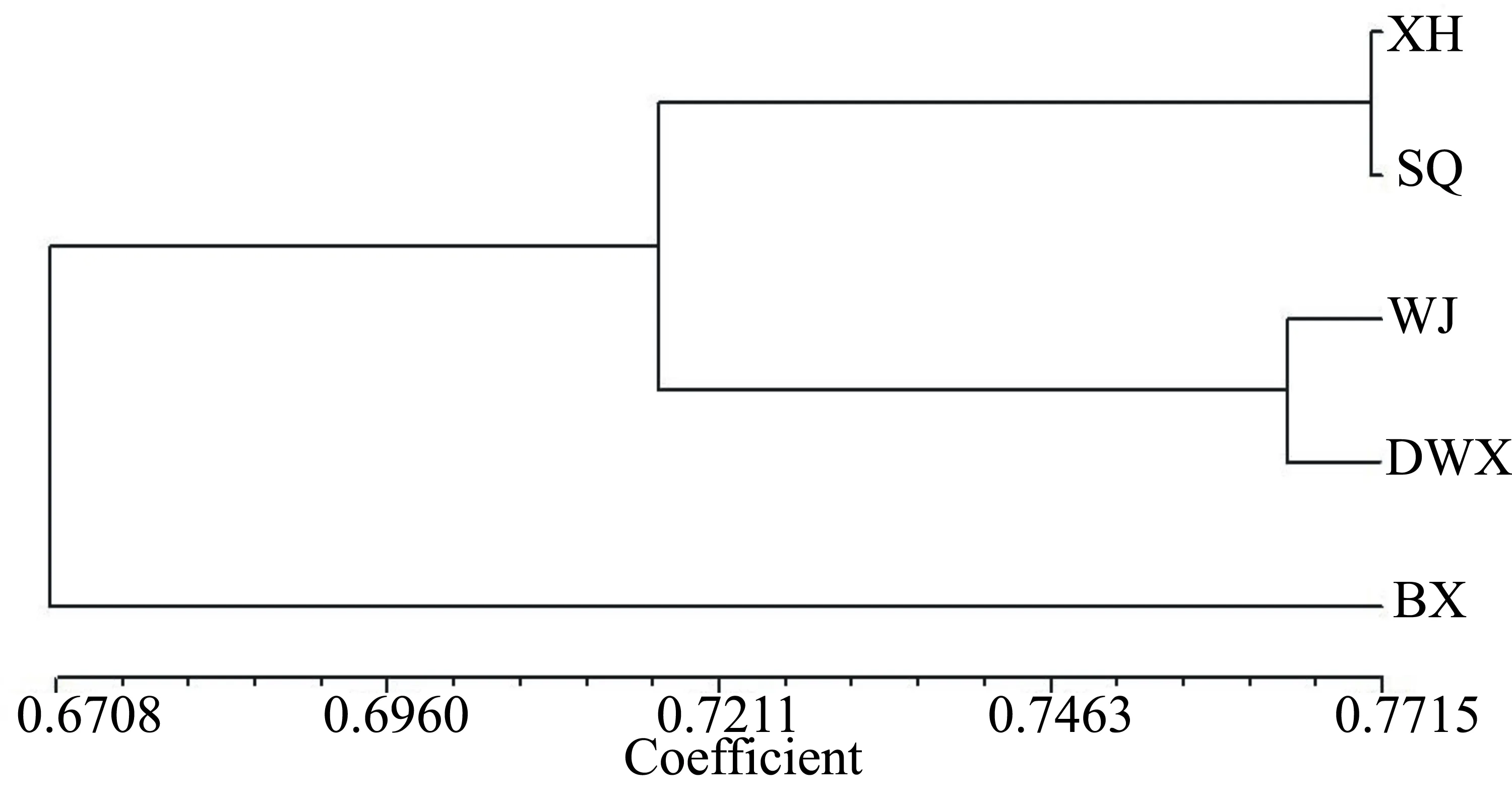
图3 UPGMA聚类分析法构建的不同群体宽体金线蛭的谱系关系
2.4 不同地理种群生长指标
仔蛭生长指标:由表4可见,黑龙江宾县仔蛭的平均体质量从11日龄开始显著低于(P<0.05)其它4个地区的平均体质量,生长较慢;江苏宿迁仔蛭11日龄的平均体质量显著低于(P<0.05)江苏吴江地区的11日龄平均体质量。不同地理种群仔蛭前16 d的平均日增重均较平稳,从16 d到21 d的平均日增重均出现较前16 d的3~5倍增长。不同地理种群仔蛭的相对增重率和特定增重率均在6 d时较高,11 d和16 d依次下降,并在16 d时达到最低值,之后迅速增长,21 d时再次到达一个峰值(见图4)。
幼蛭生长指标:由表5可见,黑龙江宾县幼蛭平均体质量从31 d就出现了显著低于(P<0.05)其他地区幼蛭平均体质量,其生长速率较慢。所有地区的幼蛭在1 d到61 d和91 d到121 d 期间的平均日增重基本差别不大,但61 d到91 d期间的平均日增重出现了零增长或极少量增长。相对增重率和特定增重率均呈现在31 d时最高,之后迅速下降,到91 d时达最低值,然后再呈增长趋势(见图5)。

表4 不同地理种群宽体金线蛭仔蛭不同日龄生长性状比较
注:同列数字字母相同表示差异不显著(P>0.05);字母不同表示差异显著(P<0.05)。下表同。

表5 不同地理种群宽体金线蛭幼蛭不同日龄生长性状比较
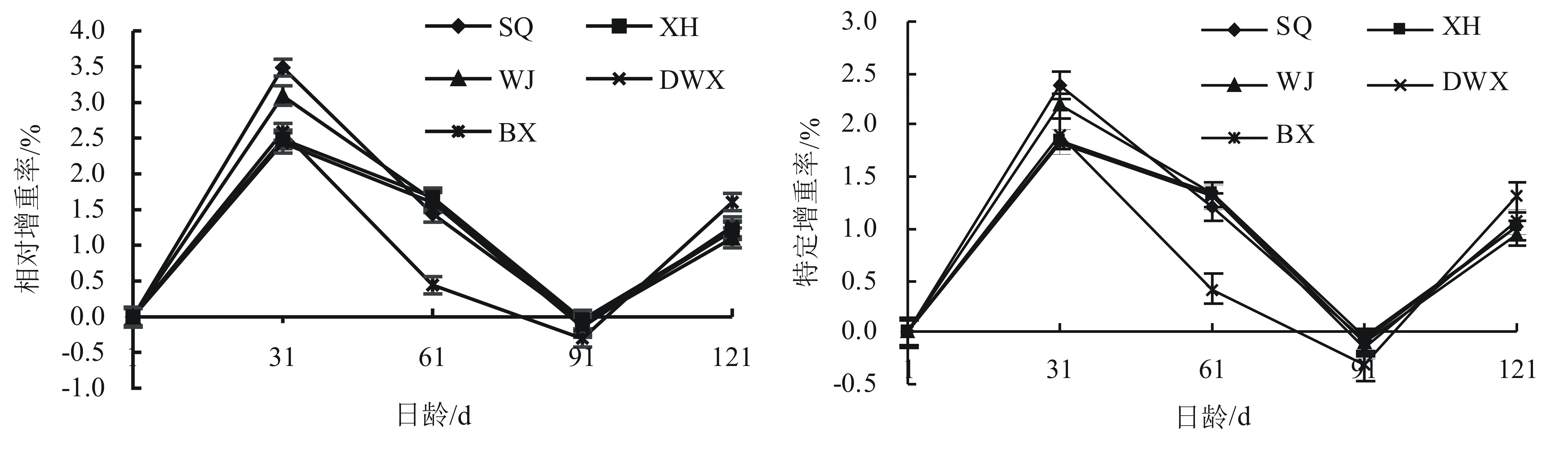
图5 不同地理种群宽体金线蛭幼蛭121日龄生长曲线
3 讨论
3.1 水蛭群体的遗传多样性
