3种免疫增强剂对异育银鲫非特异性免疫功能的影响
2018-10-15吴亚锋陈静倪金娣刘迅猛袁锐王晶晶方苹
吴亚锋,陈静,倪金娣,刘迅猛,袁锐,王晶晶,方苹
(江苏省水生动物疫病预防控制中心,江苏 南京 210036)
异育银鲫(Carassius auratus gibelio)以其生长快、个体大、抗逆性强等特点在江苏地区大量养殖,2017年江苏年产量约60万t,占全国总产量近25%。随着渔业养殖的规模化和集约化发展,各种鱼病的暴发成了影响渔业养殖的重要制约因素。大量使用的抗生素和化学药品不仅在水产品中残留,危害人类健康,也污染水环境[1-2]。因此,研制饲料免疫增强剂,促进鱼体的疾病抵抗能力,对人工养殖异育银鲫技术的发展具有极其重要的意义。
鱼类免疫增强剂主要通过增强非特异性免疫应答而发挥作用,如促进补体、溶菌酶、蛋白酶抑制剂、反应蛋白、天然溶血素、凝聚素、巨球蛋白、巨噬细胞活化因子和干扰素等的合成,活化巨噬细胞、嗜中性粒细胞、非特异性细胞毒性细胞的吞噬杀菌功能[3]。20世纪80年代以来,国内外学者发现多种天然和人工合成的物质可以促进水产动物非特异性免疫应答,从而增强其抗病力。Cárdenas等[4]研究认为葡聚糖能够刺激太平洋红鲷鱼头肾白细胞的免疫机制,激活具有免疫的特性细胞,增强对嗜水气单胞菌的抗侵袭能力。李超等[5]将黄芪、大蒜、菊花、山楂和板蓝根五味中草药按等比的水提取物制成中草药免疫增强剂,投喂后能显著增强草鱼生长性能和免疫功能。沈锦玉等[6]指出壳聚糖能提高中华绒螯蟹非特异性免疫功能,对嗜水气单胞菌免疫保护率高达75%,较好地预防病害的发生。
为进一步了解不同免疫增强剂在水产养殖病害防治上的应用前景,该试验在异育银鲫的饲料中添加3种免疫增强剂,分析异育银鲫血清中的两种非特异性免疫指标,研究添加剂能否增强鲫鱼的免疫功能,从而增强抵抗疾病的能力,为安全有效预防病害提供科学依据,为开发防病绿色饲料添加剂奠定基础。
1 材料与方法
1.1 试验用鱼及饲养管理
