秦王川湿地滨藜叶脉性状与蒸腾速率关系对种群密度的响应
2018-09-28赵成章李雪萍
任 悦,赵成章,李雪萍,张 晶,雷 蕾
西北师范大学地理与环境科学学院,甘肃省湿地资源保护与产业发展工程研究中心,兰州 730070
蒸腾速率直接决定了植物体内的水分平衡[1],影响植物体与外界环境间的物质循环效率[2]。叶片形态能够反映植物适应特定生境的光合生理生态特性,在植物与环境协同进化过程中,逐步形成了特定的叶脉性状[3],并培育了物质传输、叶片保护、光能截获、水分供给平衡等方面的支持功能,为植物应对各种胁迫和干扰,正常开展生长和繁殖活动提供了保障[4]。面对资源异质性分布的生境和植物生活史中光合产物的供需矛盾,叶片各功能性状的自我调整与适应显得至关重要[5],叶脉网络功能性状具有物质运输、叶片支撑与保护的生理学和力学双重功能[6]。叶脉密度反映了叶脉与叶肉的接触程度和叶片的碳投资,与叶面积大小、叶片厚度共同影响着植物的蒸腾速率、净光合速率和水分利用效率,且能表征叶片抵御物理损伤的能力[7];中脉直径主要承担叶片支撑和水分传输功能,是叶片展开和增大植物光截获能力的重要基础条件,直径越大,表示叶脉系统的运输能力也越强[8]。植物叶片的蒸腾速率受制于叶脉形态及其功能的发挥[9],较高的叶脉密度能够供给蒸腾作用所需大量水分和能量消耗,有助于调控植物叶片温度和维持植物体与土壤、大气之间的物质循环[10- 11];中脉直径凭借较高的导水率和机械性能使得水分运输到叶片末端比叶边缘更容易,伴随着较强的支撑力,能够防止叶缘弯曲[12],进而有效调节植物的蒸腾速率,由此可知,叶脉密度和中脉直径构建的资源配置机制,是协调统一叶片光合作用中水分运输的有效性与安全性的重要基础[13],反映了叶片在环境胁迫下为满足植物的光合生理需求所形成的叶脉结构组合和水分运输策略[7,14- 15]。密度通过影响植物群落的光照、温度和土壤水分等环境条件,能够引起不同程度的邻体隐蔽和生存空间竞争[16],改变种群内部每个植株可获得性资源的数量,进而造成生境尺度的环境胁迫和干扰;为了创造合理的光截获面积,获取满足植物光合作用的基本光照条件,避免不必要的资源浪费,植株会调整叶脉密度与中脉直径的资源投入模式,从而维持植物蒸腾失水与土壤-植物-大气连续体有效供水之间的动态平衡。因此,研究湿地植物叶蒸腾速率和叶脉性状关系对密度制约的响应机制,对认识叶脉网络建成中碳投入机制与植物光合效率之间的协同变异关系具有重要的理论意义。