本研究的5个地理种群均为野生群体。结果表明,从遗传相似系数来看江苏和湖北群体间的遗传差距较小,其中江苏境内宿迁和兴化两个群体遗传差距最小,遗传相似系数为0.7707,江苏太湖和湖北大悟县群体次之,遗传相似系数为0.7643,江苏兴化和黑龙江宾县群体间的遗传相似系数最小,为0.6433。聚类分析显示,江苏地区3个群体和湖北大悟县群体聚为第一大类,黑龙江宾县群体聚为第二大类,其中第一大类中江苏兴化和江苏宿迁群体聚为第一亚类,江苏吴江和湖北大悟县群体聚为第二亚类。可见,遗传距离与实际的地理距离是有一定的相关性的。
梁超等[17]、蒋速飞等[18]、杨慧荣[19]等用RAPD技术分别研究了魁蚶(Scaphareabroughtonii)、日本沼虾(Macmbrachiumnipponensis)、黑鲷(Sparusmacrocephalus)不同地理群体的遗传变异,群体间的遗传相似系数为0.842~0.994。相比之下,宽体金线蛭不同地理群体的遗传相似系数值较低,相互之间的遗传差异较大。
3.2 不同地理种群生长指标
宽体金线蛭的生长速度主要与环境温度和适口饵料相关。生长试验中,不同地理种群仔蛭的生长温度控制在其最适宜生长温度25 ℃左右,饵料以浮游生物和适口田螺为主,5个地理种群21 d成活率均高达90%以上,说明此环境非常适合宽体金线蛭幼蛭生长,这一结果与王宣朋等[20]、熊良伟等[21]的研究基本一致。
在幼蛭的成活率和生长试验中,前61 d各地区幼蛭成活数量均很高,基本没有死亡,但在61~91 d期间死亡数量急剧上升,91 d时黑龙江宾县和湖北大悟县幼蛭仅剩53条、65条存活,江苏3个地区宽体金线蛭成活数量也仅剩85~87条。