试验用异育银鲫,购于江苏盐城市大丰区林松养殖场,80日龄左右,体长5~6 cm,平均体质量(23.94±0.84)g。试验开始前用基础饲料驯养14 d,以适应环境和试验饲料。试验开始后,每日于8:00、17:00投喂2次,投饲量为鱼体质量的3%,并根据生长、摄食情况作调整。每天于18:00换水1次,每次换水三分之一。水族缸规格为:80 cm×60 cm×120 cm,24 h不间断充气。静水养殖期间,水温为25℃左右,pH值为7.1左右,溶解氧约为7.3 mg/L,养殖时间为30 d。
1.2 试验药饵饲料制备
基础饲料是“通威191高蛋白鲫鱼育成配合型饲料”,粒径为1.5 mm,配方表见表1。在基础饲料中分别添加不同比例的酵母多糖、壳聚糖和维生素K,共6个试验组,一个对照组,每组设3个重复,各组功能饲料的成分组成见表2。
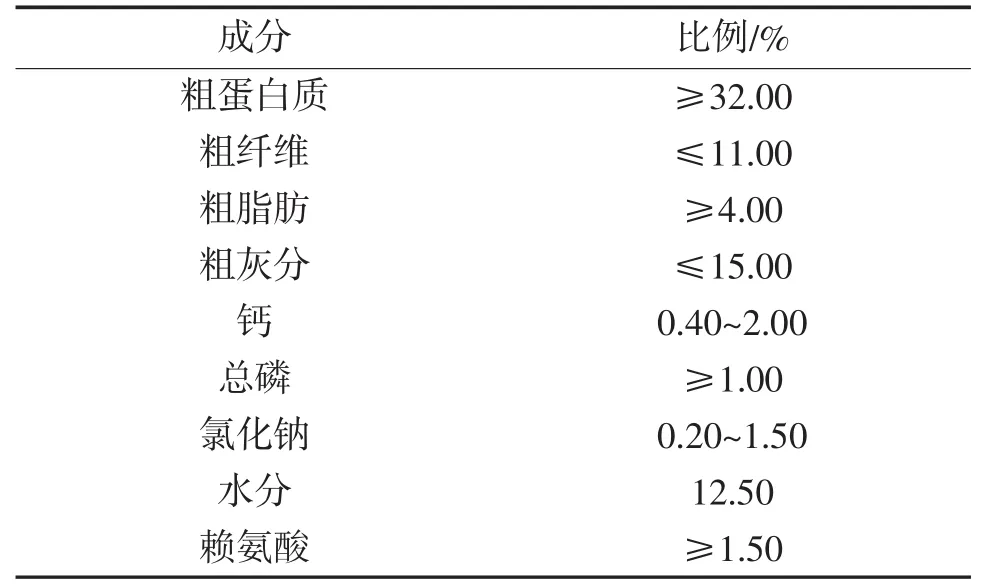
表1 基础饲料的营养指标
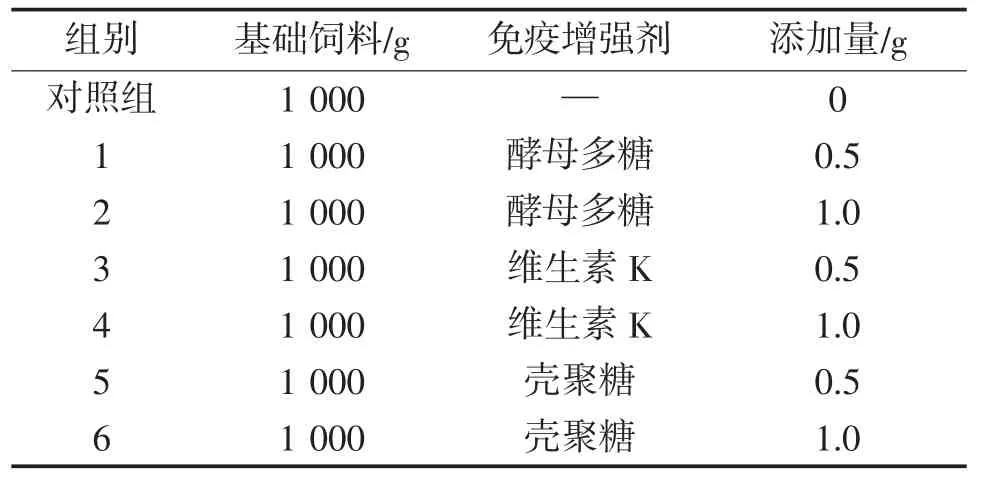
表2 各组功能饲料的成分组成
1.3 试验设计
将1 050尾暂养规格相同的健康异育银鲫随机分成7组,每组3个重复,每个重复50尾。先用基础饲料驯养,再用试验功能饲料投喂。每个重复分别在用功能饲料投喂的第 0、4、7、10、15、20、30 天随机取3尾供试鱼,测定非特异性免疫指标,测完免疫指标后的鱼不再用于该试验。
1.4 样品的采集与测定
从尾静脉无菌采血1 mL,不加抗凝剂,在37℃静置2 h后,3 000 r/min离心10 min取上层血清,在超低温冰箱(-80℃)中保存备用。使用南京建成生物工程研究所的试剂盒和酶标仪测定各样品中溶菌酶(LSZ)和超氧化物歧化酶(SOD)的活力。血清中SOD酶活性单位定义为每毫升血清中SOD抑制率达到50%所对应的SOD量为1个酶活性单位。
1.5 免疫保护率
从每个重复剩下的鱼中随机取10尾鱼测定免疫保护率。攻毒菌株为嗜水气单胞菌(Aeromonas hydrophila)JYZ170428GSU3,在预试验中根据Bliss法计算对异育银鲫半数致死量为1.37×106cfu/mL。每尾试验鱼腹腔注射0.2 mL,6.85×106cfu/mL嗜水气单胞菌新鲜菌液,对照组注射相同剂量的生理盐水,连续观察7 d,统计病死率,按照以下公式计算免疫保护率:免疫保护率(%)=[1-(受免鱼死亡尾数/对照鱼死亡尾数)]×100。
