西藏桃儿七结实特性及其与主要生境因子的关系
2018-10-11郭其强张艳福汪书丽
郭其强, 丹 曲, 张艳福, 汪书丽, 池 翔
(1.贵州大学/贵州省森林资源与环境研究中心,贵州贵阳 550025;2.西藏高原生态研究所,西藏林芝 860000
桃儿七(Sinopodophyllumhexandrum)为小檗科(Berberidaceae)桃儿七属(Sinopodophyllum)的多年生草本植物,别称为西藏鬼臼,为国家三级保护植物。植株高20~50 cm,多须根,直立茎上着生叶片2张,叶基部心形,有3~5个深裂,边缘为粗锯齿,主要分布在喜马拉雅海拔2 700~4 500 m 高山峡谷区的林下、林缘或灌丛附近[1]。桃儿七主治祛风除湿、活血通经和抑制肿瘤等疾病[2],主要以根、茎入药,使得野生资源被掠夺式采挖,遭受严重破坏,导致野生种群数量日益稀少[3]。加之藏药以果实入药,极大地影响了种群的天然繁殖能力,现处于严重濒危状态,因此被誉为“人为濒危种”[4],也被列入《中国植物红皮书》和《中国珍稀濒危植物名录》[5-6]。
桃儿七种子承担着种群有性生殖和分布区扩张的重要使命,也是其度过不良外界环境的重要载体[7]。植物果实生长、种子质量与种群生境密切相关,开展桃儿七结实特性及其与生境因子的关系研究,对于揭示野生状况下种群对环境的适应性和有性生殖规律具有重要意义[8]。近年来,学者们针对桃儿七繁殖特性[7,9]、药用价值[2,10-11]、组织培养[12-13]、栽培技术[14]和遗传变异[15-17]等方面的研究已有大量报道,这些成果对于深入认识野生资源的种质资源特性和促进人工繁育具有重要的促进作用,然而这些报道仍未能解释高寒生境下桃儿七种群的结实规律及其与生境因子间的作用关系。鉴于此,本研究拟通过对西藏东南部林芝、山南和昌都地区野生桃儿七种群分布较为集中的10个县域进行种实发育过程监测和数量调查,结合当地的气候与小生境条件,综合分析西藏桃儿七种群果实生长周期、果子大小和种子质量等特性,及其与主要生境因子的相关性,为采取合理措施促进种群天然恢复、指导人工育种和栽培提供科学依据。
1 研究区概况与方法
1.1 研究区概况