滨藜为苋科滨藜属的植物,是全世界干旱、半干旱地区的典型植物,具有适应性强、耐干旱、耐盐碱的特点,多生长于海滨、轻度盐碱湿地、草地和沙土地,也是西北荒漠区盐沼湿地的优势植物种群之一[17]。学术界围绕叶脉网络功能性状与光合作用的关系[18]、叶片蒸腾速率和叶片导水能力[19- 20]、叶脉密度与叶大小[9,21]、叶脉功能性状与叶厚度的权衡关系[4,22]、植物叶片光合参数与环境因子的关系[23]开展了广泛研究,有关滨藜的生物学特征、对盐分的适应性[17]也得到了重视,但密度制约下湿地植物叶脉性状生长关系与蒸腾速率的调节分配机理尚不清晰。鉴于此,本研究以兰州新区秦王川国家湿地公园盐沼湿地的滨藜为研究对象,通过分析不同种群密度下滨藜蒸腾速率与叶脉性状的关系,试图明晰:(1)不同种群密度滨藜蒸腾速率与叶脉性状存在何种关系?(2)形成这种生长关系的主要原因有哪些?旨在丰富对湿地植物叶脉性状建成的种群适应性策略的认识和理解。
1 材料和方法
1.1 研究区域概况
研究区位于甘肃省兰州市的秦王川国家湿地公园,地理坐标为36°27′59.6″N,103°39′5.6″E,海拔1895—1897 m,属大陆性季风气候,年平均气温6.9℃,年均降水量285 mm,主要集中在6—9月,年日照时间2700 h,平均无霜期126 d。由于地处乌鞘岭褶皱山岭南侧的边缘低山区,四周山地环绕,中心地带地势平坦开阔,形成盆地,盆地南部广泛分布第四系松散层孔隙潜水,含水层为砂碎石及中细砂层,受常年或季节性水淹的影响,形成以淡灰钙土为主的自然土壤,发育了水生和湿生植物群落,以湿生和挺水植物为主。主要植物有:滨藜、芦苇(Phragmitesaustralis)、枸杞(Lyciumchincnse)、柽柳(Tamarixchinensis)、香蒲(Typhaangustifolia)、灰绿藜(Chenopodiumglaucum)、紫菀(Astertataricus)、假苇拂子茅(Calamagrotispseudophragmites)、盐地碱蓬(Suaedasalsa)、苦苣菜(Sonchusoleraceus)、盐角草(Salicorniaeuropaea)、早熟禾(Poaannua)等。
1.2 实验设置
1.2.1 植物采样及测量
2016年9月,在实地考察的基础上,在湿地公园保育区选择一块长度约120 m、宽度约97 m、高度约2—3.1 m的台阶地,滨藜为单一优势种的盐沼湿地;随着地势的微弱抬升,滨藜的密度逐步增加、株丛直径明显增大,以“株”为取样单位进行滨藜密度调查,发现滨藜密度介于9—21株/m2之间。