1.6 数据处理
数据以平均值±标准差(X±SD)表示,用SPSS 17.0软件进行统计分析。
2 结果
2.1 3种免疫增强剂对异育银鲫血清溶菌酶(LSZ)活性的影响
由表3可知,维生素K试验组对异育银鲫血清溶菌酶活性影响最大,第4天就显著提高(P<0.05),低剂量和高剂量组LSZ活性峰值分别为34.52、35.05 U/mL,为各试验组最高;壳聚糖组和合酵母多糖组效果略差,与对照组相比,到第7天LSZ活性才有显著性差异(P<0.05),且峰值略低。
从时间进展来看,各试验组LSZ活性在第0天时与对照组无显著性差异(P>0.05),在第7天时各试验组LSZ活性均显著高于对照组(P<0.05),在第10天,酵母多糖高剂量组和维生素K高剂量组达到峰值,随后下降;在第20天,免疫增强剂刺激效应减弱,除酵母多糖高剂量组外,其余试验组与对照组无显著性差异(P>0.05)。
2.2 3种免疫增强剂对异育银鲫血清超氧化物歧化酶(SOD)活力的影响
由表4可知,酵母多糖和壳聚糖对异育银鲫血清超氧化物歧化酶(SOD)活力影响较大,从第4天至第30天,与对照组相比均差异显著(P<0.05)。维生素K对SOD活力影响较小,与对照组相比,到第7天才有显著性差异(P<0.05),且峰值远远低于多糖组。

表3 3种免疫增强剂对异育银鲫血清溶菌酶含量的影响U/mL
从时间进展分析,各个试验组总体先上升再下降的趋势,但在第30天依然差异显著。各试验组SOD活性在第0天时与对照组无显著性差异(P>0.05);在第4天,酵母多糖和壳聚糖组与对照组相比差异显著(P<0.05);第7天至第30天,各试验组SOD活性均显著高于对照组(P<0.05);壳聚糖组在第10天SOD活力达到峰值,而酵母多糖组、维生素K组在第15天SOD活力达到峰值。3种免疫增强剂对血清SOD活力影响较大,低剂量组也能很好的免疫效果,除酵母多糖组的第10、15天,以及壳聚糖组的第4、7天外,其余时间点,各增强剂低剂量和高剂量组差异不显著(P>0.05)。
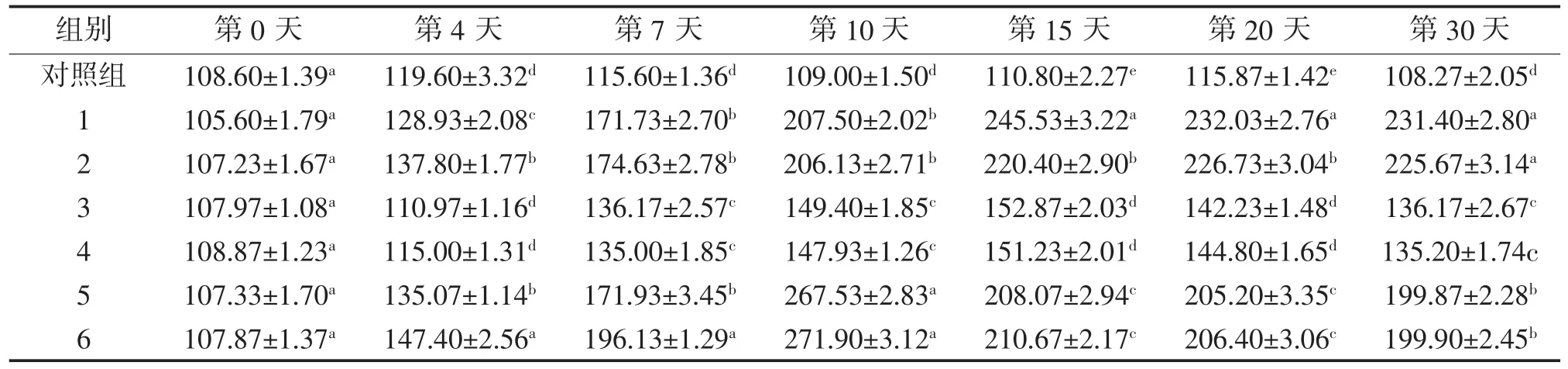