本研究位于西藏东南部的10个县域为研究区域,地理位置为28° 47′ 31″ ~31° 35′ 17″N和92° 25′ 25″ ~ 98° 22′ 38″E,总面积约为22.7万km2,也是西藏森林和高山灌丛的主要分布区。研究区地处青藏高原东南部的高山峡谷区,分布海拔为2 715~4 354 m,年平均气温为4.5~12.4 ℃。干、雨季分明,年均降水量为478~1 634 mm,且多集中在夏季(7—9月占年总量的80%左右)。土壤主要为森林褐土和高山草原土。桃儿七种群野生种群分布区主要为林下、林缘和灌丛下,不同研究地点位置、主要气候状况和小生境见表1。
2 研究方法
2.1 样地设置与监测
在桃儿七分布相对集中的区域各设置3个 5 m×5 m的典型样地38块,开展样地调查和结实特性的监测。具体内容为:(1)记录样地概况,包括经纬度、海拔高度、地形、小生境和土壤类型等;(2)由于研究区范围大,准确监测生境地温度和降水较为困难,本研究根据当地县气象局监测的数据代表研究区的年均温度和年降水量;(3)对样地内的每一个桃儿七植株进行编号,对其果实生长定期观察,从植株坐果开始记录,直到果实成熟(一半以上变红)视为果实生长期,并统计成果率(即坐果后发育为成熟果实个体的比率),其间每10 d观察1次;(4)果子成熟后,测定单果鲜质量、果长和果宽,晾干后剥开数种子数,全部称质量后计算千粒质量。样地设置统一在2015年4—5月间完成,待9—10月果实成熟后,测定鲜果性状,后带回实验室测定种子数和千粒质量,2016年重复该实验。以2年试验结果的平均值作为本研究的基础数据。鉴于研究区不同种群距离较远,研究组人员无法全面覆盖监测数据,因此在10个种群中,林芝县(LZD)、米林县(MLZ)和工布江达县(GBS)为研究者实地监测获取数据,其余7个种群聘请了当地护林员进行监测和记录数据,果实成熟后迅速带回西藏农牧学院,在实验室测定相关参数。
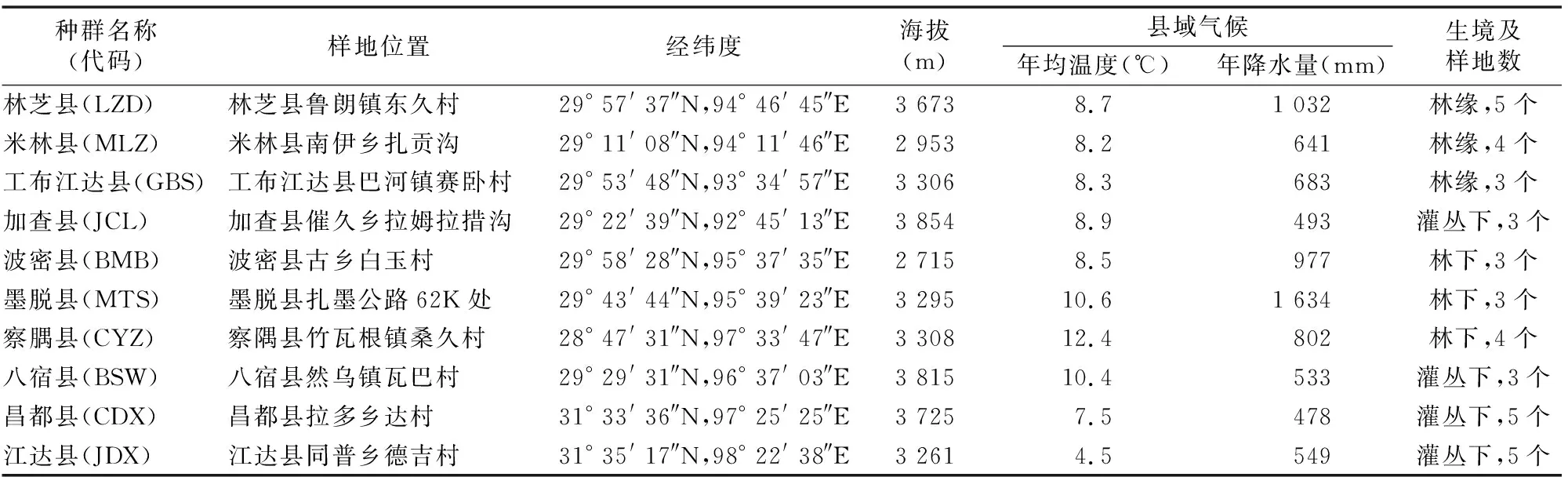
表1 西藏不同桃儿七种群位样地置及气候条件
2.2 数据处理
以不同研究区种群样地中全部果实特征参数和种子指标的平均值为统计结果;用SPSS 18.0软件比较同参数不同种群间的差异性,分析果实生长周期、果实和种子性状与主要生境因子的相关性。
2 结果与分析
2.1 果实发育特性