在台阶地的东西向坡面从坡底向坡顶间隔10 m依次布置3条宽20 m、长60 m的样带,根据滨藜种群密度的变化规律,在3条平行样带上从台地下缘向台地顶部依次设置3个10 m × 10 m的样地(I、II、III),分别为:1)高密度(I),位于台地的中心,滨藜植株密度为16—21株/m2,株丛直径和株高较大,土壤水分充足,光资源不足;2)中密度(II)位于台地中心至边缘地带,滨藜植株密度为9—15株/m2,株丛直径和株高居中,土壤水分及光资源适中;3)低密度(III)位于台地的下边缘,滨藜植株密度<9株/m2,株丛直径和株高较小,土壤水分不足,光资源充足。在每个样地随机设置3个2 m × 2 m样方,总计27个样方。然后在每一个样方随机选择6株滨藜(株高为大中小各2株),先用卷尺测量从基部到最高分枝的高度,定为株高,然后选取每个植株最外层四个方位充分伸展且健康完整的两个叶片,做好标记后进行以下步骤:
(1)植物光合参数测定。于9月2—9日,选择晴朗天气的9:00—12:00,利用GFS-3000便携式光合测量系统(Heinz Walz GmbH, Bavaria, Germany)进行每个标记叶片光合参数的测定。测量过程中使用人工红蓝光源,光合有效辐射(PAR)定为1200 μmol m-2s-1,CO2浓度约为340 μmol/mol,相对湿度(RH)保持在40%—50%,流速设定为750 μmol/s,每个叶片记录6组数据用于统计分析,分别测定叶片的Pn、Tr及VPD等参数[24],叶室面积为8 cm2,将不能充满叶室的叶片,保存于湿润的封口袋中,带回实验室,用便携式激光叶面积仪(CI-202, Walz, Camas, USA)测定叶面积,进而计算Pn和Tr。
(2)光合有效辐射(PAR)的测定。选择与光合参数测定同一天,即晴朗无云的天气,用手持光量子计(3415F, 3415F, Walz, Plainfield, USA)于9:00—11:00对3个样地内滨藜种群的上方、中层和地表处进行PAR测定。
(3)叶性状的测定。基于每个滨藜被标记的两个叶片,采集后保存于湿润的封口袋中,带回实验室用福尔马林-乙酸溶液(37%甲醛溶液,50%乙醇和13%冰醋酸溶液)固定。①每片叶子先测量叶片形态特征,将采集的叶片用干纱布擦干净,用便携式激光叶面积仪(CI-202, Walz, Camas, USA)对叶片进行扫描,测定叶面积;②叶脉密度的测量,用含5% NaOH的乙醇进行化学清理,再用番红-固绿染色[25],用水包埋呈透明薄膜状在体视显微镜(SMZ168-BL, Motic, Hong Kong, China)下放大10倍后拍照,每个叶片拍摄10个视野用于叶脉性状的观测,用Motic Images Plus 2.0软件获得各个照片中的中脉直径和叶脉的总长度,所有照片中得出的平均值即为该叶片的中脉直径和叶脉密度,叶脉密度用单位叶面积(单位: mm2)的叶脉总长度(单位: mm)表示[9]。最后将叶片装入信封,于80℃烘箱中烘干48 h,称量(精度0.0001 g)。比叶面积(SLA)用叶面积和叶干重的比值表示。