当水温从20 ℃升到27 ℃,宽体金线蛭生长正常,成活数量很高,但是当气温达到36 ℃,池塘水温在32 ℃左右,高温一直持续10 d左右时,各地区幼蛭均出现了停止摄食甚至死亡的现象,之后开始向池塘内24 h不间断加注新鲜井水,使池塘水温控制在27 ℃左右,各地区幼蛭死亡得到控制,这一结果进一步验证宽体金线蛭生长对温度条件的要求。本试验中,121 d时江苏省3个地区幼蛭成活率为70%左右,明显高于湖北大悟县(53.33%)和黑龙江宾县(44.17%),这可能跟原生长环境有关。王安纲指出,在其他条件不变的情况下,固着、隐蔽栖息和食物是构成宽体金线蛭分布的三大要素[22]。首先,江苏省3个地区幼蛭均采自湖泊,地区间气候差别不大,而湖北大悟县和黑龙江宾县的野生宽体金线蛭分别采自山溪、松花江等河流水域,常年水温较低,这使得后者繁殖时间较前者晚,或者繁殖温度较低,因此在江苏省宿迁市的养殖不利于其繁殖习性;其次,江苏省3个地区宽体金线蛭食物组成相似,但与湖北大悟县和黑龙江宾县相比,食物组成差别较大,其对宽体金线蛭的发育情况也会产生一定的影响;再次,江苏省湖泊与湖北和黑龙江河流中宽体金线蛭固着、隐蔽栖息的介质不同,其对宽体金线蛭生长也存在一定的影响。
黑龙江宾县的仔蛭和幼蛭平均体质量均表现出了显著低于(P<0.05)其他地区幼蛭平均体质量,说明黑龙江宾县地区的宽体金线蛭确实生长速度较慢。
综上,基于本研究结果,目前江苏3个地区的宽体金线蛭生长速度较快,是比较适合本地区养殖的,在宽体金线蛭的养殖购种时,最好选择与本地环境相近的亲本和蛭苗。结合RAPD研究结果,江苏地区的品种遗传距离较小,鉴于其生长较快的性状,在今后的水蛭育种工作中可以对其抗病性等相关性状进行种质改良。