监测不同种群桃儿七坐果时间和结果数,直到果实成熟的时间和果实数量,以此统计果实生长时间和成果率。由表2可知,西藏桃儿七坐果时间最早和最晚时间分别为4月上旬和5月下旬,成熟时间最早和最晚分别为9月上旬和10月中旬。种群MLZ果实生长时间最长,约为153 d,而种群JCL和MTS生长时间最短,约为112 d。结果数量最多的为种群GBS,约为25.32 个/100 m2,最少的为种群MTS,仅为 10.42 个/100 m2。坐果后能生长到成熟的比率(即成果率)以种群MLZ为最高,为84.23%,最低为MTS,仅为57.72%。
2.2 果实与种子性状
桃儿七果实为卵圆形的浆果,种子多数为果皮所包被。由表3可知,西藏不同桃儿七果实长度在种群MLZ与LZD、GBS、CYZ、CDX、JDX、JCL、BMB、MTS、BSW间均存在显著差异;果宽在种群LZD、MLZ、GBS、CDX与JCL、CYZ、BSW、JDX、BMB、MTS间均存在显著差异;长宽比在种群MLZ、GBS、CYZ与LZD、JCL、BMB、CDX、JDX、MTS、BSW间均存在显著差异;单果鲜质量则在种群MLZ、MTS 、CDX、JDX与种群LZD、GBS、BMB、CYZ、JCL间均存在显著差异。上述特征具体表现为,种群MLZ和CDX等果实长度和宽度均较大,其长宽比大,外形更趋于纺锤形,单果鲜质量值较大;而种群MTS和BSW中果实相对较小,其长宽比较小,外形更趋于圆形,其单果鲜质量均较小(种群MTS除外);种群LZD、BMB、MTS与MLZ、GBS、CDX、JCL、CYZ 、BSW、JDX的单果种子数均存在显著差异;种群MLZ与GBS、JCL、CDX、JDX、LZD、CYZ、BSW、BMB、MTS的种子千粒质量间均存在显著差异。上述特性具体表现为种群MLZ、GBS和CDX单果中种子数较多,且千粒质量也较大,而种群BMB和MTS单果种子数和千粒质量均较低。
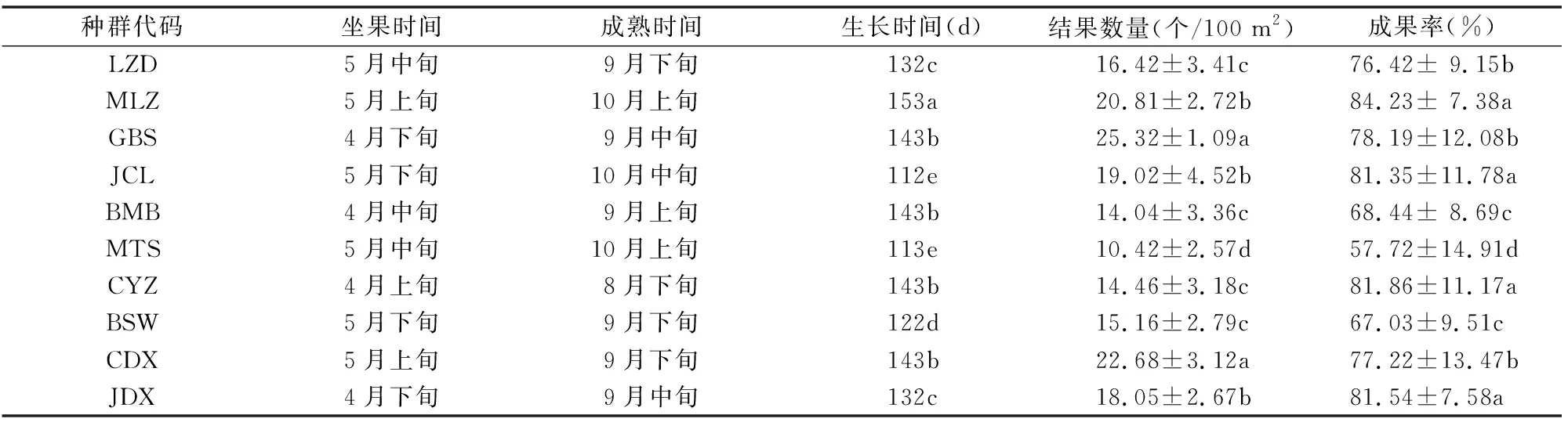
表2 西藏不同桃儿七种群果实生长时间及成果率
注:同列数据后不同小写字母表示在不同种群间存在显著差异(P<0.05)。下表同。
2.3 果实、种子特性与生境因子的相关性分析