1.2.2 土壤取样与理化性质测量
(1)土壤含水量采样。土壤取样于2016年9月下旬完成,取样时间内基本无明显降水,土壤水盐状况相对稳定。在小样方内随机选取3个样点挖掘1 m × 1 m × 0.5 m土壤剖面,用环刀分5层间隔10 cm采取土样,现场编号、各土层均匀混合称鲜量,然后带回实验室在105℃的烘箱内烘12 h,取出称质量,最后计算出各样地的0—50 cm土层土壤质量含水量。
(2)土壤盐分采样。用电导法测定土壤含盐量,在室温下称取过2 mm筛的风干土样10 g,加50 mL去CO2蒸馏水(水土比为5∶1),取浸出液,置振荡机上振荡5 min。将布氏漏斗与抽气系统相连后把悬浊的土浆缓缓倒入漏斗,直至抽滤完毕,滤液倒入三角瓶备用。用上海雷磁仪器厂生产的DDS-11C便携式电导仪测浸出液的电导率,3次重复,取平均值。
1.3 数据处理
对不同盖度样地湿地群落生物学特征和滨藜种群的各功能性状的实验数据进行分析,对滨藜的蒸腾速率(Tr)、叶脉密度(VD)、中脉直径(MD)先进行以10为底的对数转换,使之符合正态分布后再进行分析。对蒸腾速率和叶脉密度以及蒸腾速率和中脉直径两组关系的研究,均采用y=ɑxb,线性转换成lgy=lgɑ+blgx,x和y表示两个特征参数,ɑ为性状关系的截距,b为斜率,即异速生长参数或相对生长的指数,当b=1时,表示两者是等速关系;当b>1时,表示y的增加程度大于x的增加程度;当b<1时,表示y的增加程度小于x的增加程度。主要采用标准化主轴估计(SMA)方法[26],由软件SMATR Version 2.0[27]计算完成,每一个回归斜率的置信区间根据[28]方法计算。对不同密度样地植物功能性状平均值的差异比较采用单因素方差分析(one-way ANOVA),显著性水平设为0.05。实验数据分析采用SPSS 16.0软件进行,用SigmaPlot 10.0和Excel软件绘图。
2 结果分析
2.1 不同湿地生境下滨藜群落的生物学特征和土壤特性
湿地生境中不同密度下滨藜群落的生物学特征、土壤理化性质和光合有效辐射存在显著差异(P<0.05, 表1),随着种群密度的逐渐降低,滨藜平均株高、盖度、植株密度及土壤含水量呈现减小的趋势,从高密度(I)到低密度(III)分别减小了83.87%、57.42%、85.7%、11%;PAR(为上中下三层均值)呈现增大的趋势,由高密度(I)的769.25 μmol m-2s-1增加到低密度(III)的1195.00 μmol m-2s-1,增加了0.55倍。
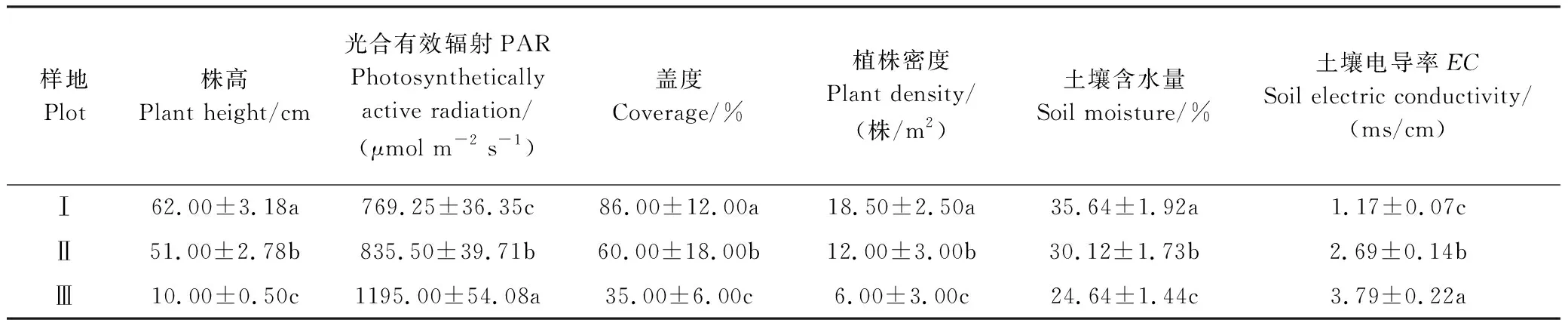