表4 3种免疫增强剂对异育银鲫超氧化物歧化酶含量的影响 U/mL
2.3 三种免疫增强剂对异育银鲫免疫保护率
由表5可知,3种免疫增强剂中,对异育银鲫免疫保护率最高的是高剂量壳聚糖组,为67.5%,最低的是高剂量维生素K组,为12.5%。从总体效果来讲,相同剂量下免疫保护率的顺序为:壳聚糖>酵母多糖>维生素K。
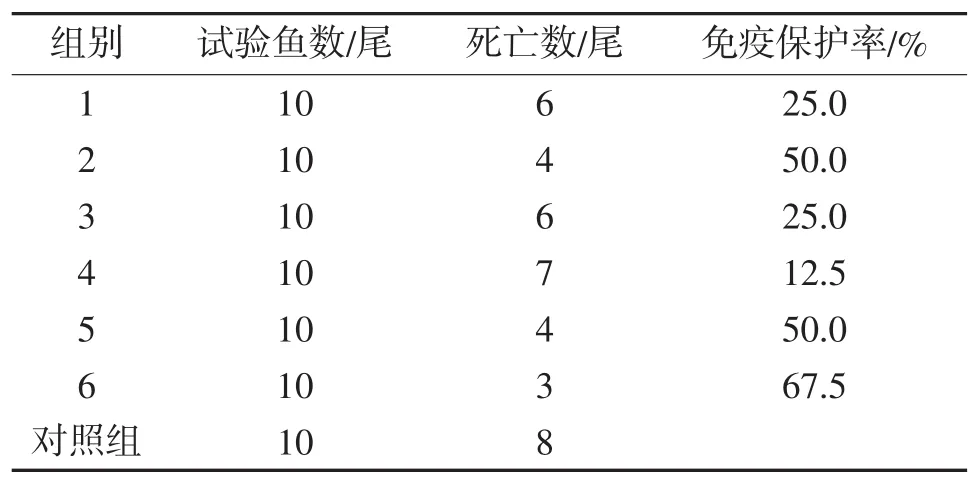
表5 3种免疫增强剂对异育银鲫的免疫保护率
3 讨论
3.1 3种免疫增强剂对异育银鲫非特异性免疫功能的影响
该试验发现在基础饲料中不同添加量的免疫增强剂均能提高异育银鲫的非特异性免疫能力,表现为血清溶菌酶(LSZ)活力和血清超氧化物岐化酶(SOD)活力均显著高于对照组。
酵母细胞壁多糖是从酵母细胞壁中分离出来的一些大分子多糖物质,存在特殊的键结方式和分子氢键的,能诱导机体产生一系列的细胞免疫和体液免疫反应,增强机体非特异性免疫力[7]。该研究中,酵母多糖对异育银鲫血清中SOD活力影响较大而对LSZ活力影响较小,这可能与添加量有关。姚嘉赟等[8]研究酵母多糖、黄芪和参芪3种不同免疫增强对黄颡鱼免疫功能的影响,也发现类似的问题,与酵母多糖低剂量组相比,酵母多糖高剂量组对LSZ活性差异极显著(P<0.01),而对SOD的活力,二者差异并不显著。但与对照组相比,酵母多糖组对增强鱼类的非特异性免疫依然具有较好的作用,而且酵母多糖的效果要远远好于黄芪和参芪。刘洪梅等[9]研究表明基础饲料中添加酵母多糖后,鲤鱼的生长性能和免疫功能等相关指标都较对照组显著提高。
壳聚糖可以活化巨噬细胞和自然杀伤细胞来表达一些引起发炎反应的细胞因子,如白介素-1、集落刺激因子和γ干扰素等。Rusell等[10]研究虹鳟甲壳素结合蛋白的发现,甲壳素结合蛋白存在于虹鳟的鳃、脾脏、肝窦、肾间质肠、皮肤、鱼鳔以及白细胞内部。这种蛋白质与人和啮齿类的一种血浆凝集素属于同源蛋白,上面还有多种受体,通过一种与乙酰化的甲壳素或甲壳素片段高亲和性的钙依赖性途径参与到细胞内吞作用中。陈勇等[11]研究认为壳聚糖通过提高血清T3水平、降低血清皮质醇水平增强异育银鲫的非特异免疫功能。该研究中,壳聚糖能显著提高LSZ和SOD的活力,异育银鲫非特异性免疫能力显著提高。