不同桃儿七种群果实生长时间、果实大小和种子特性各参数间存在显著相关性,同时也受各生境因子的影响。相关分析表明:(1)海拔因子与桃儿七果实生长时间存在显著负相关,年降水量与结果数量、成果率、单果种子数和千粒质量间均存在显著负相关,其相关度的排序依次为千粒质量>成果率>结果数>单果种子数,而年均温度与果实和种子各参数间均无显著相关性;(2)果实生长时间与果实长度和宽度间存在显著正相关性;(3)结果数量不但与成果率、果长、果宽和单果种子数间均存在显著正相关性,还与千粒质量间存在极显著正相关性,其相关度排序依次为千粒质量>单果种子数>果宽>成果率>果长;(4)成果率不但与果长存在极显著正相关性,还与果宽、长宽比和千粒质量间存在显著正相关性,其相关度排序依次为果长>千粒质量>果宽>长宽比;(5)单果质量与单果种子数呈显著正相关,且果长与果宽、长宽比,千粒质量与单果种子数间均存在极显著正相关性(表4)。
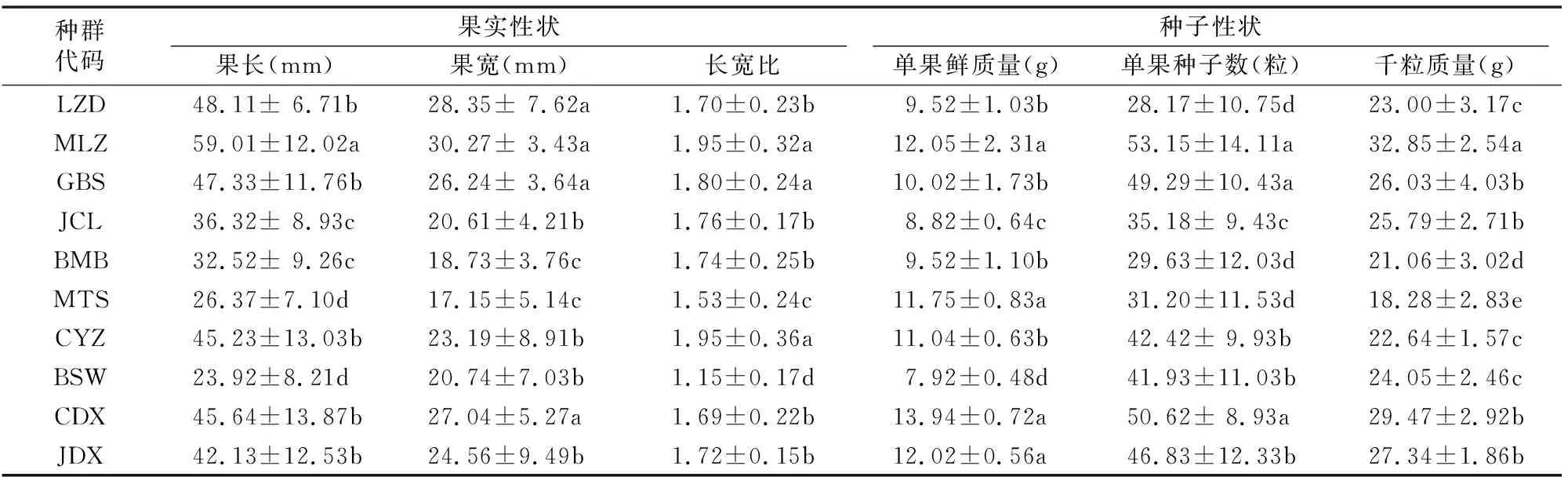
表3 西藏不同桃儿七种群果实与种子性状参数

表4 西藏不同桃儿七种群果实、种子特性与生境因子的相关性
注:*、**分别表示在0.05、0.01水平显著相关。
3 结论与讨论
西藏桃儿七种群生长在不连续分布的高寒生境区,分布区不同的气候与生境条件导致种群果实生长时间、个体大小和形状,以及种子数量和质量差异显著。由本研究可知,果实特性表现为:生长周期集中在每年4月上旬至10月中旬,即112~153 d,平均结果数量为10.42~25.32个/100 m2,成果率为57.72%~84.23%,果实长、宽度分别为23.92~59.01、17.15~30.27 mm,因长宽比的值不同,其外形为纺锤形至近圆形,单果鲜质量为7.92~13.94 g。种子特性表现为:单果种子数28.17~53.15粒,千粒质量18.28~32.85 g。果实个体发育结果和种子特性不但受海拔和当地主要气候因子的影响,而且与自身特性密切相关。