表1 不同湿地生境下滨藜群落的生物学特征和土壤特性(平均值±标准误差)
同列不同小写字母表示样地间差异显著(P<0.05),n=108。
2.2 不同密度下滨藜叶性状特征和光合特性
如表2所示,不同密度下滨藜叶性状和光合生理参数的变化趋势存在显著差异(P<0.05, 表2)。随着种群密度的逐渐降低,滨藜叶脉密度、叶干重、VPD、蒸腾速率(Tr)、Pn均呈显著增加趋势,从高密度(I)到低密度(III)分别增加了44.17%、23.72%、22.78%、52.46%、73.46%;中脉直径、叶面积、比叶面积、空气相对湿度(RH)均呈显著的减小趋势,从高密度(I)到低密度(III)分别减少了73.17%、34.91%、50.35%、12.8%;叶干重、VPD、蒸腾速率(Tr)在中密度(II)、低密度(III)均出现较大值且无显著差异(P>0.05),但低密度(III)大于中密度(II);叶面积、空气相对湿度(RH)在中密度(II)、低密度(III)均出现较小值且无显著差异(P>0.05),但低密度(III)小于中密度(II)。滨藜叶性状和光合特性在3个样地变化较显著,且叶性状具有较高的表型可塑性,可以较好的适应特殊生境条件。
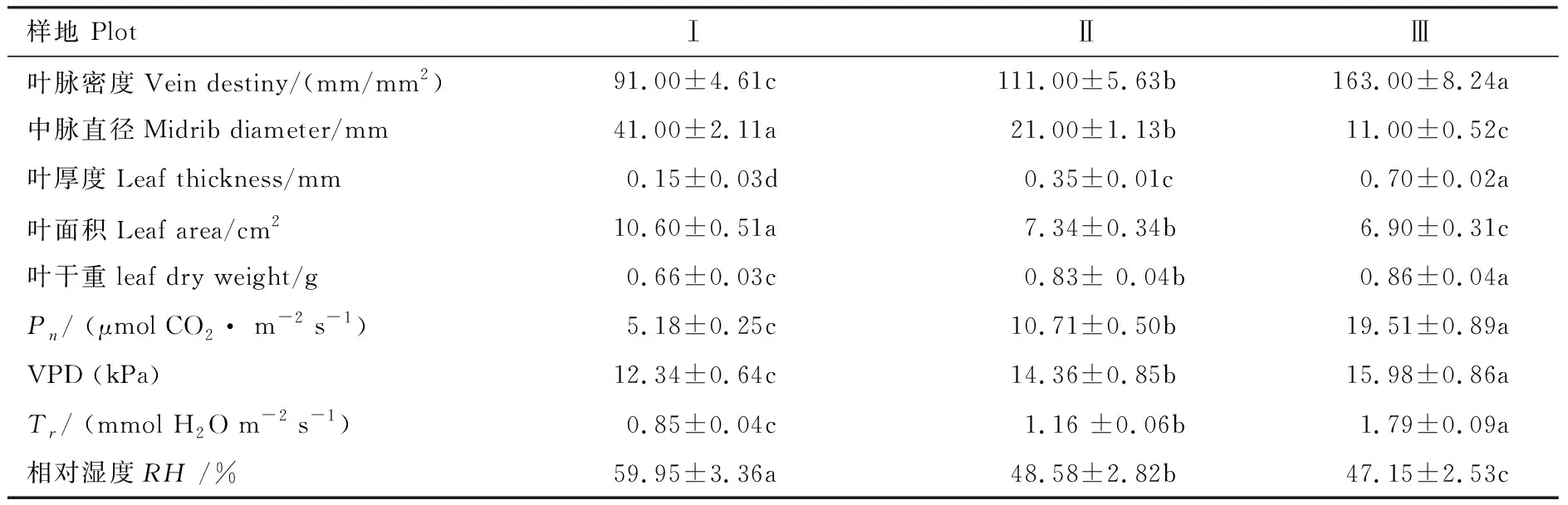
表2 不同密度下滨藜叶性状特征(平均值±标准误差)
Pn:净光合速率,net photosynthetic rate;VPD:饱和水汽压差,vapor pressure deficit;Tr:蒸腾速率,transpiration rate;RH:相对湿度,relative humidity;同行不同小写字母表示样地间差异显著(P<0.05),n=108
2.3 不同密度下滨藜蒸腾速率(Tr)和叶脉密度的关系
不同生境中滨藜蒸腾速率和叶脉密度的权衡关系如图1所示。不同密度梯度下滨藜蒸腾速率(Tr)与叶脉密度的关系存在显著差异(P<0.05)。通过SMA分析得到,滨藜蒸腾速率与叶脉密度在高密度(I)和低密度(III)均呈极显著正相关关系(P<0.01),在中密度(II)呈显著正相关关系(P<0.05)。滨藜在不同生境间的标准主轴化斜率存在差异,高密度(I)为0.97(95%的置信区间:CI=0.82, 0.99)、中密度(II)为1.00(95%的置信区间:CI=0.96, 1.05)、低密度(III)为1.25(95%的置信区间:CI=1.17, 1.26),从高密度(I)到低密度(III)呈现逐渐增大趋势,高密度(I)的斜率显著小于1.0(P<0.05),中密度(II)的斜率与1.0无显著差异(P>0.05),低密度(III)的斜率显著大于1.0(P<0.05)。从高密度(I)到低密度(III),滨藜叶脉密度与蒸腾速率呈显著的协同关系,蒸腾速率越大,叶脉密度越大,且在高密度(I)滨藜叶脉密度增加的速度小于蒸腾速率增加的速度;在中密度(II)叶脉密度与蒸腾速率呈等速的增长关系;在低密度(III)叶脉密度增加的速度大于蒸腾速率增加的速度。