这与张涛[12]在饲料中添加了0.22%、0.44%、0.66%的酵母多糖后,草鱼血清溶菌酶(LSZ)活性均显著提高(P<0.05)相近。华雪铭等[13]也认为壳聚糖、益生菌、益生菌与壳聚糖混合物或甘露聚糖与益生菌混合物作为饲料添加剂具有调节鱼类免疫功能和免疫重建的作用。
在水产动物中,维生素K的研究主要集中在体内发挥的生理功能和对它的需求上[14],对于维生素K对水生动物非特异性免疫功能影响的研究报道比不多见。该研究发现饲料中添加一定剂量的维生素K,也能显著提高异育银鲫非特异性免疫能力。这与王洋等[15]认为适当浓度维生素K对SOD活性有激活效应相符合。但相同剂量下维生素K提高LSZ的能力显著高于多糖组,维生素K影响调控LSZ的机制也有待进一步研究。
3.2 不同添加量免疫增强剂对异育银鲫非特异性免疫功能的影响
水产养殖中,同种免疫增强剂添加剂量不同,其使用效果也存在一定的差异。该研究中,在第10天至第30天,酵母多糖低剂量组SOD活力的增长反而显著高于酵母多糖高剂量组;维生素K对血清中LSZ活力的影响也存在同样的现象。刘爱君等[16]用奥尼罗非鱼做试验,发现饲料中添加0.50%~0.75%甘露聚糖,均可提高营养物质消化率和机体免疫力,但添加剂量为0.50%时最佳。Siwicki等[17]研究了口服不同剂量的左旋咪唑对虹鳟抗出血病的作用,结果显示添加量10~20 mg/Kg,有良好的免疫保护力;当剂量上升到50 mg/Kg时,对机体的免疫系统产生抑制现象。这可能是因为机体免疫系统对免疫增强剂刺激的代偿性反应。
近年来,随着人们对食品安全问题的日益重视,免疫增强剂以毒副作用小,易分解,在环境和动物体内无残留,既能提高水产动物生长性能,又能提高水产动物的免疫力和抗病力的优点,可以作为预防水产疾病暴发的好产品,在水产养殖中将越来越受欢迎。目前,多种复合型免疫增强剂的研究已经成为热点和趋势[18-20]。该研究证实了酵母多糖和壳聚糖对异育银鲫非特异性免疫的增强作用,探究了维生素K与鱼类免疫系统的相互关系,为进一步研发新型复合型免疫增强剂提供科学依据。这3种免疫增强剂具有廉价易得,且性质稳定易储存,值得在今后养殖生产中加以大力推广。