桃儿七球果和种子特性不但与自身的生物学特性密切相关[18],而且受生境条件的影响[19]。本研究发现,海拔越高,桃儿七果实生长时间越短,而降水量增加,其结果数量和质量趋于降低。从植物生理学和生态学角度分析,随海拔升高,温度趋于降低,造成春季土壤升温较晚,使得宿根植物的萌发期推迟,其开花时间必然较低海拔晚,而秋天来临时高海拔区降温较早,植物会加快果实成熟,尽快形成能够成熟的种子,以维持天然种群的存在。夏季大量的降水会降低到达植物叶表面的光合有效辐射强度和持续时间[20],造成植物光合作用效率和养分吸收能力的下降,导致植物体营养生长和生殖生长变慢,并影响果实膨大和种子发育,最终导致桃儿七结果数量和质量降低。马绍宾等研究报道,云南迪庆桃儿七(海拔 3 300~3 600 m)果实生长期为45~120 d,平均单果种子数约为60 Ind.,单果质量与单果种子数呈线性正相关[9]。与其相比,本研究中桃儿七果实生长期更长,但单果种子数却较低。主要是因为西藏桃儿七生长在高海拔的高山峡谷区,较低的年均温度使得果实成熟延迟,生长期变长;此外,西藏桃儿七在4—5月开花,后靠自花传粉完成受精作用[4],而此时西藏亚高山区仍有少量降雪[21],一旦积雪覆盖花朵就必然降低授粉概率,同时低温造成的凝冻也会降低花粉的活力[22-23],由此导致单果种子数减少;单果质量与种子数的关系与本研究结论相同。
桃儿七生殖生长特性与生境条件有关[9,24]。本研究发现,西藏桃儿七种群不同,其结果数越多,果实个体越大,成果率和单果千粒质量也越高,该结论可能与不同种群分布区的生境条件和养分供给状况有关。以往研究发现,桃儿七为半阴生植物,在林缘、林窗和灌木林下生长较好,且喜欢肥沃土壤[1,9]。本研究显示,种群MLZ、GBS和CDX均位于林缘或灌丛下,土壤相对肥沃,光照条件适中,为种群的生殖生长提供了较为有利的条件,因而结果数和种子质量均较高。而种群BMB和MTS位于原始乔木林下,上层林冠的遮阴使得林下光照条件较差,影响叶片的光合碳同化效率,使得生殖生长效率低下。因此,林缘和灌丛下的生境条件对桃儿七的果实发育较为有利。
未来如何采取有效措施促进野生桃儿七种群恢复和满足市场对资源的需求,是当前亟待解决的问题。由于西藏桃儿七以有性生殖为主,在优良种源区应加强对野生植株的保护并减少人为采种和外界干扰,以维持种群生境的相对稳定性,促进种群天然更新。同时,在结实量较大的年份可适当开展人工采种,大力发展人工育苗和高效栽培,以弥补野生资源不足和市场需求量大的矛盾,最终实现西藏桃儿七资源的合理开发和可持续利用。