2.4 不同密度下滨藜蒸腾速率(Tr)和中脉直径的关系
不同生境中滨藜蒸腾速率和中脉直径的权衡关系如图2所示。不同密度梯度下滨藜蒸腾速率(Tr)与中脉直径的关系存在显著差异(P<0.05)。通过SMA分析得到,滨藜蒸腾速率与中脉直径在高密度(I)和低密度(III)均呈显著负相关关系(P<0.05),在中密度(II)不存在相关关系(P>0.05)。滨藜在不同生境间的标准主轴化斜率存在差异,高密度(I)为-0.85(95%的置信区间:CI=-0.86, -0.81)、中密度(II)为-1.04(95%的置信区间:CI=-1.08, -0.99)、低密度(III)为-1.30(95%的置信区间:CI=-1.36, -1.27),从高密度(I)和低密度(III)呈现逐渐减小趋势,高密度(I)的斜率显著大于-1(P<0.05),中密度(II)的斜率与-1无显著差异(P>0.05),低密度(III)的斜率显著小于-1(P<0.05)。从高密度(I)到低密度(III),滨藜中脉直径与蒸腾速率呈显著的权衡关系,蒸腾速率越大,中脉直径越小,且在高密度(I)滨藜中脉直径减小的速度大于蒸腾速率增加的速度;在中密度(II)中脉直径与蒸腾速率呈等速变化关系;在低密度(III)中脉直径减小的速度小于蒸腾速率增加的速度。
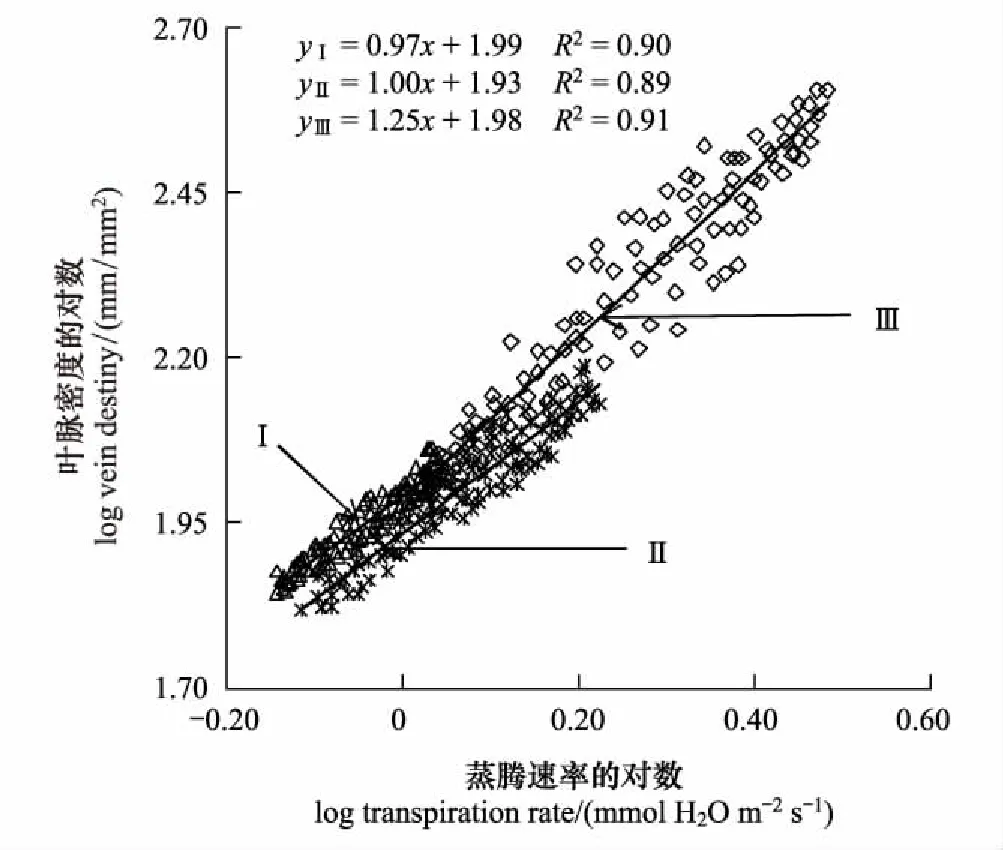
图1 不同密度下滨藜蒸腾速率(Tr)和叶脉密度的关系
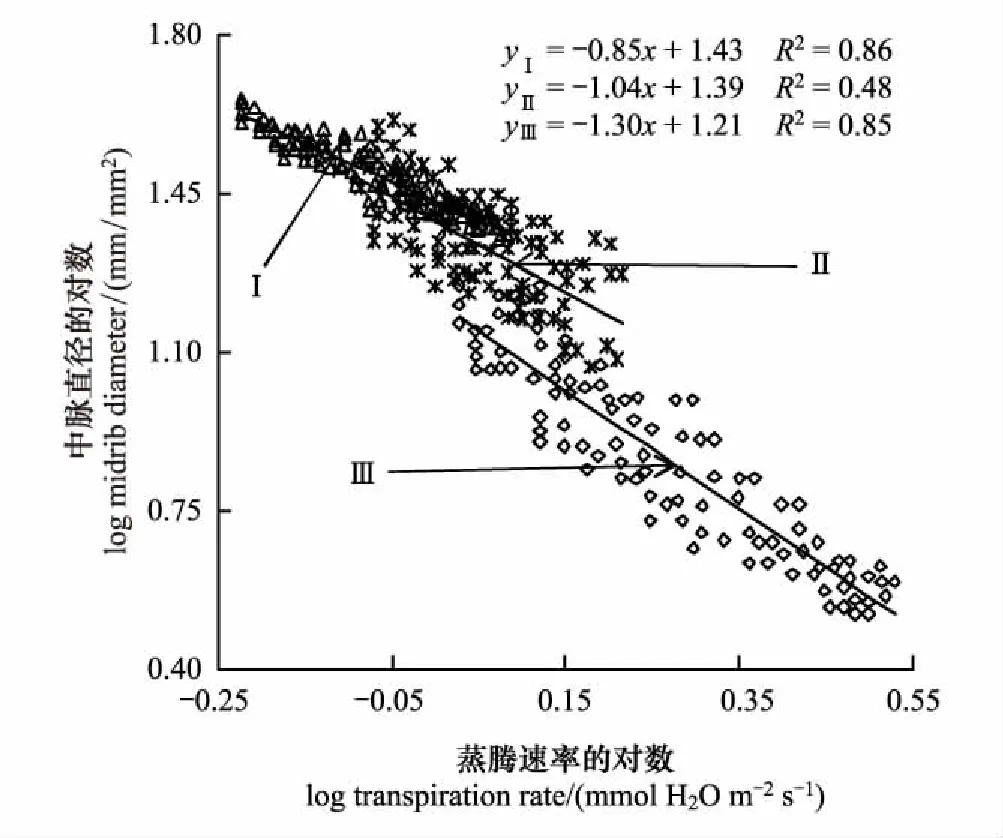
图2 不同密度下滨藜蒸腾速率(Tr)和中脉直径的关系
3 讨论
植物叶片的蒸腾速率能够影响或指示植物对环境的适应特征或能力[29],植物往往通过叶脉功能性状的调整,形成与蒸腾速率相适应的光能拦截、光合碳获取、水分供给和展叶支撑系统[30]。研究发现,在高密度(I)和低密度(III)样地,滨藜叶片蒸腾速率与叶脉密度呈极显著正相关关系(P<0.01),与中脉直径呈显著负相关关系(P<0.05);在中密度样地(II),滨藜叶片蒸腾速率与叶脉密度呈正相关关系(P<0.05),与中脉直径关系不显著(P>0.05)(图1, 图2)。不同密度湿地群落滨藜叶脉性状与蒸腾速率关系的空间异质性,既是植物应对邻体间遮阴、可获得性光合有效辐射和空间资源竞争的表型可塑性,也是密度制约下植物权衡碳投入-收益的必然结果。
在密度制约和邻体干扰条件下,植物需要在光合产物有限性约束下合理配置叶肉、叶脉等叶性状[31],以满足植物叶片光合和蒸腾的生理需求,进而达到生存和生长之间的协同适应[32],实现光合碳获取效率的最大化。低光照条件下植物叶脉构型的改变是以较低的光合生理需求以及较高的水分运输能力和利用效率为基础保障,最大限度的减少物种生态位重叠,提高植物在异质生境中的资源利用效率[12]。在高密度样地,湿地群落的盖度和高度最大(表1),滨藜生长繁茂、植株之间产生了严重的邻体干扰,光竞争成为影响植物生长的首要胁迫因子,滨藜叶片的蒸腾速率(Tr)较低,与叶脉密度呈极显著正相关关系(P<0.01),与中脉直径呈显著负相关关系(P<0.05)(图1, 图2)。主要原因有:(1)强烈的遮阴影响了到达群落内部的光资源的同时也降低了土壤水分的潜在蒸散,缓解土壤盐碱化,促进有机质积累[11],使得土壤水分含量较高(表1),缓解了叶片光合作用的水盐胁迫,同时滨藜获得的光和有效辐射水平较低(表1),温度变幅较小,使得叶片与空气间形成较小的水汽压差(VPD),叶片失水相对较少、蒸腾速率较低,滨藜不需要构建高的叶脉密度亦能满足叶片的供水需求,因此,生长在高密度样地的滨藜选择了较低的叶脉密度,使蒸腾速率与叶脉密度呈极显著正相关关系(P<0.01)。(2)中脉直径是表征叶脉系统机械支撑能力的主要指标[9],在邻体遮蔽严重和群落内部光合有效辐射较低的状态下,为了提高叶片的光拦截和光合碳获取能力,使有限的光合产物创造最大的光截获面积,滨藜叶片中的叶肉细胞趋于平铺状排列,即形成增大叶面积和减小叶干重的“大叶”构建模式,相应的需要增大中脉直径以满足展叶的功能需求,使中脉直径与蒸腾速率呈显著负相关关系(P<0.05)。
为了顺利完成自身的生命过程且实现光合产物投资-收益的最大化,植物会主动调整、转换和补偿叶脉性状的功能及状态[33],将生物量优先分配给具有最易获取能力的性状,是植物提高资源利用效率的生活史对策[22]。在低密度样地,滨藜叶片蒸腾速率增加,与叶脉密度呈极显著正相关关系(P<0.01),与中脉直径呈显著负相关关系(P<0.05)(图1, 图2),主要原因有:(1)该生境种间遮蔽较小,光环境明显改善,群落内部的光合有效辐射增大,在空气相对湿度(RH)降低(表2),伴随着叶片的饱和水汽压差增大,导致水分扩散阻力减小,使得蒸腾和光合作用旺盛(表2),同时土壤含水量降低(表1),叶脉系统必须增加水分运输的量与速率,才能有效地吸收土壤水分和养分以适应较高的光合需求[34],因此,滨藜选择了大的叶脉密度,从而增大了水分离开叶脉到叶肉细胞的交换面积,提高了水分运输通道的选择性,缩短了水分运输到蒸腾部位的距离,使得叶片在水分胁迫造成叶脉木质部栓化的情况下,能够通过栓塞周围的叶脉运输水份保证叶片的高蒸腾速率;(2)在干旱和光照充足条件下,大而薄的叶片相对更易蒸腾失水,叶片寿命短、耗损速率高,不利于光合碳的获取[35]。低密度湿地群落,土壤含水量低(表1),滨藜叶片长期暴露在强烈光照环境下,为了避免灼伤、减少叶片空穴和栓塞现象引起的水分运输阻力[36- 37]以及提高叶片的光合碳同化效率,滨藜选择了小而厚的叶片,小叶模式不仅有助于植物实现热量和物质交换的目的,同时叶肉组织更接近中脉,有利于控制叶片内自由水的含量,维持植物叶片内水分的动态平衡[11],有效避免了植物组织面临水分亏缺的风险,因此,滨藜不需投入过多的生物量用于构建叶片的支撑系统,从而选择了较小的中脉直径,以便更好地满足植株在高蒸腾速率下的水分供需平衡。
叶片是植物重要的物质生产器官,其生长规律直接决定了植物的光合作用效率、蒸腾速率及资源分配策略[38],叶片大小和叶脉密度的资源权衡机制,体现了密度制约、邻体干扰等因素影响下植物叶片功能性状的表型可塑性[32]。在中密度样地,植株叶片接收的光资源充足,滨藜进行了叶内生物量分配和叶片构型的调整,适度增加了叶面积,减小了叶厚度,形成了大而薄的叶片和较高的比叶面积,拉近了气孔与叶表面的距离,提高了叶片的气孔导度,将蒸腾速率和净光合速率维持在适中水平(表2)。中密度湿地群落土壤水分相对充足,滨藜植株不需要构建强大的供水系统亦可保证自身正常的生理活动,没有投入过多生物量用于叶片的支撑,中脉直径变化不大。因此,生长在该样地的滨藜在兼顾植株水分输送、利用与保持等功能的前提下,形成了均衡的叶片生长模式,使植株蒸腾速率与叶脉密度呈现出显著的相关性(P<0.05),与中脉直径间不存在相关关系(P>0.05)(图1, 图2)。
4 结论
植物蒸腾速率与叶脉性状的关系是植物叶片生理功能对环境适应过程的表现,体现了密度制约下叶脉密度和中脉直径之间的资源权衡机制。本研究发现:随着密度的改变,滨藜群落环境和土壤水分发生了明显变化,滨藜通过调整叶脉性状的特征来适应环境的变化。蒸腾速率与叶脉密度间存在一种协调性,生长在高密度样地的滨藜蒸腾速率较低,为了获得足够的光合产物,叶片选择了较小的叶脉密度和较大的中脉直径,以竞争更多的光资源;而生长在低密度样地的滨藜选择了增大叶脉密度,减小中脉直径的构建模式,实现了高蒸腾速率及水分利用效率,较好地验证了植物对生境异质性的适应机制。本文仅从密度制约视角分析了滨藜叶脉性状与蒸腾速率的关系,土壤养分、地形、植物激素等也是影响植物叶脉性状与功能的重要因素,探索不同因素的交互作用对湿地植物叶片光合作用适应性的调控机制,是今后关注的重点。
